
GHRHR 基因在湘西黄牛不同组织中表达差异性研究
2020-10-22 12:09资琼涛黄胥莱陈东万发春沈维军易康乐唐江权李付强
中国畜禽种业 2020年10期
资琼涛 黄胥莱 陈东* 万发春 沈维军 易康乐 唐江权 李付强
(1,湖南农业大学动物科学技术学院 410128;2,湖南畜牧兽医研究所 410130;3,湖南天华实业有限公司 417000)
随着人们收入水平的不断提高,人们的消费从追求数量转而追求质量,牛肉以其高营养价值也渐渐受到消费者的青睐。肉牛体内的肌内脂肪沉积,不仅影响牛肉的断面大理石评分,还可显著提高牛肉的嫩度、多汁性和风味等感官品质,从而最终影响其肉质等级与经济价值[1]。脂肪沉积量主要取决于脂肪细胞的增殖和分化程度,这一过程受许多分化转录因子的协同调控作用,转录因子的表达量及其活性的大小决定了分化过程[2]。因此,研究湘西黄牛脂肪沉积调控的分子机制对品种选育具有重要意义。
GHRHR 属于G 蛋白偶联受体家族的成员之一,对动物机体的生长发育起关键作用,主要由GHRHR 基因调控分泌[3]。GHRHR 主要通过与GHRH 结合,以cAMP 为第二信使的信号通路激活垂体细胞合成并分泌GH,从而调控动物机体的生长发育[4]。张存芳等[5]等用PCR-SSCP 和DNA 测序鉴定GHRHR-5'UTR 中的一个新的单核苷酸多态性(NM_181020:c.102C>T),发现该位点与纽约牛6 个月的体重(BW)显著相关。刘艳丽[6]等对西农萨能奶山羊和关中奶山羊的GHRHR基因进行研究时发现,P3 位点上的多态性与西农萨能奶山羊的身高和体长相关性显著,与关中奶山羊的胸围、身高和体长显著相关。这些研究都表明,GHRHR 基因对动物机体体重、身高等都有重要的影响。基于目前的研究现状,本研究旨在研究GHRHR 基因在湘西黄牛不同组织中的发育性表达,为改善牛肉品质及选育优良品种提供可借鉴的遗传学资料。
1 材料与方法
1.1 试验动物与饲养管理
分别选取湖南德农牧业集团有限公司国家级湘西黄牛保种场6、18 月龄和30 月龄湘西黄牛各10 头。试验牛在4 月龄时断奶,每日精补料按照体重0.5%饲喂,精补料由55%玉米、豆粕24%、小麦麸15%、预混料4%、食盐0.5%、小苏打0.5%和碳酸氢钙1%,其中每千克预混料含VA162.5K IU,VD25K IU,VE500 IU,碘12.5mg,钴2.5mg,铜250mg,铁125mg,锌750mg,锰1g,硒7.5mg。于每天的07:00 和14:30 进行饲喂,先粗后精,粗料由稻草组成,粗料自由采食,自由饮水。所有试验牛均健康无病,免疫程序一致,并进行统一管理。
1.2 屠宰及样品采集
在6、18 和30 月龄试验牛中随机挑选4 头体况和体重相近的牛,空腹后于晨饲前进行屠宰取样,分别采集肝脏、12~13 肋骨背最长肌、皮下脂肪和腹腔脂肪4 个组织的样品,迅速放入液氮中保存,之后置于实验室-80℃冰箱保存备用。
1.3 主要试剂
