
北虫草刺五加组合物抗疲劳活性及机理研究
2020-10-14 09:31盛瑜白丽丹段懿涵安丽萍郭笑王曼力
食品研究与开发 2020年20期
盛瑜,白丽丹,段懿涵,安丽萍,郭笑,王曼力
(北华大学药学院,吉林吉林132013)
随着生活节奏的加快,越来越多的人处于亚健康状态,亚健康状态的众多临床表现中以“无法解释的疲劳”最为常见。疲劳常会导致一系列严重的继发性问题,如焦虑、抑郁、认知障碍、睡眠质量差、身体功能障碍、能量失衡,甚至能够引发与生物调节和免疫系统有关的严重疾病[1-2]。疲劳已成为威胁人类健康的一个重大问题。因此,通过有针对性地补充外源性营养补剂或药物以延缓或防止疲劳的发生,是解决疲劳以及调节亚健康状态的有效途径之一[3]。
疲劳不仅与现代生活方式引起的压力水平升高有关,而且通常伴随着机体运动能力下降[4]。正常的疲劳,通常发生在强烈的体力劳动之后,可视为一个机体保护信号,提示机体需要休息[5]。导致运动性疲劳的机制复杂,包括:代谢产物如乳酸(lactate,LA)和尿素氮(urea nitrogen,UN)的过量产生和积累导致肌肉衰竭;活性氧(reactive oxygen species,ROS)在肌肉或其他相关器官(如肝脏)中产生的氧化应激和损伤;肝脏和肌肉的血氧浓度平衡和糖原内稳态的改变降低了机体运动的可持续性和运动后的恢复等[6]。近年来的临床应用表明,一些复方或单方中草药通过消除上述代谢物、减少氧化应激,改善糖原代谢等途径预防和缓解运动性疲劳,可见中草药在抗疲劳和提高运动能力方面具有得天独厚的优势。
北虫草(Cordyceps militaris)作为药材和民间滋补食品在东亚地区广为应用,中医认为,其入肺肾二经,可补肺阴和肾阳。据报道,北虫草提取物(Cordyceps militaris fruit body extract,CM)及其多糖(polysaccharide from C.militaris,PCM)具有显著的抗疲劳功效[7-8]。刺五加(Acanthopanax senticosus)作为药物的应用历史十分悠久,在历代本草著作中均有记载,其具有“补中、益精、坚筋骨、强志意”的功效[9]。研究发现,刺五加茎皮提取物(extracts of stem bark from Acanthopanax senticosus,ASSE)具有抗疲劳作用,能提高运动耐力[10]。
本课题组基于现代医学对运动性疲劳的病因病机的研究,结合传统中医养生理论,将北虫草与刺五加进行配伍,制备了北虫草刺五加组合物(Cordyceps militaris Acanthopanax senticosus composition,CAC)。本研究通过检测小鼠疲劳转棒时间、力竭游泳时间及小鼠体内血乳酸(lactic acid,LA)、血乳酸脱氢酶(lactate dehydrogenase,LDH)、血尿素氮(blood urea nitrogen,BUN)、肝糖原(liver cogen,LG)、肌糖原(muscle glycogen,MG)等指标考察该组合物对小鼠的抗疲劳作用,并通过检测小鼠肝脏中总超氧化物歧化酶(total superoxide dismutase,T-SOD),丙二醛(malondialdehyde,MDA),活性氧(reactive oxygen species,ROS)的活性或含量,以及磷酸化-蛋白激酶B(phosphorylated-protein kinase B,p-AKT)、磷酸化-糖原合成酶激酶(phosphorylated-glycogen sythesis kinase-3,p-GSK3β) 和葡萄糖转运子 4(glucose transporter 4,GLUT4)蛋白的表达情况,探究北虫草刺五加组合物抗疲劳的可能作用机制,以期为该组合物作为抗疲劳功能性食品的开发提供依据。
