
Species richness,forest types and regeneration of Schima in the subtropical forest ecosystem of Yunnan,southwestern China
2020-10-20 08:13CindyTangPengBinHanShuaifengLiLiQinShenDiaoShunHuangYunFangLiMingChunPengChongYunWangXiaoShuangLiWeiLiWeiWangandZhiYingZhang
Forest Ecosystems 2020年3期
Cindy Q.Tang ,Peng-Bin Han,Shuaifeng Li,Li-Qin Shen,Diao-Shun Huang,Yun-Fang Li,Ming-Chun Peng,Chong-Yun Wang,Xiao-Shuang Li,Wei Li,Wei Wang and Zhi-Ying Zhang
Abstract
Keywords: Forest structure, Regeneration dynamics,Schima,Species richness,Forest succession, Yunnan
Background
Schima, a genus belonging to Theaceae,is widely distributed in the eastern Himalaya of Nepal,Bhutan,northeastern India,Bangladesh and northern Myanmar, Indochina (Thailand,Cambodia,Laos,and Vietnam),southern China,Japan(Ryukyu Archipelago and Ogasawara Islands),Malaysia,Indonesia and Brunei (Fig. 1a, Table 1). Fossil species of Schima have been found from the Miocene of Germany(Mai 1971),Yunnan of China(Li et al.2013a),Japan(Choi et al.2010),Styria of Austria(Kovar-Ender and Hably 2006)and the Oligocene of Guangxi of China (Shi et al. 2017). Recent molecular phylogenetic analysis showed Schima is closely related to Gordonia and Franklinia,forming the monophyletic tribe of Gordoniae (Prince and Parks 2001; Li et al. 2013b; Zhang et al. 2014). There has been an argument as to the number of species of Schima (Prince 2007). For example, Bloembergen (1952) recognized only one species, Schima wallichii with nine subspecies and three varieties. Airy Shaw (1985)mentioned that Schima has 15 species. Min and Bartholomew (2007) and Byng (2014) reported that there are about 20 species. We adapt Min and Bartholomew (2007) that 13 species present in China;among them 6 species are endemic to China.Most of the 13 species occur as one of dominants with some species of other genera in the subtropical evergreen broad-leaved forest ecosystem of China.The evergreen broad-leaved forests of China are generally found in large,discontinuous patches in the subtropical zone of China.However, due to human intensive activities on the subtropical ecosystem many primeval evergreen broad-leaved forests were replaced by secondary vegetation in China (Wu 1980;Tang 2015a,b).
Among the Schima species of China, Schima superba which widely distributed in the eastern subtropical regions of China, has been studied from aspects of its forest communities (e.g. Wang and Li 1999; Yang et al. 2006; Song 2007), population structure (e.g. Cai and Song 1997; Hu 2005; Li et al. 2008) and genetics (e.g. Zhang et al. 2006;Jin et al. 2007). In contrast, to date the knowledge on the other species of Schima is very scarce.It is critical to elucidate spatial patterns of species richness of the genus and ecological role of other Schima species in forest succession and regeneration dynamics of the subtropical ecosystem.
There has been a longtime debate as to whether forests dominated by Schima species are early or late successional forests. Eight species of Schima are distributed in Yunnan, southwestern China. Yunnan affords us a unique opportunity to explore forests containing Schima species as one of the dominants in various landscape contexts. Understanding species richness patterns,forest types and regeneration characteristics of Schima are a prerequisite to understand the dynamics of the forests and provide a reference for forest reforestation and site restoration programs. We address the following questions: (1) How are the distribution patterns of species richness of Schima in Yunnan as compared with those in other regions of subtropical China? (2) What are the forest types and structure dominated by Schima species? (3) What are the regeneration dynamics of Schima species in Yunnan?
Methods
Study area
Yunnan is a highland province (21°08′-29°15′ N,97°31′-106°11′ E) located in southwestern China, and having the greatest elevation difference on earth, from the northwest (6740 m a.s.l.) to the southeast (76 m a.s.l.).Set in the foothills of the Himalayas and Tibet, Yunnan’s climate ranges from the icy highland climate of the northwestern frontier to the tropical lowland climate of the area bordering Myanmar and extending south and east toward Laos and Vietnam (Fig. 1b). The vegetation is from lowland tropical, to hilly/mid-elevational subtropical evergreen broad-leaved forests to alpine coldtemperate coniferous forests, scrubs, meadows and grasslands at the high altitudes. The Yunnan area is included in ‘Indo-Burma’ global hotspot of biodiversity(Myers et al. 2000). Most of the province falls into latitudes classified as middle subtropics (25°00′-29°15′ N)and southern subtropics (22°30′-25°00′ N). The small area between 21°08′-22°30′ N is considered as northernmost tropics. The climate is monsoonal, with rainy summers. The monsoon, moving mainly from the Bay of Bengal, causes heavy rainfall on the southern aspect of the plateau, then it diminishes northward, creating a rainfall gradient. About 80% of the rain falls during rainy season-summer, and the most of the remainder falls in autumn and spring, leaving the winter cool and dry. The subtropical forests of Yunnan are remarkable for their globally high plant diversity and extraordinary local endemism and diverse plant communities (Tang 2015a).
