
藏北3种高寒草地植物根系碳氮磷密度的非生长季和生长季差异
2020-09-24 08:25马星星鄢燕洪江涛王小丹
生态环境学报 2020年7期
马星星,鄢燕,洪江涛,王小丹*
1. 山西师范大学地理科学学院,山西 临汾 041004;2. 中国科学院水利部成都山地灾害与环境研究所山地表生过程与生态调控重点实验室,四川 成都 610041
随着对碳(C)排放的重视,中国进行了大量生态系统C储量的研究,主要集中在森林(王效科等,2001)和草地生态系统(Fan et al.,2008;高添等,2012)。对中国生态系统氮(N)磷(P)的研究主要集中于土壤N、P库的估算(Yang et al.,2007),对植被N、P的研究主要集中在叶片尺度,对植物N、P库的研究目前还较少。C作为植物的结构性物质,受环境的影响较小,含量较为稳定。氮(N)、磷(P)在生态系统中的储量和分配受多种因素的影响(范燕敏等,2018),如温度、水分和养分等,同时也影响到植物生长和生态系统功能。退化程度的加剧和生长季的推移,使根系P元素含量逐渐降低(聂志刚等,2018)。随着植物的生长,普遍认为植物组织中N、P元素含量会有一个稀释效应(Sardans et al.,2008)。根系是植物的重要组成部分,伴随着根系的生长和凋落而产生的C、N、P养分循环对生物地球化学循环有重要的生态学意义。高寒草原、高寒草甸草原和高寒草甸是西藏地区最主要的草地类型,3种草地面积占西藏草地总面积的77%以上(王建林等,2009)。由于干旱及寒冷等极端环境条件,植物将更多生物量分配于根系(胡中民等,2005;朴世龙等,2004)。N、P是限制陆地生态系统初级生产力的关键因子(Hans et al.,2010),同时也影响生态系统的物种丰富度、群落组成,尤其在受低温限制的高寒和极地生态系统中更是如此(Bardgett et al.,2007)。石福孙等(2008)对亚高山草甸的研究表明:增温提高了根系的碳储量,却降低了根系的氮储量。围栏显著增加了海拔4500 m处9.8%的植物群落地下部分N含量,显著增加了海拔4700 m处 4.2%的植物群落地下部分 C含量(沈振西等,2016)。在全球变化大背景下,温度和降水格局的变化可能引起高寒植物C、N、P密度的改变,从而影响植物—土壤间的养分循环过程,因此认识高寒植物根系及其C、N、P密度具有重要的意义。
目前对高寒植物根系的研究多集中在生长季根系生物量及其垂直分布上(Li et al.,2011;Ohtsuka et al.,2008;Wu et al.,2013;Yang et al.,2008),对其 C密度的计算方法多基于使用地上生物量和NDVI数据进行估算(Fan et al.,2008;王建林等,2009)。Fan et al.(2008)对中国草地生态系统C储量的估值为3.32 Pg C,其中青藏高原区草地C储量占56.4%,北方温带草地占17.9%;草地植被平均C密度从沼泽湿地的2.400 kg·m-2到高寒荒漠草地的 0.149 kg·m-2。而国家草地调查数据基于根茎比和 NDVI数据估算中国草地植物 C密度为215.8—348.1 g·m-2,平均值为 300.2 g·m-2(方精云等,2010)。不同的估算方法,导致结果有很大的差异。常常因为缺乏实地植被C密度数据导致对C储量的估算不准确(Ni,2004),因此增加植被实测C、N和P密度研究非常必要。