1 材料与方法
1.1 材料与试剂
北虫草刺五加组合物(CAC):由北华大学药学院研发制备,其中原料药材北虫草购自沈阳聚鑫北虫草菌业有限公司、刺五加购自国药控股潍坊有限公司。
血清尿素氮(BUN)、乳酸(LA)、乳酸脱氢酶(LDH)、肝糖元(LG)、肌糖元(MG)、丙二醛(MDA)、总超氧化物歧化酶(T-SOD)试剂盒:南京建成生物工程研究所;活性氧簇(ROS)试剂盒:酶联生物公司;βactin、p-AKT(Ser 473)、p-GSK3β(Ser 9)、GLUT4 兔抗鼠单克隆抗体、辣根过氧化物酶标记山羊抗兔IgG(H+L):ABclone公司。
SPF级雄性ICR小鼠:4~5周龄,体重18 g~22 g,40只,实验动物许可证编号为SCXK(吉)-2016-0003,长春亿斯实验动物技术有限责任公司。
1.2 仪器与设备
紫外可见分光光度计(UV-2550型):SHIMADZU公司;酶标仪(Infinite M200型):TECAN公司;低温高速离心机(5810型):EPPENDORF公司;电子分析天平(ME55):METTLER TOLEDO 公司;自动疲劳转棒仪(ZB-200):成都泰盟软件有限公司。
1.3 方法
1.3.1 动物分组及给药
SPF级健康雄性ICR小鼠40只,适应性喂养1周,实验期间小鼠自由饮食、饮水,适应性喂养7 d后,按每组10只随机分成4组,并结合中药配伍组方剂量及人体给药剂量的5、10、20倍设计小鼠给药剂量,即空白对照组(记为CON)、北虫草刺五加组合物低剂量组[low dosage of CAC,LCAC,50 mg/(kg·d)]、北虫草刺五加组合物中剂量组 [medium dosage of CAC,MCAC,100 mg/(kg·d)]、北虫草刺五加组合物高剂量组[high dosage of CAC,HCAC,200 mg/(kg·d)],每天进行灌胃给药,其中CON组灌胃给药生理盐水,连续给药5周。每间隔10 d称重,观察给药后小鼠体重,精神状态,摄入食水量,毛色变化等情况。具体实验设计方案如图1。

图1 实验方案Fig.1 Experiment scheme
1.3.2 小鼠疲劳转棒实验
连续给药第29天时,给药30 min后,将各组小鼠置于转速为30 r/min转棒疲劳仪上,进行小鼠疲劳转棒训练,1次/d,连续3 d。第32天,给药30 min后,进行正式转棒实验,将小鼠放置于转速以1 r/min递增至30 r/min的电动转棒疲劳仪上,开启电源,以3 min内不落降为标准,记录小鼠的疲劳转棒时间[11]。
1.3.3 小鼠负重游泳实验
连续给药第35天时,末次灌胃给药30 min后,进行力竭负重游泳实验,对小鼠进行耐力评估。在小鼠尾部固定5%体重的铅线,并将各给药组小鼠轻轻置于水深40 cm、水温(25±2)℃的恒温游泳箱中,使其可以自由游泳。小鼠自游泳开始至完全没入水中7 s,不能露出水面用鼻子进行呼吸时,可认为小鼠达到力竭状态,需立即将其打捞,并记录小鼠力竭游泳时间[12]。
1.3.4 小鼠血清与组织的采集与制备