TRIzol 试剂盒(Thermo Fisher Scientific 公司)、逆转录试剂盒(北京百泰克生物技术有限公司)、荧光定量试剂盒(百泰克);氯仿和无水乙醇均购自康为世纪生物有限公司。
1.4 引物设计及合成
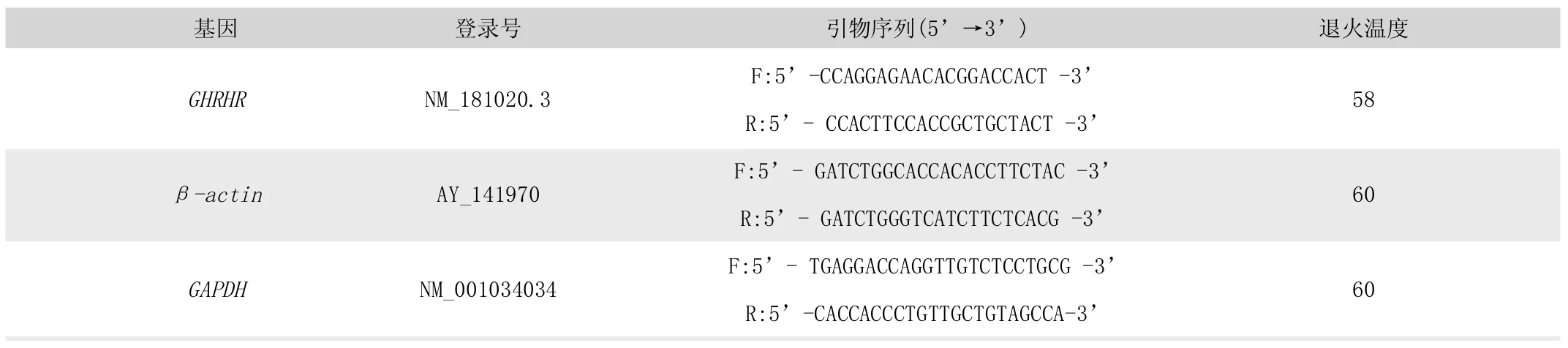
表1 实时定量PCR 引物序列
根据GenBank 中牛GHRHR 基因序列,采用Primer Premier 5.00 软件设计特异引物,以牛GAPDH 和β-actin 基因作为内参,由上海生工生物股份有限公司设计合成实时定量PCR 引物,引物序列见表1。
1.5 总RNA 提取及cDNA 第一条链合成
取约50mg 的组织样,置于液氮中研磨之后,用Trizol 法(Thermo Fisher Scientific 公司) 提取总RNA。用超微量紫外分光光度计检测其浓度和OD 值(OD260nm/OD280nm=1.8~2.0)。用超微量分光光度计定量,OD260nm/OD280nm值均在1.82~2.04 之间,说明提取的总RNA 纯度较高,可以用于后续的反转录及实时荧光定量PCR。根据逆转录试剂盒(北京百泰克生物技术有限公司)说明书将总RNA(1ug)反转成cDNA。20uL 体系,反应条件为:65℃ 5min,42℃60min,25℃5min,70℃5min,逆转录后的cDNA 置于-80℃冰箱保存。
1.6 实时荧光定量PCR
采用SYBR Green 荧光染料法在实时荧光定量PCR 仪上进行定量。反应体系(20ul):2×Fast SYBR.Green Master Mix 10μL,cDNA 1μL,无菌水8μL,下游引物0.5μL 和上游引物0.5μL。反应条件:95℃15s,95℃5s,60℃30s,95℃60s,60℃60s,40 个循环后进行溶解曲线分析,以每5s 上升0.5℃的速率从65℃升高到95℃,荧光信号在循环结束时检测,每个样品做3 个重复。PCR 反应产物用1.0%琼脂糖凝胶电泳进行检测。
1.7 统计分析
实时荧光定量PCR 得出的基因原始表达量用2-△△Ct法计算,2-△△Ct表示试验目的基因mRNA 的相对表达量。利用SAS 9.0 软件one-way ANOVA 程序进行统计分析,表示差异显著,P<0.01 表示差异极显著。
2 结果
2.1 GHRHR 基因在不同月龄湘西黄牛的相同组织中发育性表达的差异
由表2 可知,湘西黄牛在18 月龄GHRHR 基因的表达量极显著高于6 月龄和30 月龄GHRHR 基因的表达量30 月龄GHRHR 基因的表达量极显著高于6 月龄
2.2 GHRHR 基因在相同月龄湘西黄牛不同组织中的表达差异
由表3 可知,GHRHR 基因在6 月龄的湘西黄牛各组织中的表达量差异不显著在18 月龄时,背最长肌与皮下脂肪中GHRHR 基因的表达量显著高于肝脏中的表达量30 月龄时,各组织中的表达量两两之间差异显著且皮下脂肪>背最长肌>腹腔脂肪>肝脏。

表2 GHRHR 基因在不同月龄湘西黄牛中的表达差异