Study species
We made efforts to study species of Schima in the wild in Yunnan Province. Among the thirteen Schima species of China, eight occur in Yunnan (Table 1). Since one(Schima brevipedicellata) of the eight species does not appear as a dominant in the forests, we exclude it. Thus,our focal species in this paper are seven species, i.e.
Schima khasiana, Schima noronhae, Schima villosa,Schima sinensis, Schima wallichii, Schima sericans,Schima argentea, each of which is found as one of the dominants in the primary or secondary subtropical evergreen broad-leaved forests of Yunnan. Most of the seven species become quite tall (some can reach 45 m).They have showy, fragrant white flowers, with five petals and numerous yellow stamens. The leaves are alternate,spirally arranged, and entire or crenate or serrate.Schima flowers are hermaphrodite, axillary and solitary,or rarely three to five flowers are crowded along a short raceme. The fruit is a woody, globose or subglobose capsule, which splits for half its length into (four to) five (to six) segments; a stout, central column remains. The seeds are small and kidney-shaped, flat, with a narrow marginal wing. Representative characteristics of the Schima species and their forest communities are shown in Figs. 2a-l.

Table 1 Schima species of East Asia.Data Sources: this study,eFlora of China (http://flora.huh.harvard.edu/china/),Chinese Virtual Herbarium database(http://www.cvh.org),GBIF database(www.gbif.org) and International Union for Conservation of Nature(www.iucnredlist.org)
Data collection and analysis
The forests in the study area are subjected to a range of natural and anthropogenic factors such as elevation, topography, natural disturbances and human activities; thus,forests are structurally and floristically heterogeneous and the landscape pattern of vegetation takes the form of mosaic patches. We selected plots in each patch containing Schima species in the study area. During 2015-2019, we established 71 plots dominated by Schima species in Yunnan (Fig. 3). The plots were established in the locations where it was possible for us to access. The plot size varied between 20 m×30 m to 30 m×40 m where plot size depended on the size of a patch. Patch size was determined by species composition and topographic similarity.
For the species in each plot,all individuals at least 1.3 m in height were identified to species level, numbered and tagged, and their diameter at breast height (DBH) and height(H)were recorded.In addition,general information about each plot was noted, such as slope positions, altitude, slope exposure, slope inclination, as well as disturbance histories. Tree stems were classified into four types based on their vertical position, crown position, and height: emergent layer (greater than 28 m tall), canopy(20-28 m tall), subcanopy (8-20 m tall), and shrub layer(1.3-8 m tall).All understory woody species less than 130 cm tall,were divided into two classes:(1) 5-50 cm tall for seedlings and (2) 50-130 cm for saplings. Within these two classes, each individual was identified, counted, and measured for height and percent cover.
Based on tree sizes, ages of the forest stands, species composition and community structure, as well as degrees of naturalness or disturbance regime of the forests,we judged whether or not the forest stands were in early,middle, or late successional stages.
For all individuals greater than 1.3 m tall,DBH was used to calculate basal area and then basal area(BA)for each species found in a plot could be determined.In each plot,the relative importance value [RIV=(Relative density+Relative basal area)/2)]of each species was used to measure the abundance of the species.Plant communities were classified using a floristic similarity dendrogram with Relative S orensen and Group Average clustering [PCORD software (McCune and Mefford 1999)].The communities were named according to dominant species. Diversity was calculated for each forest stand using species richness (number of species), the Shannon-Wiener’s diversity index (Shannon-Wiener index,H′; Pielou 1969) and Simpson’s diversity index (Simpson index,D;Lande 1996).Differences in species richness and diversity indices among habitats were analyzed by the nonparametric Kruskal-Wallis all-pairwise comparisons test,using Analyze-it software(United Kingdom).
Species richness (number of species) and the rarityweighted richness (RWR) (each species is assigned a score—based on the inverse of the number of sites in which a species occurs) (Williams et al. 1996) are used to show the Schima species distribution patterns. A square of 1° latitude ×1° longitude (a cell per a grid size)was chosen as a geographic unit to show species richness and rarity-weighted richness at present on the map,using ArcGIS v. 10.5. The correlation of species richness and rarity-weighted richness was analyzed using Pearson’s correlation coefficient.
Results
Schima species richness and rarity-weighted richness patterns
The patterns obtained for species richness and rarityweighted richness (the latter representing concentrations of limited-range species, as well as a high turnover of species between adjacent cells) show that the Schima species are confined to the subtropical and warmtemperate areas of China (Fig. 4a and b). Among 13 species found in China, 8 are distributed in Yunnan (Table 1). The areas of southeastern, south-central, southwestern and western Yunnan show the high species richness(4-7, yellow and red cells in Fig. 4a), while the other regions of China most often have 1-3 species (dark green,blue and light green cells) except for south-central China. The rarity-weighted richness (RWR) greater than 0.2 (yellow and red cells) was found in the areas of northwestern, southeastern, southern and south-central Yunnan, with two sites in Guangxi Province. The outstanding rarity weighted richness in northern Gaoligongshan, NW Yunnan is mostly contributed by Schima sericans which has a very narrow distribution (endemic to NW Yunnan and SE Tibet), and Schima khasiana which is limited to western and southern Yunnan and SE Tibet (Fig. 4b). The distribution patterns of Schima species richness (Fig. 4a) and rarity-weighted richness(Fig. 4b) in the Yunnan area have linear relationships with significant positive correlations (Pearson’s r=0.74).