负重游泳小鼠在强迫游泳力竭后,休息30 min,腹腔注射20%的乌拉坦(10 mL/kg)进行麻醉,摘除眼球,收集眼球后血管丛血液,血样静置10 min后,放入低温高速离心机中,在4℃下,以3000r/min离心10min,取上清,重复1次,将两次上清合并,储存于-80℃冰箱中备用。
取血完毕后,以断髓方式处死小鼠,并快速解剖取出肝组织和腓肠肌,并制备10%组织匀浆:称重2 mg肝组织和腓肠肌组织,置于4 mL离心管内,按照料液比1∶9(g/mL)加入生理盐水,冰上充分匀浆。将组织匀浆液放入低温高速离心机中,在4℃下以3 000 r/min离心10 min取上清,储存于-80℃冰箱中备用。
1.3.5 生化指标检测
小鼠血清中 BUN、LA、LDH,肝脏组织中 LG、ROS、MDA、T-SOD,肌肉组织中MG均按照试剂盒说明书的方法测定。
1.3.6 Western Blot检测
小鼠肝 AKT、GSK3β、GLUT4蛋白表达水平采用Western Blot法测定。称取部分肝组织,加入蛋白裂解液,匀浆离心,提取总蛋白,并测定蛋白质浓度,分装样品,-20℃储存备用。样品蛋白质通过12%十二烷基硫酸钠-聚丙烯酰胺(sodium dodecyl sulfate polyacrylamide gel electrophoresis,SDS-PAGE)凝胶电泳分离,转模2 h至聚丙二氟乙烯(polyvinylidene fluoride,PVDF)膜,将PVDF膜置于含5%脱脂奶粉的三乙醇胺缓冲盐(tris buffered saline/tween,TBS-T)封闭液中封闭1 h,在4℃下一抗孵育过夜,用TBS-T洗5次,每次15 min,加入辣根过氧化物酶(horseradish peroxidase,HRP)标记的二抗,室温(10℃~30℃)孵育 1 h,TBS-T洗5次,15min/次,按照化学发光(electrochemiluminescence,ECL)试剂盒的说明进行曝光,内参βactin蛋白与目的蛋白灰度的比值即为目的蛋白的相对表达量。
1.3.7 数据统计分析
2 结果与分析
2.1 北虫草刺五加组合物对小鼠转棒时间的影响
疲劳转棒实验可直观体现药物对实验小鼠耐力和平衡性的影响,各组小鼠在转棒仪上停留时间结果如表1所示。
表1 不同样品对小鼠转棒停留时间的影响(n=10,±s)Table 1 Effects of different samples on the rotating rod lasting time in mice(n=10,±s)

表1 不同样品对小鼠转棒停留时间的影响(n=10,±s)Table 1 Effects of different samples on the rotating rod lasting time in mice(n=10,±s)
注:给药组与正常对照组对比,**表示差异显著,P<0.01。
组别剂量/(mg/kg)停留时间/s CON 组 49.8±7.8 LCAC 组 50 84.5±7.4**MCAC 组 100 92.1±8.1**HCAC 组 200 96.8±4.4**
与CON组相比,LCAC组小鼠停留时间延长至(84.5±7.4)s,MCAC组小鼠停留时间进一步延长至(92.1±8.1)s,HCAC组小鼠停留时间达到各组最高值(96.8 ±4.4)s,并差异显著(P<0.01),由此可见,北虫草刺五加组合物对小鼠转棒的停留时间提高较为明显,同时具有剂量依赖性,该组合物具有一定的抗疲劳作用。
2.2 北虫草刺五加组合物对小鼠负重游泳时间的影响
