
基于模糊隶属函数法评价无花果品种抗寒性
2020-10-17 01:18房海灵梁呈元亓希武
江苏林业科技 2020年4期
曾 杰,房海灵*,梁呈元*,亓希武,于 盱,李 莉,柏 杨,郭 强
(1.江苏省中国科学院植物研究所,江苏 南京 210014; 2.句容市虎耳山无花果专业合作社,江苏 句容 212400 )
无花果(FicuscaricaL.)为桑科(Moraceae)榕属落叶小灌木,又称映日果、奶浆果、蜜果等,是人类最早驯化栽培的果树之一[1]。无花果果实肉质甜糯,风味独特,富含丰富的氨基酸、维生素、膳食纤维和多糖等成分,具有降血压、抗衰老、抗癌、提高机体免疫力等功能[2-5],深受广大消费者的喜爱。这些年来,全国多地已陆续出现了种植无花果的热潮[6]。然而,低温却成了制约无花果安全越冬的关键因子。因此,研究无花果品种的抗寒性,对于栽培地品种选择,乃至耐寒品种的选育,都具有很重要的意义和价值。
目前,关于植物抗寒性的鉴定方法,主要有寒冻灾害调查法、生长恢复法、组织褐变法、生理生化指标测定法、组织细胞结构观察法等[7],其中生理生化指标测定法具有用时短、不受气候等条件限制等特点,已广泛应用于果树、绿化树种等植物(或作物)的抗寒性鉴定[8-9]。本研究拟对15个无花果品种进行人工低温处理,通过测定低温条件下它们的相对电导率、MDA等生理生化指标,利用模糊隶属函数法综合分析其抗寒性,为无花果引种地的品种选择及耐寒品种的选育工作提供借鉴与基础。
1 材料与方法
1.1 材料
采自江苏省中国科学院植物研究所无花果种质资源圃中2年生健壮、无病虫害、栽培管理措施一致的树体,种质名称、来源等信息见表1。

表1 供试材料
1.2 试验处理
于2019年12月取生长势接近、粗细一致的1年生休眠枝条,用于抗寒性试验。先用去离子水洗净枝条,吸干水分,剪成长10 cm左右的枝段,石蜡封住2端。每10段为1组,每个品种分5组,于低温冰箱中进行低温处理,即1,-6,-13,-20,-27 ℃共设置处理5个,每个处理重复3次。处理组以7 ℃/h速度降温,达到目的温度后保持24 h,取出后于1 ℃条件下解冻12 h,进行相关指标的测定。
1.3 指标检测