Forest types, stratification and species diversity
Primary forests
According to the similarity dendrogram, vegetation data of 22 plots in Yunnan were classified into five primary forest types (Fig. 5a): Type PF1, the Schima noronhae-Castanopsis ceratacantha evergreen broad-leaved forest on mountain slopes at elevations 1920-2480 m in Wuliangshan, south-central Yunnan; Type PF2, the Castanopsis echidnocarpa-Schima khasiana evergreen broad-leaved forest at 2000-2710 m on mountain slopes in Wanmingshan, SW Yunnan; Type PF3, the Tetracentron sinense-Schima villosa deciduous and evergreen broad-leaved mixed forest on steep slopes and stream sides at 2450-2600 m in Zheonghoushan of the Ailao Mountains, south-central Yunnan; Type PF4, the Castanopsis platyacantha-Schima sinensis evergreen broadleaved forest on mountain slopes at 1800-1950 m in Daguan, NE Yunnan; Type PF5, Schima wallichii-Castanopsis hystrix evergreen broad-leaved forest in valleys or on mountain slopes at 950-1380 m in Xishuangbanna, S Yunnan. No major anthropogenic disturbances have been found, though some natural disturbances including slight landslides, moderate wind-blow and occasional lightning strike occurred in the forests.
In each forest type, Schima and Castanopsis (S. noronhae and C. ceratacantha in Type PF1, S. khasiana and C. echidnocarpa in Type PF2, S. sinensis and C. platyacantha in Type PF4, S. wallichii and C. hystrix in Type PF5) dominated the canopy (18-28 m), except that Type PF3 was dominated by Tetracentron sinense and S. villosa (Fig. 5b). In Type PF1, PF2, PF3, PF5 forests,Schima also reached emergent layer (28-36 m). In the subcanopy (8-18 m), the shrub layer (3-8 m), and the understory (<3 m) of the forest Types PF1, PF2, PF4,PF5, Schima was commonly found, while in Type PF3 it was absent in 8-12 m and infrequent in the shrub layer and the understory.
All the 22 plots of primary forests, taken together,contained 240 woody species, representing 147 genera in 72 families. Each forest type’s species composition with the relative importance value (RIV) is shown in Table 2.Type PF1 was dominated by Schima noronhae and Castanopsis ceratacantha. The other major components were Lithocarpus truncatus, Machilus longipedicellata,Eurya yunnanensis, etc. In the understory, a bamboo species-Yushania polytricha’s coverage reached 80%.Type PF2 was co-dominated by Castanopsis echidnocarpa and Schima khasiana along with Lithocarpus xylocarpus, Cyclobalanopsis myrsinifolia, Ternstroemia gymnanthera and Craibiodendron stellatum, etc. Type PF3 was co-dominated by Tetracentron sinense and Schima villosa, and the accompanying species were Illicium simonsii, Ilex triflora, Symplocos sumuntia, etc.Type PF4 was co-dominated by Castanopsis platycantha and Schima sinensis with Acer oliverianum, Lindera thomsonii, Pittosporum tobira, etc. Type PF5 was codominated by Schima wallichii and Castanopsis hystrix.The other components were Castanopsis mekongensis,Lithocarpus truncates, Wendlandia tinctoria and so on.The Shannon-Wiener diversity index H′ and the Simpson diversity index D ranged respectively from 2.364 to 3.076, and from 0.864 to 0.931 among the five forest types. The diversity indices were not significantly different among the various forest types (Table 2).
For per hectare basal area, forest Types PF2 and PF1 scored relatively high(111.35,109.33 m2·ha-1,respectively),followed by Types PF3 (75.25 m2·ha-1) and PF4 (61.55 m2·ha-1), and Type PF5 (40.37 m2·ha-1) (Supplementary Table S1).