本文以那曲地区申扎县高寒草地(高寒草原、高寒草甸草原和高寒草甸)为研究对象,通过2013年4月和8月的野外采样获得根系C、N、P密度数据,摸清3种高寒草地根系养分密度在非生长季和生长季的变化特征,并比较同一时期草地类型间根系养分密度的差异,探究草地根系养分密度垂直分布及时期差异,为研究高寒地区草地根系养分密度季节变化提供基础资料,对进一步认识草地在全球C、N循环和气候变化环境下的作用提供理论支持。
1 材料与方法
1.1 研究区概况
实验地点位于西藏自治区那曲地区申扎县内(30º57′N,88º42′E,海拔 4675 m)。该地区为高原亚寒带半干旱季风气候,寒冷干燥,空气稀薄,日照辐射强。年均气温、年均降水量和年均蒸发量分别为0 ℃、300 mm和2181.1 mm左右(张晓克等,2014),且绝大多数降水发生在生长季(5—9月)。霜期持续天数为279.1 d,年日照时数为2915.5 h。该区植被以高寒草原为主,也分布有一定面积的高寒甸状草原(以下简称为草甸草原)和高寒草甸,具体植物群落特征见表 1。高寒植被以紫花针茅(Stipa purpurea)、青藏苔草(Carex moocroftii)、矮火绒草(Leontopodium nanum)、棘豆(Oxytropis)、狼毒(Stellera chamaejasme)、青藏苔草、矮嵩草(Kobresia humilis)和大嵩草(Kobresia macrantha)为主,以及一些杂类草构成。高寒草原和高寒草甸的土壤类型依次为:低温干旱土和冰冻始成土(Cai et al.,2013)。草甸草原土壤介于高寒草原和高寒草甸土壤类型之间。
1.2 样品采集和分析
在试验区内自东向西挑选地势平坦、地形均一的高寒草原、草甸草原和高寒草甸(形成原因与距离水源的远近有关,高寒草原到高寒草甸由东向西分布,在高寒草甸的西边有申扎藏布和格仁措)样地各一块,3块样地一字排开,且相邻两块样地间距离约2 km。高寒草原样点位于申扎高寒草原与湿地观测站的草原站内,草甸草原和高寒草甸样点位于湿地站外。本研究分别在2013年4月(非生长季)和8月(生长季)进行根系样品采集,每种草地3次重复。根系生物量采用挖土块法,土块大小为50 cm×50 cm,深30 cm,每10 cm一层,将所得根系分层装入布袋,带回实验室。根系用清水洗净后,根据根系颜色和韧性去除死根保留活根,并装入信封中。将样品在 65 ℃的烘箱内烘干至恒重后称重(精确到0.01 g),然后将烘干后的根系样品磨碎,过0.25 mm筛子后装袋备用。植物根系全C、全N含量采用元素分析仪(vario Macro cube,德国Elementar公司)进行测定,全P含量采用钼锑抗比色法(硝酸-高氯酸前处理)测定。
1.3 数据分析
植物根系C、N和P密度由单位面积内根系生物量干重和其C、N、P元素含量相乘估算,其中每层根系C、N和 P密度由公式(1)计算,总根系C、N和P密度用公式(2)计算。用EXCEL对数据进行整理,并采用SPSS 17.0对数据进行分析,具体包括:同时期不同深度3种草地根系C、N和P密度的差异和同一深度不同草地类型间根系养分密度采用单因素方差分析(One-Way ANOVA),比较草地类型和土壤深度对非生长季C、N和P密度和生长季的 P密度非参数检验中的 Scheirer-Ray-Hare检验,而对生长季根系C和N密度的差异时用到了双因素方差分析(Univariate Analysis of Variance);比较非生长季和生长季根系C、N和P密度的差异采用独立样本T检验。运用 SigmaPlot 12.5进行图形绘制。