表3 GHRHR 基因在湘西黄牛不同组织中的发育性表达
3 讨论
Wang 等[7]研究发现,GHRHR 基因的表达主要局限于垂体或大脑。王英明等[8]以泸宁鸡和罗斯白羽肉鸡为研究对象,发现GHRHR 基因主要在垂体中表达,在其他组织上极少表达或基本不表达。本研究发现,GHRHR 基因在湘西黄牛脂肪代谢关键部位均有表达,随着月龄的增加,GHRHR 基因的表达量先增加后减少,在18 月龄的表达量达到最高,说明GHRHR基因的表达与湘西黄牛生长发育阶段有关,且GHRHR 基因在各组织的表达存在月龄间差异性。在6 月龄时,各组织间GHRHR 基因的表达量无显著差异;而18 月龄和30 月龄时肝脏中GHRHR 基因的表达量最低,皮下脂肪最高,且各组织间GHRHR 基因的表达量存在显著差异,这说明GHRHR 基因的表达量存在组织差异性。
研究表明,GHRHR 基因在生殖系统中受GH 基因的调控发挥生物学功能[9]。脂肪沉积与脂肪细胞的发生、脂肪合成和脂解作用有关,GH 可促进脂类分解,抑制脂肪的合成[10,11]。Straus 等[12]研究报道GH 可诱导前脂肪细胞系(3T3-L1、3T3-F442A 和Ob1771)中的前脂肪细胞分化为脂肪细胞,促进脂肪生成。而Wabitsch 等[13]在体外原代培养脂肪的前体细胞试验中发现,GH 显著降低新脂肪细胞的生成,且剂量依赖性的降低甘油3-磷酸脱氢酶的活性,抑制脂肪的生成。上述研究表明,GH 对脂肪生成的影响与其对脂肪的合成和分解作用存在一定差异性。Alba 等[14]研究发现,GHRHR 敲除小鼠的体重、肝脏重、肾脏重和心脏重均低于正常小鼠。张海波等[15]通过敲除小鼠皮下脂肪组织的CD29 和CD34 基因相对表达量的测定发现,GHRHR 基因可以增加干细胞的增殖,促进脂肪沉积。上述研究表明,GHRHR 基因在动物机体生长发育及脂肪代谢过程中发挥重要作用。本研究初步探讨了6 月龄、18 月龄和30 月龄湘西黄牛脂肪代谢关键部位GHRHR 基因的表达规律,对GHRHR 基因与牛脂肪沉积之间的关系有待进一步探究。
4 结论
GHRHR 基因在6 月龄、18 月龄及30 月龄湘西黄牛脂肪代谢关键部位均有表达,且差异显著,存在月龄和组织间的差异性。GHRHR 基因对湘西黄牛机体脂质代谢具有重要作用,为湘西黄牛品种选育及肉质改善奠定了基础。
猜你喜欢
中国食物与营养(2022年5期)2022-06-17
当代畜禽养殖业(2020年9期)2020-10-21
家庭医学·下半月(2020年2期)2020-04-26
科学生活(2019年7期)2020-01-01
创新作文(3-4年级)(2019年3期)2019-09-03
湘潮(上半月)(2019年4期)2019-05-22
散文诗(2017年18期)2018-01-31
文艺论坛(2017年4期)2017-05-08
创业家(2015年2期)2015-02-27
文艺生活·上旬刊(2009年4期)2009-06-11