Secondary forests
According to the similarity dendrogram based on vegetation data of 49 plots of secondary forests, there are 6 forest types(Fig.6a).Type SF1,the Schima argentea secondary evergreen broad-leaved forest was widely found on mountain slopes, in valleys, along roadsides or near farmland at elevations 1920-2650 m in Yongde daxueshan of SW Yunnan, Dachaoshan of SW Yunnan,Wuliangshan of south-central Yunnan and Jiaozixueshan of central Yunnan; Type SF2, the Schima khasiana-Dipentodon sinicus secondary evergreen and deciduous broad-leaved mixed forest was found along roadsides or stream sides at 2390-2650 m in Chaojian, NW Yunnan;Type SF3, the Schima wallichii secondary evergreen broad-leaved forest was commonly found on mountain slopes, in valleys, along roadsides or near farmland at 950-2460 m in SW, NW and S Yunnan; Type SF4, the Schima sericans-Castanopsis ceratacantha secondary evergreen broad-leaved forest was found on mountain slopes at 2040-2300 m in Yun County, SW Yunnan;Type SF5, Schima villosa-Rhoiptelea chiliantha secondary evergreen and deciduous broad-leaved mixed forest was found on mountain slopes at 1680-2100 m in Malipo County, SE Yunnan; Type SF6, the Schima sinensis-Castanopsis platyacantha secondary evergreen broad-leaved forest was found on mountain slopes at 1860-2110 m in Daguan County, Yiliang County and Zhenxiong County of Zhaotong, NE Yunnan. In thesecondary forests, there were evidences of anthropogenic disturbances such as occasional cattle and goats browsing or selective firewood cutting or collecting litters on the forest floor. Besides the moderate human activities,natural disturbances including landslides, wind-blow,and lightning strikes also occurred there. Especially in forest Types SF1 and SF3, there were many different types of disturbances.