表1 3种草地信息Table 1 Information about three types of alpine grassland

式中D代表根系的C、N和P的密度,单位为g·m-2;i代表C、N和P中的某一种元素;j代表不同的土壤深度;B代表根系生物量,单位为g·m-2;ω代表根系 C、N和 P中某一种的质量分数,单位为 g·kg-1。

式中Dt为总根系C、N和P元素某种元素的密度。
2 结果与分析
2.1 根系C、N、P密度的垂直分布规律
除非生长季高寒草甸外,3种草地的根系C密度呈“T”型分布,即在两个时期均随土壤深度的增加显著降低(P<0.05,见表2和表3),最大值均出现在0—10 cm土层。在非生长季,高寒草原、草甸草原和高寒草甸根系分层 C密度最大值分别为(52.708±3.578)、(82.835±0.637)、(383.470±50.432)g·m-2;最小值分别为 (1.713±0.311)、(5.601±0.576)、(97.045±34.005) g·m-2。在生长季,高寒草原、草甸草原和高寒草甸根系分层 C密度的最大值分别为(79.976±6.945)、(49.428±2.892)、(716.135±97.648)g·m-2;最小值分别为 (1.481±0.223)、(0.513±0.078)、(178.373±22.775) g·m-2。在两个时期,草甸草原根系C密度最小值出现在20—30 cm根系,而高寒草原和高寒草甸10—20 cm与20—30 cm深度的根系C密度无显著性差异。

表2 生长季根系C、N、P密度的双因素方差分析Table 2 Analysis of two-factor variance of root C, N and P densities in non-growing season

表3 生长季根系C、N、P密度的双因素方差分析Table 3 Analysis of two-factor variance of root C, N and P densities in growing season
3种草地的根系N密度在两个时期均随土壤深度的增加显著降低(P<0.05,见表2和表3)。在非生长季,高寒草原、草甸草原和高寒草甸根系分层N 密度最大值分别为 (1.202±0.031)、(2.405±0.080)、(12.187±0.576) g·m-2;最小值分别为 (0.036±0.009)、(0.156±0.016)、(1.766±0.627) g·m-2。在生长季,高寒草原、草甸草原和高寒草甸根系分层N密度最大值分别为 (2.001±0.037)、(1.348±0.127)、(28.141±2.179) g·m-2;最小值分别为 (0.025±0.008)、(0.013±0.003)、(4.168±0.441) g·m-2。
3种草地的根系P密度在两个时期均随土壤深度的增加显著降低(P<0.05,见表2和表3)。在非生长季,高寒草原、草甸草原和高寒草甸根系分层P 密度最大值分别为 (0.132±0.006)、(0.236±0.045)、(1.144±0.175) g·m-2;最小值分别为 (0.004±0.001)、(0.019±0.001)、(0.236±0.074) g·m-2。在生长季,高寒草原、草甸草原和高寒草甸根系分层P密度最大值分别为 (0.137±0.021)、(0.079±0.007)、(2.230±0.123) g·m-2;最小值分别为 (0.003±0.001)、(0.001±0.000)、(0.391±0.065) g·m-2。