相对电导率测定:将低温处理枝条剪成长0.3 cm的片段,称取1 g置于50 mL试管,加入25 mL去离子水,于25 ℃摇床摇动90 min,测定初始电导率(R1),随后置于沸水浴中20 min,取出恢复室温后测定其终电导率(R2),计算相对电导率(%),即(R1/R2)×100。
MDA,Pro,可溶性糖、可溶性蛋白含量均采用南京建成生物科技有限公司生产的试剂盒完成测定。
1.4 数据统计与分析
采用Microsoft Excel 2010和SPSS22.0软件对试验数据进行计算分析,其中以处理温度x和相对电导率Y拟合Logistic方程Y=k/(1+ae-bx),k=100代表细胞伤害率的饱和容量,求得拟合度R2和参数a,b,再根据LT50=lna/b得到低温半致死温度(LT50)[10]。抗寒性综合指数采用模糊隶属函数法综合评价得到[11]。隶属函数值Uij=(Xij-Xjmin)/(Xjmax-Xjmin),反隶属函数值=1-(Xij-Xjmin)/(Xjmax-Xjmin),其中Xij为i品种第j个指标测定值,Xjmin和Xjmax为所有参试品种j指标最小测定值和最大值,j表示某抗寒指标。抗寒性综合指数=(Uij+Uik…+Uin)/n,n为指标个数。
2 结果与分析
2.1 低温胁迫下无花果品种相对电导率及半致死温度
植物受到低温胁迫时易出现质膜损伤现象,胞内电解质会发生不同程度的渗漏。因此,通常以电解质外渗率作为评价植物受冻害程度的指标之一[12]。图1表明,低温下无花果各品种枝条的相对电导率出现3个阶段的变化。在处理阶段Ⅰ(1—-6 ℃)时,部分品种相对电导率出现下降趋势。在处理阶段Ⅱ(-6—-20 ℃)时,所有测试品种相对电导率急剧上升。在处理阶段Ⅲ(-20—-27 ℃)时,部分品种相对电导率变化趋于平缓。其原因可能是,在低温初期,部分品种细胞膜功能未受到损伤,可通过减少电解质外运来降低冰点,以适应低温环境;随着胁迫温度进一步降低,质膜损伤,导致大量电解质渗漏,相对电导率急剧上升;达到极限温度时,质膜已受到不可逆损伤,电解质外渗趋于缓慢。
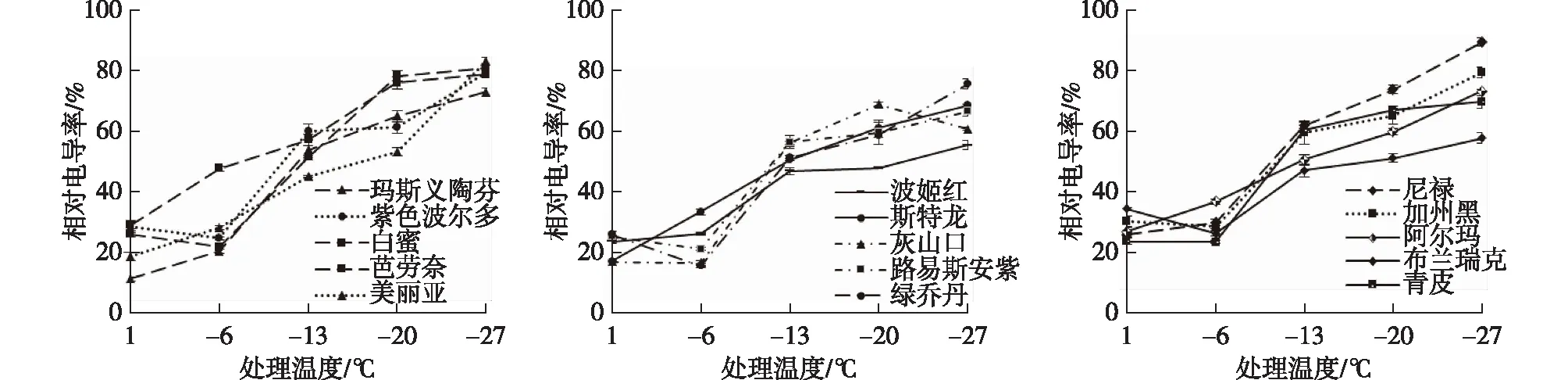
图1 低温胁迫对无花果枝条相对电导率的影响