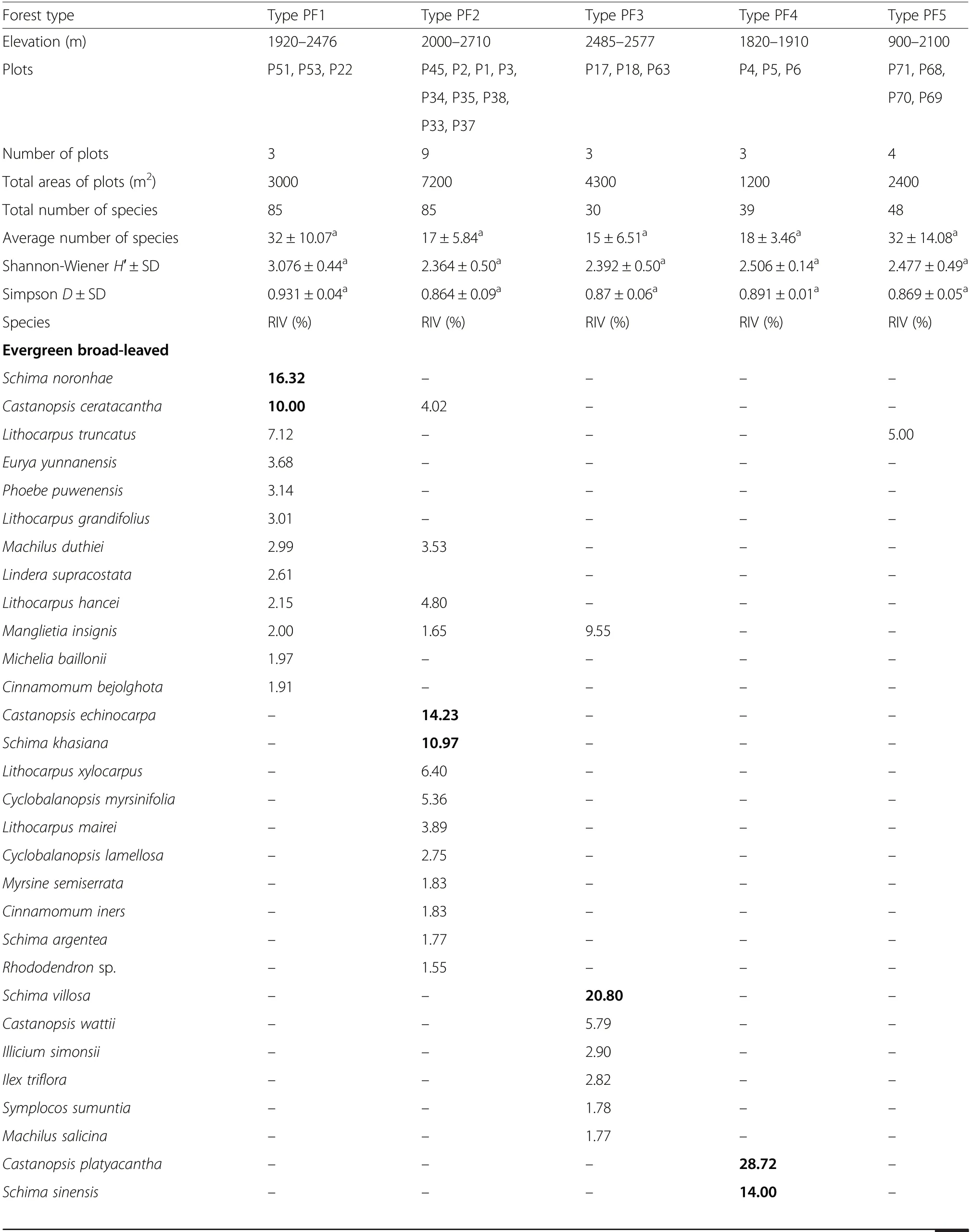
Table 2 Floristic composition of woody species (H ≥1.3 m) for primary forest types containing Schima species as one of the dominants.Only species with relative importance value(RIV)≥1.5% are shown.The same letter indicates no significant difference(p >0.05)for average number of species and diversity indices among those forest types.SD=Standard deviation.RIV=relative importance value.Dominant species are indicated by boldface
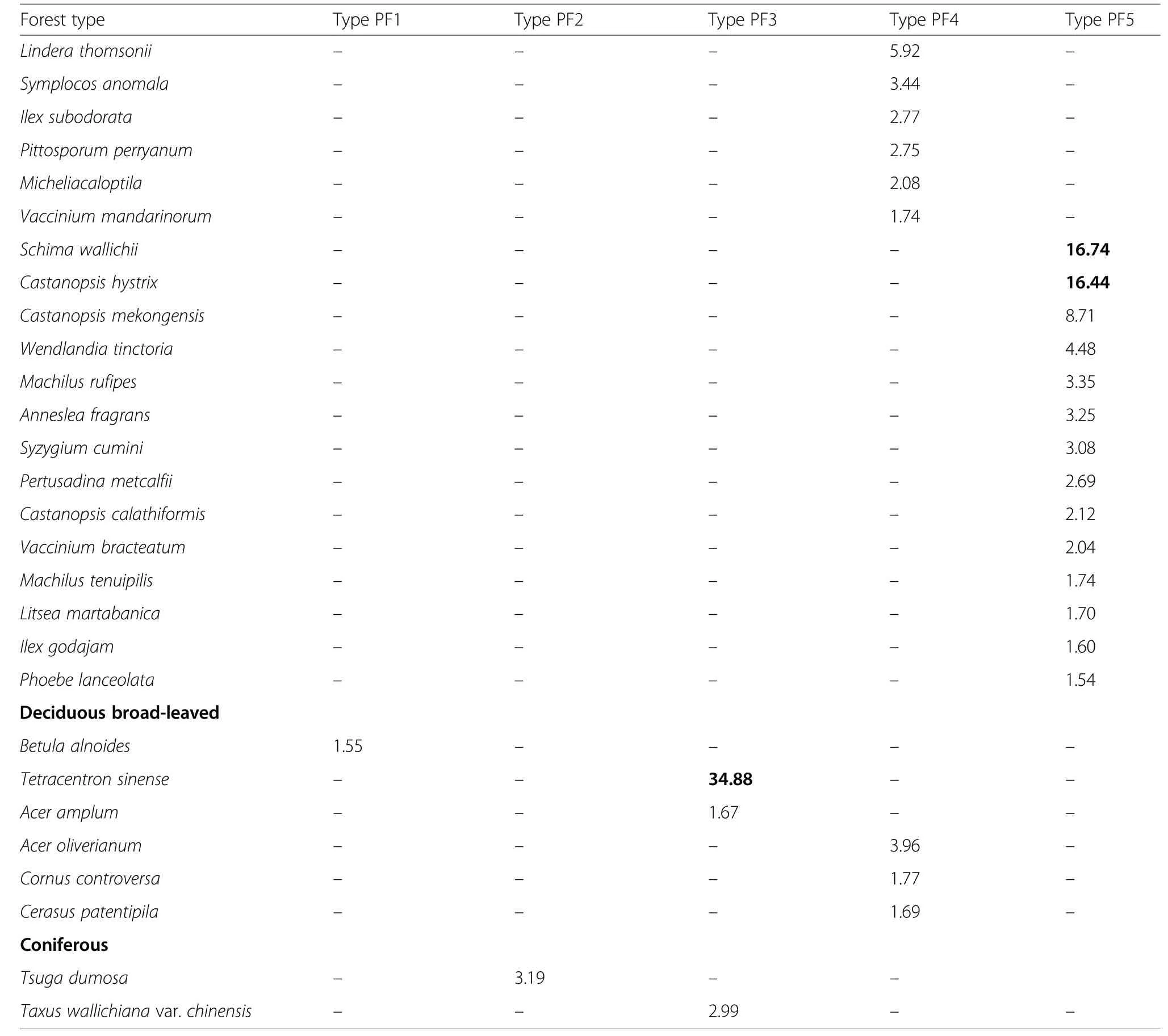
Table 2 Floristic composition of woody species (H ≥1.3 m) for primary forest types containing Schima species as one of the dominants.Only species with relative importance value(RIV)≥1.5% are shown.The same letter indicates no significant difference(p >0.05)for average number of species and diversity indices among those forest types.SD=Standard deviation.RIV=relative importance value.Dominant species are indicated by boldface(Continued)
In each secondary forest type, Schima species generally dominated the canopy (18-28 m) (Fig. 6b). In Type SF1, SF3, SF5 forests, S. argentea, S. wallichii and S. villosa reached the emergent layer (28-36 m).In the subcanopy (8-18 m), the shrub layer (3-8 m),and the understory (<3 m) of Types SF1, SF2, SF3,SF5, Schima was commonly found, while only few trees of S. sericans were in 8-28 m in Type SF4, S.sinensis was absent in 1.3-4 m in Type SF6. Except Type SF2, all the secondary forests had species of Castanopsis as a major component.