运动耐力可直接反映药物或保健食品的抗疲劳活性,因此,小鼠力竭游泳试验被广泛用于抗疲劳活性功效检测[13]。北虫草刺五加组合物对实验小鼠负重游泳时间的影响如表2所示。
表2 不同样品对小鼠负重游泳时间的影响(n=10,±s)Table 2 Effects of different samples on loading swimming time in mice(n=10,±s)

表2 不同样品对小鼠负重游泳时间的影响(n=10,±s)Table 2 Effects of different samples on loading swimming time in mice(n=10,±s)
注:给药组与正常对照组对比,**表示差异显著,P<0.01。
组别剂量/(mg/kg)停留时间/s CON 组 31.0±3.1 LCAC 组 50 47.5±4.7**MCAC 组 100 50.9±5.3**HCAC 组 200 52.9±3.8**
与CON组相比,LCAC组小鼠负重游泳时间延长至(47.5±4.7)min,MCAC组小鼠负重游泳时间进一步延长至(50.9±5.3)min,HCAC组小鼠负重游泳时间达到各组最高值(52.9±3.8)min,并差异显著(P<0.01)。结果表明,北虫草刺五加组合物可以延长小鼠力竭游泳时间,具有抗疲劳活性,且中、高剂量组抗疲劳活性优于低剂量组。
2.3 北虫草刺五加组合物对小鼠抗疲劳相关生化指标的影响
研究发现,高强度运动能够扰乱机体的抗氧化系统,同时,脂质过氧化能够改变细胞膜的流动性从而阻碍蛋白质等营养物质的传输,继而影响肌肉细胞的正常运作,导致疲劳产生[12],因此,考察物质体内抗氧化作用的强弱可间接体现其是否具有抗疲劳作用。自由基损伤是机体产生疲劳的重要原因之一,ROS是细胞内主要的氧自由基,是机体有氧代谢过程中的一种副产物,能反映细胞内氧化系统的情况[14]。MDA是机体自由基诱导的脂质过氧化的最终产物,其含量可反映细胞发生氧化损伤的程度,此外,机体长期处于疲劳状态时,MDA的含量随之升高,目前,MDA是评价疲劳状况最常用的指标之一[15]。T-SOD是细胞内重要的抗氧化酶类,T-SOD可通过降低机体内自由基的活性,从而有效地减弱其对机体的破坏,T-SOD活性的提高能够延缓运动性疲劳[16]。本研究通过测定T-SOD活性、ROS和MDA的含量,观察北虫草刺五加组合物对疲劳小鼠作用,结果见表3。
表3 不同样品对小鼠肝组织氧化相关指标的影响结果(n=10,±s)Table 3 Effects of different samples on liver oxidation-related indicators(n=10,±s)
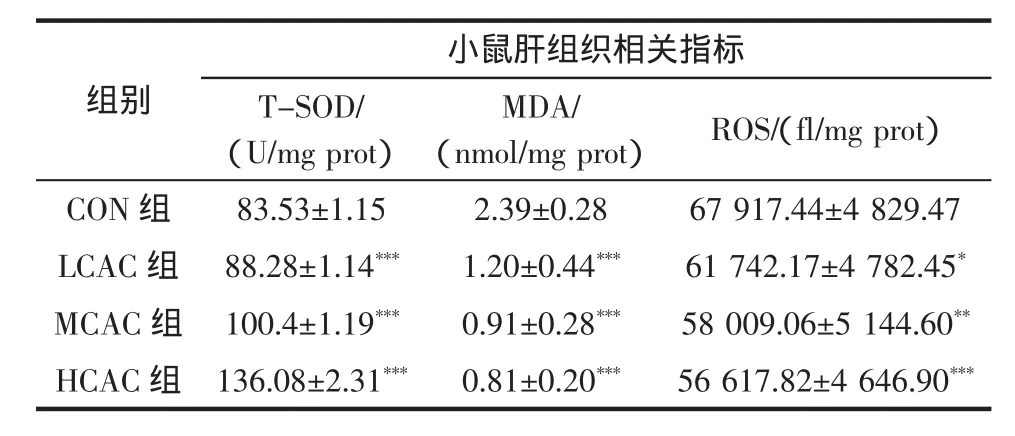
表3 不同样品对小鼠肝组织氧化相关指标的影响结果(n=10,±s)Table 3 Effects of different samples on liver oxidation-related indicators(n=10,±s)
注:给药组与正常对照组对比,*表示有差异P<0.05;**表示差异显著P<0.01;***表示差异极显著P<0.001。