利用Logistic回归方程,分析并计算供试无花果品种低温半致死温度LT50(见表2)。结果表明,Logistic回归方程可较好地对低温胁迫下无花果品种相对电导率与处理温度的关系进行拟合,拟合度为0.816—0.996,达到显著差异水平。15个品种LT50在-10.034—-17.538 ℃范围内,其中,抗寒能力最差的为尼禄,LT50为-10.034 ℃;抗寒能力最强的是波姬红,LT50为-17.538 ℃;其后抗寒能力强弱依次为布兰瑞克>灰山口>芭劳奈>斯特拉>路易斯安紫>青皮>玛斯义陶芬>白蜜>绿乔丹>美丽亚>紫色波尔多>加州黑>阿尔玛>尼禄。

表2 供试无花果品种的低温半致死温度(LT50)
2.2 低温胁迫对无花果生理活性的影响
2.2.1 低温胁迫对无花果MDA含量的影响 MDA是衡量植物逆境胁迫的重要指标,其含量的高低代表细胞受损程度[13]。由图2可知,低温处理下无花果各品种枝条中MDA含量均呈上升趋势,且上升幅度具有品种差异性。处理温度1—-6 ℃时,尼禄、路易斯安紫和芭劳奈等品种枝条中MDA上升趋势较为缓慢,而其余品种则对早期低温反应较为灵敏,MDA含量急剧升高;处理温度-6—-13 ℃时,白蜜、芭劳奈、波姬红、绿乔丹、加州黑、阿尔玛和尼禄中MDA含量上升幅度平缓外,其余品种均急剧上升;处理温度-13—-20 ℃时,各品种MDA含量上升幅度不同,波姬红、斯特拉、绿乔丹和尼禄等品种MDA含量急剧增加;在处理温度-20—-27 ℃时,除玛斯义陶芬、美丽亚、斯特拉和布兰瑞克外,其余品种中MDA含量均急剧增加,表明细胞膜受到严重损伤。
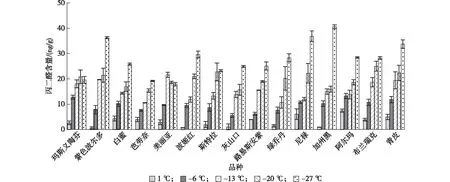
图2 低温胁迫对无花果品种MDA含量的影响
2.2.2 低温胁迫对无花果中游离脯氨酸含量的影响 低温胁迫下,植物通过提高细胞内游离脯氨酸(Pro)、可溶性蛋白和可溶性糖等渗透调节物质含量,维持细胞内环境稳定,进而减轻低温对植物体的损伤。由图3可见,低温处理下无花果品种枝条中Pro含量均表现为一定的上升趋势,表明各品种应对低温胁迫表现出积极的应激反应。尼禄和加州黑表现为Pro含量随着胁迫温度的降低呈先上升后降低的趋势,在-20 ℃时达到最大值。分析原因可能是低于一定温度时,植物体内Pro合成受到阻碍造成含量下降;其他品种则表现为随温度降低而升高,芭劳奈、波姬红和灰山口在-20—-27 ℃间急剧上升,可能是由于极低温度对植物脯氨酸合成酶基因表达诱导,促进了脯氨酸的合成。阿尔玛、绿乔丹、尼禄和加州黑Pro含量总体呈较低的水平,表明该类品种对低温胁迫的应激反应较弱。