All the 49 plots of secondary forests, taken together,contained 370 woody species, representing 169 genera in 68 families. Species composition of each forest type with the relative importance value (RIV) is shown in Table 3.Type SF1, the Schima argentea secondary forest was dominated only by S. argentea. The other components were Castanopsis orthacantha, Castanopsis echinocarpa and Rhododendron delavayi, etc. Type SF2, the Schima khasiana-Dipentodon sinicus secondary forest was codominated by S. khasiana and Dipentodon sinicus. The other components were Illicium simonsii, Cyclobalanopsis augustinii and so on. In the understory,many Hypericum monogynum invaded. Type SF3, the Schima wallichii secondary was dominated only by S. wallichii,and its components were Castanopsis calathiformis,Lithocarpus xylocarpus, Betula alnoides, Alnus nepalensis, Pinus yunnanensis, etc. Type SF4, the Schima sericans-Castanopsis ceratacantha secondary forest, was codominated by S. sericans and C. ceratacantha, along with other components including Schima wallichii, Lyonia doyonensis, Ilex polyneura, Eurya nitida, Anneslea fragrans, Rhododendron vialii, etc. Type SF5, Schima villosa-Rhoiptelea chiliantha secondary forest was codominated by S. villosa and deciduous R. chiliantha.The companion species were Castanopsis ceratacantha,Lithocarpus truncates, Eurya impressinervis, Craibiodendron stellatum, ect. Type SF6, the Schima sinensis-Castanopsis platyacantha secondary forest was codominated by S. sinensis and C. platyacantha along with Lithocarpus hancei, Camellia pitardii, Symplocos anomala and so on. In the secondary forests, there were more pioneer deciduous broad-leaved species such as Alnus nepalensis, Betula alnoides and coniferous Pinus yunnanensis compared to the primary forests.
Among the 6 secondary forest Types SF1-SF6, the Shannon-Wiener diversity H′ and the Simpson diversity D respectively ranged from 2.109 to 2.429 and 0.793-0.950. No significant difference among the diversity indices were found in all the secondary forest types. However, the Shannon-Wiener indices of the secondary forests were significantly lower than those of the primary forests (p ≤0.05) (Table 3).

Table 3 Floristic composition of woody species (Height ≥1.3 m) for secondary forest types containing Schima species as one of the dominants.Only species with relative importance values (RIV)≥1.5% are shown.The same letter indicates no significant difference(p >0.05)for average number of species and diversity indices among those forest types.SD=Standard deviation.RIV=relative importance value.Dominant species are indicated by boldface

Table 3 Floristic composition of woody species (Height ≥1.3 m) for secondary forest types containing Schima species as one of the dominants.Only species with relative importance values (RIV)≥1.5% are shown.The same letter indicates no significant difference(p >0.05)for average number of species and diversity indices among those forest types.SD=Standard deviation.RIV=relative importance value.Dominant species are indicated by boldface(Continued)
For per hectare basal area, all the six secondary forest Types SF1-SF6 scored relatively low (23.23-45.95 m2·ha-1, Supplementary Table S2). It is clear that the basal area per hectare of the secondary forests is much lower than that of the primary forests.
Along latitudinal, longitudinal and elevational gradients, the distribution pattern of forests containing Schima species as one of dominants in Yunnan is shown in Fig. 7. For both the primary forests and the secondary forests as a whole, Schima species were found at elevations between 990 and 2600 m. S. noronhae as a top dominant was only found in primary forests in Wuliangshan, while S. wallichii appeared in both primary and secondary forests in Xishuangbanna, Nangunhe and Yongde daxueshan. S. khasiana in Gaoligongshan,Schima villosa in Ailaoshan and Malipo, S. sinensis in Daguan as a second or third dominant appeared in primary forests. However, the four species, i.e. S. wallichii,S. khasiana, Schima villosa, and S. sinensis, were more often seen as a top dominant found in secondary forests,and S. argentea and S. sericans were only found as a top dominant in secondary forests.
Regeneration dynamics
The DBH-class frequency distribution of species can reveal their population structure and regeneration dynamics. In the primary forests, the DBH-class frequency distribution of each of the five Schima species i.e. S. noronhae, S. khasiana, S. wallichii, Schima villosa, S. sinensis shows a sporadic type with several peaks, indicating that recruitments varied by chance with the regeneration mainly depending on disturbances (e.g. canopy gap regeneration) (Fig. 8). S. noronhae had peaks at 25-30 and 45-50 cm DBH, with very few individuals less than 25 cm DBH. Some S. noronhae trees larger than 80 cm DBH were found, and its maximum DBH reached 145 cm (maybe ca. 280 years old based on our limited data of core samples). S. khasiana had a discontinued distribution between 5 and 155 cm DBH, and more individuals smaller than 10 cm DBH were found. S. wallichii had relatively more trees in 30-35 and 5-10 cm DBH,and reached 135 cm DBH (maybe ca. 152 years old based on our limited data of core samples) but was absent in 40-45,55-60,80-100,105-130 cm DBH.Schima villosa only had 2 trees in the DBH-classes less than 50 cm,then it discontinuously distributed in 55-105 cm DBH.Schima sinensis had more trees in 10-25 cm DBH and reached 70 cm DBH, but was completely absent between 35 and 69 cm DBH. For all the Schima species except S.khasiana,it evidently shows the great lack of young trees and poor regeneration. The DBH-class frequency distribution of the major species of each primary forest type is provided in Supplementary Table S1.