组别小鼠肝组织相关指标T-SOD/(U/mg prot)MDA/(nmol/mg prot)ROS/(fl/mg prot)CON 组 83.53±1.15 2.39±0.28 67 917.44±4 829.47 LCAC 组 88.28±1.14*** 1.20±0.44*** 61 742.17±4 782.45*MCAC 组 100.4±1.19*** 0.91±0.28*** 58 009.06±5 144.60**HCAC 组 136.08±2.31*** 0.81±0.20*** 56 617.82±4 646.90***
由表3可知,灌胃北虫草刺五加组合物小鼠运动后T-SOD水平明显高于CON组,其中,LCAC组、MCAC组、HCAC组均极显著高于CON组(3组均P<0.001),而灌胃北虫草刺五加组合物小鼠运动后MDA水平均明显低于CON组,其中,LCAC组、MCAC组、HCAC组均极显著低于CON组(3组均P<0.001),此外,灌胃北虫草刺五加组合物小鼠运动后ROS水平均低于CON组,其中,LCAC组低于CON组(P<0.05),MCAC组显著低于CON组(P<0.01),HCAC组极显著低于CON组(P<0.001),呈剂量依赖性。以上实验结果说明北虫草刺五加组合物能够提高小鼠肝脏中TSOD活力,降低MDA和ROS的含量,一定程度上避免小鼠剧烈运动后氧化应激导致的细胞损伤,从而减轻小鼠的运动性疲劳。
本实验通过测定小鼠血清中BUN的含量,考察北虫草刺五加组合物对小鼠体内蛋白质和氨基酸代谢影响,进而观察其抗疲劳作用,结果见表4。
机体在生命活动中需要不断地消耗能量。糖原、脂肪和蛋白质是机体主要的能量物质,其中糖原在能量代谢中占主导位,蛋白质仅起辅助作用。糖原主要分布于骨胳肌和肝脏,其主要作用是在运动期间补充血糖消耗,以维持机体血糖稳定。剧烈运动时,血糖转移至肌肉,形成肌糖原,肌糖原分解为肌肉运动提供能量,从而导致血糖耗尽,此时,肝糖原氧化分解为葡萄糖,并释放入血,继而提供血糖,据报道,肝糖原被耗尽时,机体会发生体力疲劳[17],因此,肌糖原和肝糖元可作为反映机体疲劳程度的相关指标。此外,机体在剧烈运动时,肌肉组织供氧不足,葡萄糖需通过无氧酵解供能,乳酸(lactic acid,LA)是糖酵解过程的终产物,LA堆积使机体血液pH值降低,进而引起一系列生化改变,如神经肌肉结点处兴奋传递受阻,形成肌肉酸痛,最终产生运动性疲劳[18]。LA的堆积取决于其产生和消除的速度,LA可在乳酸脱氢酶(lactate dehydrogenase,LDH)的作用下氧化为丙酮酸并进入线粒体进行三羧酸循环,从而进行消除,减少LA的堆积,减缓机体疲劳[19]。因此,通常用血清中LA水平和LDH活性反映机体无氧代谢能力、疲劳的产生和消除速度。综上,本实验通过测定小鼠肝脏中肝糖原(liver cogen,LG)以及腓肠肌中肌糖原(muscle glycogen,MG)的含量,以及血清中LA和LDH的水平,考察北虫草刺五加组合物对小鼠体内糖原代谢影响,进而观察其抗疲劳作用。
表4 不同样品对小鼠血清、肝组织、腓肠肌抗疲劳相关指标的影响结果Table 4 Effect of different samples on anti-fatigue related indexes of mice serum,liver tissue and gastrocnemius muscle(n=10,±s)

表4 不同样品对小鼠血清、肝组织、腓肠肌抗疲劳相关指标的影响结果Table 4 Effect of different samples on anti-fatigue related indexes of mice serum,liver tissue and gastrocnemius muscle(n=10,±s)