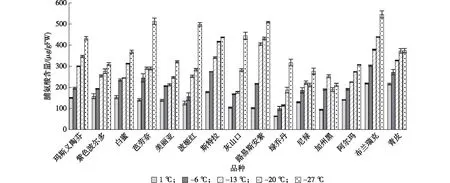
图3 低温胁迫对无花果品种Pro含量的影响
2.2.3 低温胁迫对无花果中可溶性糖含量的影响 由图4可见,多数无花果品种枝条中可溶性糖含量随着处理温度的降低而呈增高的趋势,如玛斯义陶芬、紫色波尔多、白蜜、芭劳奈、斯特拉、尼禄和青皮等品种在处理初期可溶性糖含量呈相对较低水平,之后随着处理温度的降低而稳定上升,处理温度-27 ℃时达到最高值;而灰山口、路易斯安紫、加州黑、青皮和阿尔玛等品种则随着处理温度的降低表现为先升高后降低的趋势,其原因可能是该类品种在极度低温条件下,可溶性糖合成相关酶因低温出现失活现象,影响了可溶性糖的合成;美丽亚和尼禄则表现出与以上2种不同的变化趋势,即在随着处理温度的降低呈倒“M”变化,分析原因可能是可溶性糖是能量的来源,植物通过消耗可溶性糖来促进其他渗透调节物质的合成,而其他渗透调节物质为可溶性糖的合成提供稳定的环境,促进了其积累。
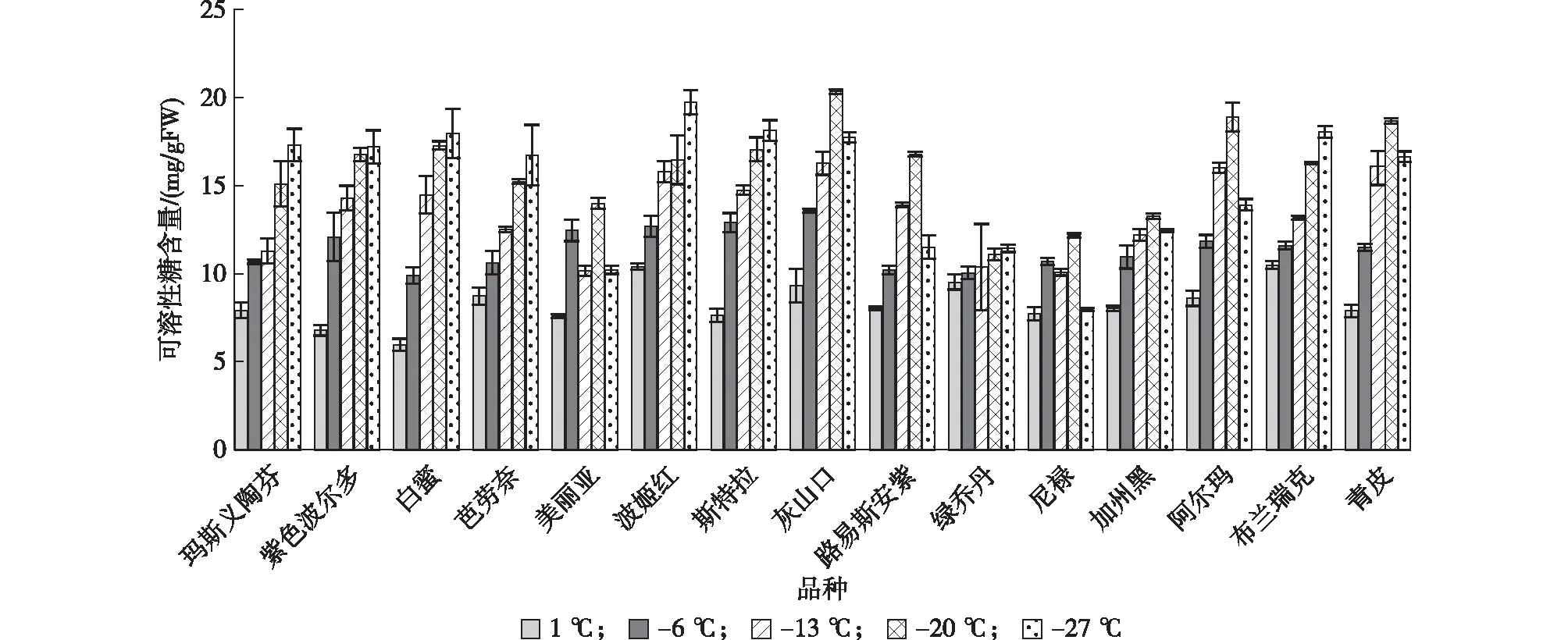
图4 低温胁迫对无花果品种可溶性糖含量的影响
2.2.4 低温胁迫对无花果中可溶性蛋白含量的影响 可溶性蛋白具有较强的亲水性,可作为渗透调节物质维持细胞渗透势,降低冰点,增强植株耐寒性。由图5可知,低温胁迫下无花果品种枝条中可溶性蛋白含量变化趋势与可溶性糖相似。玛斯义陶芬、美丽亚、白蜜、波姬红、路易斯安紫和加州黑等品种可溶性蛋白含量随温度降低而升高;灰山口和阿尔玛则表现为先上升后下降趋势;紫色波尔多、芭劳奈、斯特拉、绿乔丹、尼禄、布兰瑞克和青皮等品种可溶性蛋白含量随温度升高先升高后下降再升高。尼禄和加州黑在整个处理温度区间均表现为较低水平,表明该类品种抵御低温的能力较弱。
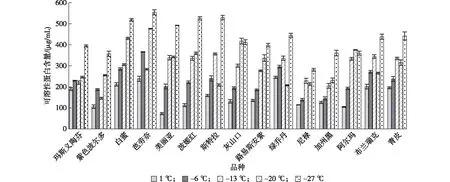
图5 低温胁迫对无花果品种可溶性蛋白含量的影响
2.3 无花果品种抗寒性综合评价