In the secondary forests, the DBH-class frequency distribution of each of the six Schima species, i.e. S. argentea, S. khasiana, S. wallichii, S. villosa, S. sinensis and S.sericans also generally shows a sporadic type with multipeaks, indicating that recruitments varied by chance with the regeneration depending on disturbances (e.g. canopy gap regeneration) (Fig. 8). Since moderate human activities or moderate natural disturbances in the secondary forests (Fig. 6a), many forest gaps formed, so abundant individuals of S. argentea, S. khasiana, S. wallichii in DBH-classes smaller than 30 cm, and overwhelmingly more young trees in 0-10 cm DBH (Fig. 8). S. argentea and S. wallichii respectively reached 75 cm DBH, while S. khasiana, S. villosa, S. sinensis and S. sericans reached 55 cm DBH, respectively. The DBH-class frequency distribution of the major species of each secondary forest type is provided in Supplementary Table S2. Many small trees of non-Schima canopy species were also found in the secondary forests.
Seedlings/saplings of these seven Schima species were mainly found in well-lit sites including canopy gaps, forest edges and roadsides (Fig. 9). Canopy gaps contributed the most to the establishment of seedlings/saplings.However, S. noronhae, S. khasiana and S. wallichii had some established seedlings/saplings under canopies. The dense coverage (80%) of Yushania polytricha of bamboo in the understory of the forest Type PF1 impeded the regeneration of S. noronhae. Notably, abundant seedlings/saplings of S. wallichii and S. argentea were found, while the numbers of seedlings/sapling of S. sinensis, S. villosa,S. sericans were rather limited (Fig. 9). Since there were more canopy gaps in the secondary forests than the primary forests, the regeneration of these Schima species in the secondary forests had much better recruitments than those in the primary forests.
Discussion
Forest characteristics
Our study demonstrates that Schima is often codominated with or associated with various species of Castanopsis(e.g.Castanopsis ceratacantha,C.echinocarpa,C.platyacantha and C. hystrix) in the evergreen broadleaved forests of Yunnan.In fact,it is also true in other regions of China and East Asia, as exemplified by Schima superba associated with Castanopsis carlesii in Tiantong,Zhejiang (Da et al. 2004; Yang et al. 2006; Song 2007),associated with Castanopsis eyrei in Gutianshan, Zhejiang(Zhu et al. 2008) and Wuyishan, Jiangxi (Cai and Song 1997), associated with Castanopsis chinensis on Mt.Dinghu, Guangdong (Huang et al. 2000), associated with Castanopsis fabri and Castanopsis fargesii in Wanmulin,Fujian (Huang et al. 1997), associated with Castanopsis cuspidata var. carlesii in Taiwan (Tang et al. 2013).Schima argentea associated with Castanopsis carlesii var.spinulosa in Jinyunshan, Chongqing (Zeng et al. 2010).Schima sinensis associated with Castanopsis platyacantha on Mt.Emei,Sichuan(Yang et al.1994;Tang et al.2007).S. wallichii associated with Castanopsis sieboldii in Okinawa, southwestern Japan(Xu et al. 2008).S.wallichii associated with Castanopsis tribuloides in Bhutan Himalaya (Wangda and Ohsawa 2006), associated with Castanopsis indica in eastern Nepal Himalaya (Ohsawa 1986),and associated with Castanopsis lanceifolia in Meghalaya,NE India(Tripathi and Shankar 2014).
The Shannon-Wiener index H′ of our studied primary forests and secondary forests is 2.563±0.48 (average±SD) and 2.232±0.36, respectively. The indices of the forests in Yunnan are higher than those (1.768 for Dongbeishan, 1.566 for Fuzhou) of the Schima superba-Pinus massoniana forest in Dongbeishan, Zhejiang (Wang et al.2015) and in the National Forest Park of Fuzhou, Fujian(Cheng 2006), and higher than those (1.255-1.886) of Castanopsis platyacantha-Schima sinensis forest in Wawushan, Sichuan (Bao et al. 2000), also higher than that (1.72) of Schima superba-Castanopsis sclerophylla forest and that (2.19±0.46) of Schima superba forest in Tiangtong, Zhejiang (He et al. 1998), while the indices in Yunnan are lower than that (2.76) of Schima superba-Castanopsis carlesii in Tiantong(He et al.1998),Zhejiang and that (3.150) of Castanopsis chinensis-Schima superba in Dingfoshan, Guangdong (Huang et al. 2000). This is probably related to the different degrees of naturalness of the forests in the various study sites.