注:给药组与正常对照组对比,*表示有差异P<0.05;**表示差异显著P<0.01;***表示差异极显著P<0.001。
组别 血清指标 肝脏LG/(mg/g)腓肠肌MG/(mg/g)LA/(mmol/L)LDH/(U/L)BUN/(mmol/L)CON 组 5.10±0.37 526.74±6.07 20.3±1.48 13.55±0.45 3.48±0.37 LCAC 组 4.53±0.55* 538.42±2.68*** 17.18±0.61*** 15.10±0.57*** 4.1±0.36**MCAC 组 3.65±0.39*** 541.60±4.89*** 15.08±1.00*** 15.51±0.41*** 5.27±0.34***HCAC 组 3.02±0.42*** 543.47±5.34*** 13.95±0.65*** 16.28±0.43*** 6.09±0.44***
由表4可知,灌胃北虫草刺五加组合物低、中、高剂量小鼠运动后肝糖原LG水平均极显著高于CON组(3组均P<0.001),同时灌胃北虫草刺五加组合物各组小鼠运动后肌糖原MG水平均明显高于CON组,其中,LCAC组显著高于 CON组(P<0.01)、MCAC组、HCAC组均极显著高于CON组(两组均P<0.001)。此外,灌胃北虫草刺五加组合物低、中、高剂量小鼠运动后血LA水平均低于CON组,其中,LCAC组低于CON组(P<0.05)、MCAC组、HCAC组均极显著低于CON组(两组均P<0.001);灌胃北虫草刺五加组合物低、中、高剂量小鼠运动后血清LDH水平均极显著高于CON组(3组均P<0.001)。以上实验结果说明,北虫草刺五加组合物能够提高小鼠肝糖原和肌糖原的储备,为其剧烈运动补充能量,减缓疲劳发生,同时,可通过提高机体LDH活性,减少无氧糖酵解产物LA的堆积,从而缓解其疲劳程度。
当机体不断地消耗能量,糖原和脂肪分解代谢不足以提供充足的能量时,蛋白质分解代谢供能相应增强,蛋白质和氨基酸在分解代谢中产生氨并在肝脏中转变为尿素,尿素氮进入血液循环形成血尿素氮(blood urea nitrogen,BUN),最终从肾脏排出体外。BUN含量的增加,可降低机体对负荷适应性。据报道,运动后BUN清除越快,则疲劳感消除越快[20],因此,BUN水平是判断机体疲劳程度的主要指标之一。
由表4可知,灌胃北虫草刺五加组合物低、中、高剂量小鼠运动后BUN水平均极显著低于CON组(3组均P<0.001)。本实验结果表明,北虫草刺五加组合物对于运动所造成的小鼠BUN水平异常升高有良好的调节作用,可促进机体在运动时维持正常的身体机能,并在运动后消除疲劳。
2.4 北虫草刺五加组合物对小鼠肝脏中糖代谢相关蛋白表达的影响
北虫草刺五加组合物对小鼠肝脏中糖代谢相关蛋白表达的影响见表5和图2。
表5 不同样品对小鼠肝组织中蛋白表达的影响Table 5 Effect of different samples on the expression of proteins in liver of mice(n=10,±s)
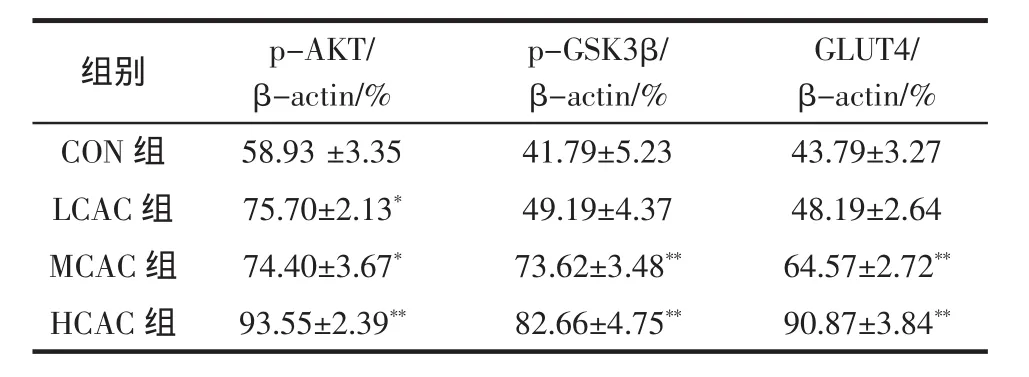
表5 不同样品对小鼠肝组织中蛋白表达的影响Table 5 Effect of different samples on the expression of proteins in liver of mice(n=10,±s)