2.2 不同草地类型间植物根系C、N、P密度差异
在两个时期,不同草地类型间根系的C、N、P密度均有显著差异(P<0.05,见表2和表3)。在非生长季,3种草地同一深度根系的C、N、P密度均有显著差异(P<0.05,见表2)。所有深度的根系C密度均表现为:高寒草甸各层均最高,高寒草原最低,3种草地之间有显著性差异(P<0.05,表2)。0—10 cm、20—30 cm和总根系的N密度及总根系的P密度,高寒草甸各层均最高,高寒草原最低,3种草地之间有显著性差异(P<0.05)。10—20 cm的根系N密度、0—10 cm及20—30 cm的根系P密度,高寒草甸显著高于草甸草原和高寒草原(P<0.05),而高寒草原和草甸草原间无显著性差异(P>0.05)。
在生长季,高寒草甸各层根系C、N、P密度显著高于高寒草原和草甸草原(P<0.05,见表3)。0—10 cm和总根系的C、N密度,高寒草甸最高,草甸草原最低,且两两之间有显著性差异(P<0.05);其他的根系C、N、P密度均表现为:高寒草甸显著高于草甸草原和高寒草原(P<0.05),且草甸草原与高寒草原无显著性差异(P>0.05)。
2.3 根系C、N、P密度的季节变化
高寒草原生长季的总根系C、N密度均显著高于非生长季(P<0.05,见图1a、b),而总根系P密度无显著差异(P>0.05,见图1c)。生长季,高寒草原0—10 cm深度的根系C、N、P密度显著高于非生长季;而10—20 cm和20—30 cm的根系N、P密度显著低于非生长季(P<0.05,见图 1a—c);10—20 cm和20—30 cm的根系的C密度在两个时期无显著性差异(P>0.05,见图1a)。草甸草原总的和每层的根系C、N、P密度均表现为:生长季显著低于非生长季(P<0.05,见图1d—f)。高寒草甸生长季的总根系及0—10 cm的根系C、N、P密度均显著高于非生长季(P<0.05,见图1g—i),而10—20 cm和20—30 cm的根系C、N、P密度无显著的季节差异(P>0.05,见图 1g—i)。
3 讨论
3种草地的根系C、N、P密度在垂直空间上呈“T”型分布,与根系生物量的垂直分布规律相同(马星星等,2016),这一规律与Fan et al.(2008)的研究结果一致。高寒草原、草甸草原和高寒草甸 0—10 cm的根系C、N、P密度分别占总C、N、P密度的92.013%、82.255%和62.460%以上。这一结果表明:高寒草原根系 C、N、P密度在表层最为集中,高寒草甸最深,这与3种草地根系生物量分布特征完全一致。生长季,高寒草原、草甸草原和高寒草甸0—10 cm的根系C、N、P密度占比分别增加了2.175%、4.260%和0.872%以上,这表明3种草地的根系养分向表层转移,且草甸草原的养分表层转移幅度最大。这可能是因为生长季为植物的快速生长及繁殖阶段,植物将更多的生物量分配于地上,从而导致养分由根系向地上部分转移。
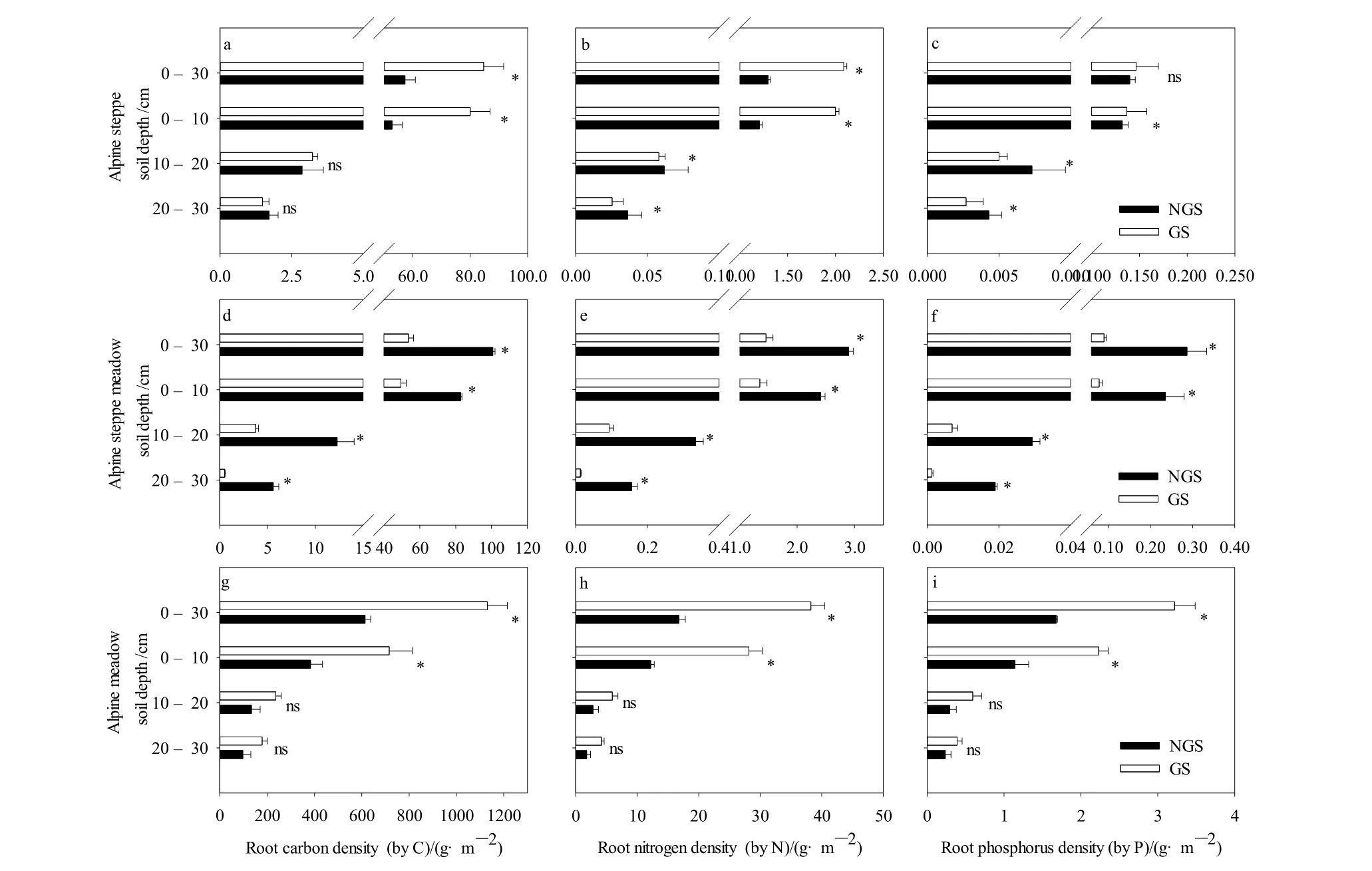