Ecological traits
The seven Schima species grow in a warm or mild climate with ample moisture and humidity in the study area. They mainly regenerate in canopy gaps formed under an existing gap disturbance regime (moderate anthropogenic and natural disturbances). Many of Schima species are shade-intolerant, and they more commonly dominate the early or middle successional forests (secondary forests). The long life-span of Schima species assures their presence in late successional forests (primary forests), and even co-dominate the canopy layer along with dominants of different genera. However, three species, i.e. S. noronhae, S. khasiana, S. wallichii are bimodal type species having shade-intolerant and somehow shade-tolerant traits since their seedlings/saplings were found in both well-lit and shady micro-sites.Schima species with winged seeds widely disperse by wind. After their regeneration is established, they will form a mosaic of forest stand structure across the landscape in the subtropical zone of Yunnan. The seven Schima species survive best in moderately disturbed sites. Much less seedlings/saplings of the three species i.e. S. sinensis, S. villosa and S. sericans were found than the other four species (S. wallichii, S. argentea, S. noronhae, S. khasiana), because the three species are found only in a few forest patches in Yunnan.
The Schima species under this study, ecological traits are similar to those of Schima superba in eastern China,which is a fast-growing tree with a higher growth rate during its youth (Wei 2017) and has a growth plasticity and a certain shade-tolerance, meanwhile forest canopy gaps support its seed germination and seedling/sapling establishment (Song 2007). The S. superba forest in Guangxi is an earlier type of recovering evergreen broad-leaved forest resulting from serial communities following destruction of original evergreen broadleaved forest through progressive succession. It will be replaced by Castanopsis species (Wang and Li 1999). In our study area of Yunnan, S. noronhae is found in the primary forests at the late successional stage as the top dominant,and either S. khasiana, S. wallichii, S. villosa or S.sinensis presents as one of the dominants in both late successional forests and early or middle-seral successional forests. However, S. argentea or S. sericans as one of dominants is only found in early or middle successional forests. As time goes by, the dominance of S.argentea and S. sericans would be replaced by their associated dominant taxa such as Castanopsis species(Supplementary Table S2).
In view of the potential effects of global climate warming on distribution, the Schima species under study may be forced to move to locations at higher latitudes or altitudes. Schima species in general have as strong dispersal capacity because of their lightweight and the fact that the seeds are wind-dispersed, so, they possibly can migrate to new sites. But they may not be able to evolve needed new adaptations to new localities in the time projected for a rapid climate change. Especially, Schima sericans endemic to NW Yunnan and SE Tibet and Schima villosa endemic to Yunnan and having a narrow distribution range will be greatly vulnerable under rapid global climate change.
Conclusions
Yunnan is found to harbor a high species richness and rarity-weighted richness of Schima. Canopy gaps created by moderate human and natural disturbances have supported Schima species’ regeneration establishment in forming a mosaic of forest stand structure across the landscape in the subtropical zone of Yunnan. Some of the Schima species studied as a second dominant (rare as the top-dominant) present in the late-successional forests.Some of them are more often found as the top-dominant in early or middle successional forests,where as time goes by the dominance of Schima species would be replaced by associated dominant taxa such as Castanopsis species.Schima species can be used for restoration projects to restore native forests as part of the subtropical evergreen broad-leaved forest ecosystem.
Supplementary information
Supplementary informationaccompanies this paper at https://doi.org/10.1186/s40663-020-00244-1.
Additional file 1. Supplementary Table S1.DBH-classes of species (IV
≥1.5%) of each primary forest type.
Additional file 2. Supplementary Table S2. DBH-classes of major
species (RIV ≥1.5%) of each secondary forest type.
Abbreviations
DBH: Diameter at breast height; RIV:Relative importance value; SD: Standard deviation; H: Height
Acknowledgements
Not applicable.
Authors’contributions
C.Q.T. designed the study, analyzed the data and wrote the manuscript. P.-B.H organized and analyzed the data. S.L. identified the botanical specimens.C.Q.T., L.-Q.S.,P.-B.H., D.-S.H., Y.-F.L., M.-C. P., C.-Y. W.,X.-S. L, W.L. and Z.-Y.Z.conducted the fieldwork. All the authors contributed discussion to improve the manuscript.
Funding
The study is financially supported by Ministry of Science and Technology,China (Project No.2015FY210200-15).
Availability of data and materials
The datasets generated and/or analyzed during the current study are not publicly available due limitation from the organization provided the research fund but are available from the corresponding author on reasonable request.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Author details
1Institute of Ecology and Geobotany, College of Ecology and Environmental Science, Yunnan University, Kunming 650504, China.2Research Institute on Resource Insects, Chinese Academy of Forestry, Kunming 650224,China.
3Caojian Forest Farm of Yunlong Forestry Bureau, Yunlong County 672711,Yunnan, China.4Yunnan Academy of Forestry, Kunming 650201, China.5Tibet Agriculture and Animal Husbandry College, Linzhi 860000, China.
Received: 23 January 2020 Accepted: 24 April 2020

- Forest Ecosystems的其它文章
- Soil-plant co-stimulation during forest vegetation restoration in a subtropical area of southern China
- Addressing soil protection concerns in forest ecosystem management under climate change
- Effects of afforestation of agricultural land with grey alder(Alnus incana(L.)Moench)on soil chemical properties,comparing two contrasting soil groups
- Delineating forest stands from grid data
- Tree diversity effects on forest productivity increase through time because of spatial partitioning
- Mapping data gaps to estimate biomass across Brazilian Amazon forests