采用模糊隶属函数法对相对电导率、MDA、Pro、可溶糖和可溶性淀粉等5个能反映无花果抗寒能力的指标进行分析,并计算平均值,其中Pro、可溶糖和可溶性淀粉函数值采用隶属函数公式计算,相对电导率和MDA则采用反隶属函数计算,结果见表3。按照隶属函数值大小,对供试无花果品种的抗寒性进行排序,依次为波姬红>布兰瑞克>灰山口>斯特拉>芭劳奈>路易斯安紫>青皮>玛斯义陶芬>白蜜>美丽亚>绿乔丹>阿尔玛>紫色波尔多>加州黑>尼禄。
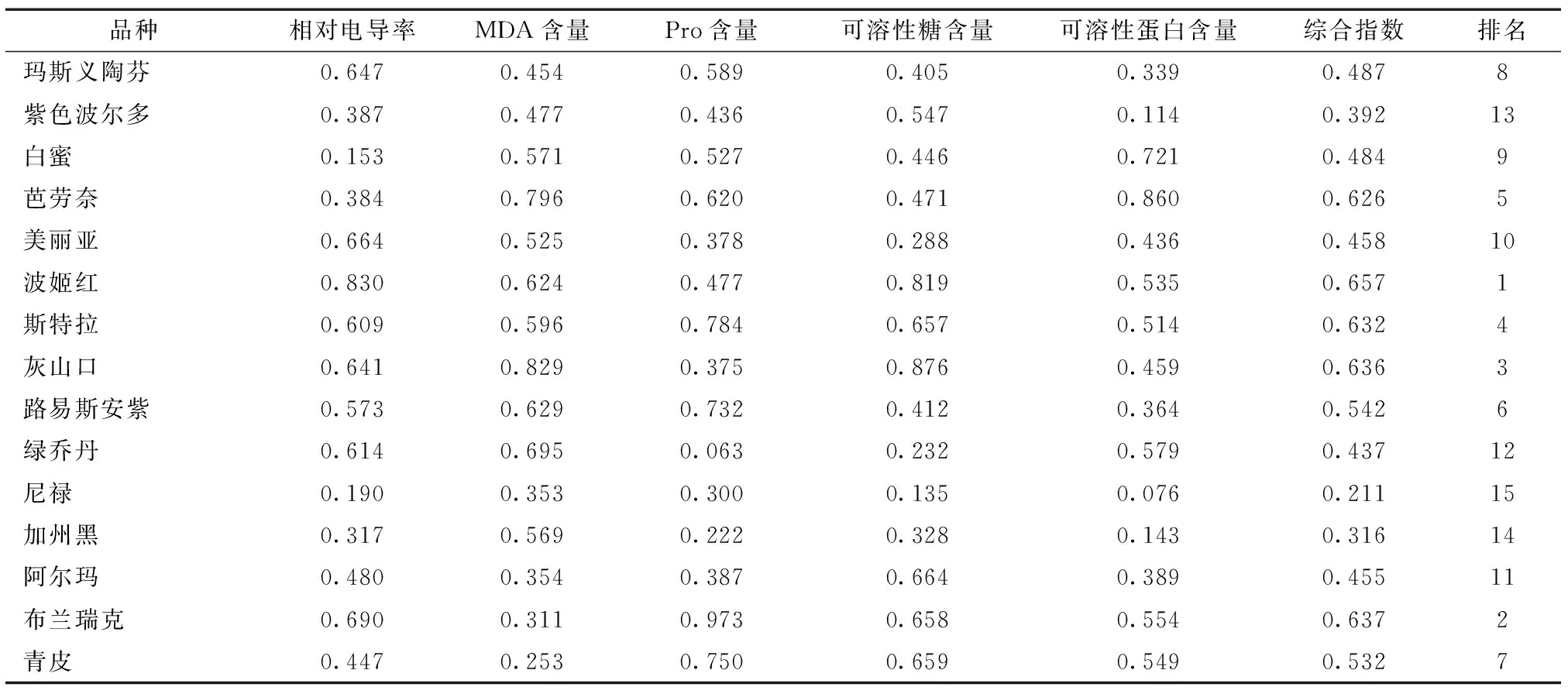
表3 无花果品种抗寒性指标隶属度值及综合指数
3 讨论
低温冻害是常见的自然灾害,一定程度上限制了植物的生长、发育及分布。研究表明,冻害可通过破坏生物质膜结构和功能对植株造成伤害。相对电导率是衡量膜透性的重要指标,值越高表示膜受损程度越严重。大量研究表明,植物在受到低温胁迫时相对电导率变化呈先缓慢增加后急剧上升最后趋于平缓的“S”型曲线,且开始急剧上升时所处温度因植物种类不同而各异,如桃为-21 ℃[10],苹果为-30 ℃[14],火龙果为4 ℃[15]。本研究表明,大部分参试无花果品种在处理温度为-6—-20 ℃间相对电导率呈激增状态,表明无花果开始激增的温度为-6 ℃。根据相对电导率计算所得的低温半致死温度被用来反映植物的抗寒性,并在多种植物如桃树[10]、苹果树[14]、火龙果[15]等树种中得到验证。古丽尼沙·卡斯木等对无花果品种抗寒性进行研究,表明供试11个品种LT50均在-12.62 ℃以下[16]。本研究亦表明80.0%种质枝条LT50在-15 ℃以下,与其起源于“古波斯和小亚细亚,即今天的土耳其、伊朗和叙利亚等地中海区域”相符[17]。
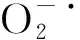
游离脯氨酸、可溶性蛋白和可溶性糖含量是衡量植物抗寒性的重要指标。研究表明,植物在低温胁迫下会合成大量的游离脯氨酸、可溶性蛋白和可溶性糖等渗透调节物质来保持细胞渗透势水平,为膜内物质提供稳定的环境,进而提高其对寒冷的适应性[22]。李桂荣等[20]研究表明,低温条件下葡萄中游离脯氨酸、可溶性蛋白和可溶性糖含量均有所提高。杨捷等[23]比较天目琼花和花叶锦带抗寒性发现,花叶锦带可溶性蛋白和脯氨酸含量均高于天目琼花,超过可承受的范围时,出现下降趋势,结合MDA等指标综合评价认为花叶锦带抗寒性强于天目琼花。