注:给药组与正常对照组对比,*表示有差异P<0.05,**表示差异显著P<0.01。
GLUT4/β-actin/% CON 组 58.93 ±3.35 41.79±5.23 43.79±3.27 LCAC 组 75.70±2.13* 49.19±4.37 48.19±2.64 MCAC 组 74.40±3.67* 73.62±3.48** 64.57±2.72**HCAC 组 93.55±2.39** 82.66±4.75** 90.87±3.84**组别 p-AKT/β-actin/% p-GSK3β/β-actin/%

图2 不同样品对小鼠肝组织中蛋白表达的影响(n=10,±s)Fig.2 Effects of different samples on expression of proteins in liver of mice(n=10,±s)
在肝糖原合成的调控中,糖原合成酶(glycogen synthase,GS)发挥重要作用,其中,通过胰岛素介导的PI3K/AKT信号通路可激活GS,进而促进糖原合成。PKB/AKT的活化能够增加其下游糖代谢的负调节GSK3β的磷酸化,抑制其活性,并进一步解除GSK3β对于GS的抑制,从而增加GS的活性,促进肝细胞糖原合成[21]。此外,激活的AKT能够增加细胞膜中的GLUT4的转位,促进肝细胞对糖的吸收[22]。因此,PI3K/AKT/GSK3β信号通路的激活及GLUT4的膜转位可促进肝脏中糖原的合成,减缓机体疲劳程度。本实验通过测定小鼠肝脏中 p-AKT(Ser 473)、p-GSK3β(Ser 9)以及GLUT4蛋白的表达,初步探索考察北虫草刺五加组合抗疲劳的机制。
由表5、图4可知,灌胃北虫草刺五加组合物低、中、高剂量小鼠运动后肝脏中p-AKT(Ser 473)、p-GSK3β(Ser 9)以及GLUT4蛋白的表达量均有所增加,其中LCAC和MCAC组p-AKT(Ser 473)蛋白表达均高于 CON 组(P<0.05),HCAC 组 p-AKT(Ser 473)蛋白表达显著高于CON组(P<0.01);MCAC和HCAC组p-GSK3β(Ser 9)和GLUT4蛋白表达均显著高于CON组(P<0.01),LCAC组与CON组无差异。本实验结果表明,北虫草刺五加组合物能够通过促进AKT/GSK蛋白的磷酸化,提高GLUT4的表达,促进肝脏中糖原的合成,减缓疲劳发生。
3 结论
综上所述,北虫草刺五加组合物具有抗疲劳功效,能够延长小鼠转棒的停留时间和力竭游泳时间,并通过提高肝脏中T-SOD活力,降低MDA和ROS的含量,减轻机体因高强度运动引起的氧化应激损伤,通过增加肝糖原和肌糖原储备,提高机体LDH活性,降低LA堆积,以及降低BUN产生,减缓疲劳发生。机制研究显示,北虫草刺五加组合物能够通过激活PI3K/AKT信号通路,促进GSK3β磷酸化,并增加GLUT4的表达,提高肝原糖的合成,从而发挥抗疲劳的作用。
猜你喜欢
广西糖业(2022年5期)2022-11-24
今日农业(2022年15期)2022-09-20
实用肿瘤学杂志(2019年5期)2019-02-10
天然产物研究与开发(2018年7期)2018-08-21
中成药(2018年3期)2018-05-07
成都体育学院学报(2017年1期)2017-02-21
中成药(2016年4期)2016-05-17
药学与临床研究(2015年4期)2015-06-05
特产研究(2014年4期)2014-04-10
食品工业科技(2014年13期)2014-03-11