本研究结果亦发现,低温诱导了无花果枝条中游离脯氨酸、可溶性蛋白和可溶性糖的合成以抵御低温对细胞膜造成的损伤,极限低温条件对部分品种可溶性糖和可溶性蛋白的合成具有抑制作用,该现象与张昕欣[24]在腊梅中的研究结果相似。分析原因可能是,第一, 极限低温抑制了可溶性糖和可溶性蛋白合成相关酶的活性,使得可溶性糖和可溶性蛋白的合成减弱,从侧面抑制了脯氨酸参与蛋白质合成的过程,导致脯氨酸的大量积累[25]。第二,可溶性糖是植物的能源物质之一,在极限低温下植物需要消耗更多的能量来抵御冷害胁迫,从而引起可溶性糖含量的下降。此外,抗寒性较强的无花果品种中3种渗透调节物质含量上升幅度大,而抗寒性弱的品种则上升幅度较小,这与王孝宣等[26]和吴行昶[25]在番茄、葡萄中的研究结果相一致。
模糊隶属函数法通过计算测定指标加权平均值来全面系统的反映评价指标,广泛应用于植物抗逆性评价[27-28]。翟新秘等[27]利用隶属函数分析法评价玉米抗旱性,得出的抗旱评级与干旱处理后玉米表型结果一致。罗尧幸等[28]基于隶属函数法对鲜食葡萄品种抗寒性进行评价,认为该方法优于单一指标评价法,能够全面反映葡萄实际抗寒能力。本研究采用隶属函数法对无花果品种抗寒性进行评价,结果发现波姬红的平均隶属函数值最大(0.657),抗寒性相对最强,与LT50结果相符;绿乔丹、阿尔玛、紫色波尔多、加州黑和尼禄等品种平均隶属函数值相对较低,表明其抗寒能力较弱。综上可见,隶属函数法能更为全面反映无花果品种的抗寒能力,可为无花果抗寒品种的选择与育种提供一定的参考。
猜你喜欢
中老年保健(2022年3期)2022-11-21
热带作物学报(2022年5期)2022-06-01
矿冶工程(2021年4期)2021-09-15
棉花学报(2020年3期)2020-08-08
科学与财富(2019年15期)2019-10-21
浙江工业大学学报(2017年5期)2018-01-22
现代园艺(2017年23期)2018-01-18
中国塑料(2016年11期)2016-04-16
中国果菜(2016年9期)2016-03-01
中国马铃薯(2015年5期)2016-01-09