图1 高寒草原、草甸草原和高寒草甸根系C、N和P密度的季节差异Fig. 1 Root C, N and P densities of alpine steppe, alpine meadow steppe and alpine meadow in non-growing season and growing season
C是植物的结构性元素,C含量在植物体内的波动很小。氮磷元素受多种环境因素的影响和制约,温度、降水、N沉降和放牧均对植物C、N和P密度有一定的影响(白春利等,2016)。石福孙等(2008)对亚高山草甸的研究表明:增温提高了根系的碳储量,却降低了根系的氮储量。围封前期对根系C、N和P储量影响不明显,随围封年限增加,高寒草原根系C、N和P储量显著增加(洪江涛等,2015)。围栏显著增加了海拔4500 m处9.8%的植物群落地下部分N含量,显著增加了海拔4700 m处 4.2%的植物群落地下部分 C含量(沈振西等,2016)。在本研究中,3种草地的根系C、N、P密度,表现出显著的季节差异。高寒草原生长季的总根系C、N密度增加了47.822%和60.910%,而总根系P密度无显著差异。高寒草甸生长季的总根系C、N、P密度是非生长季的1.842倍以上。生长季,降水较集中,土壤水分条件较非生长季明显改善。有研究表明,根系N含量与干旱度指数呈负相关关系(Liu et al.,2019),且高寒草地根系碳密度呈现随降水增加而增加趋势(Fan et al.,2008),这可能是高寒草甸和高寒草原生长季根系养分密度增加的一个重要原因。草甸草原总的和每层的根系C、N、P密度在生长季显著降低,其中总根系C、N、P密度分别减少了46.687%、49.707%和68.641%。根据之前的研究,生长季草甸草原植物快速生长,地上部生长消耗大量根系部贮存的营养和能量,导致根系生物量持续减少(黄德青等,2011),这可能是根系C、N、P密度显著降低的一个重要原因。草甸草原土壤N含量高于其它两种草地类型,植物可能将更多的生物量分配到地上来获取其他的资源(Dawes et al.,2015)。
3种草地的根系C、N、P密度在草地类型间也有差异,且整体上表现为:高寒草甸显著高于其他两种草地。高添等的研究表明青藏高原草地生物量的平均C密度为223 g·m-2(高添等,2012),可见高 寒 草 原 (84.683±6.977) g·m-2和 草 甸 草 原(53.690±2.703) g·m-2的C密度低于青藏高原草地生物量的平均C密度,而高寒草甸 (1130.753±84.246)g·m-2根系C密度高于平均值。高寒草原的根系N、P密度与董晓玉等(2010)在黄土高原典型草原的研究结果基本一致。高寒草甸的根系N、P密度与Bardgett et al.(2007)和石福孙等(2008)对亚高山草甸的研究结果基本一致(Li et al.,2011),但远低于魏巍等在灌丛中的研究结果(N,50.510—60.970 g·m-2;P,5.37—7.38 g·m-2)(魏巍等,2012)。不同物种或者群落组成对土壤N、P养分的吸收和响应不同(Liu et al.,2019)。物种组成不同影响土壤的养分状况,反过来土壤的养分状况也影响植物中的N、P含量(Yan et al.,2018)。土壤中N含量与细根的N含量呈正相关关系(Liu et al.,2019),且土壤总P含量影响根系的N/P(Hong et al.,2015)。高寒草甸的优势物种为矮嵩草和大嵩草,生长周期长,其表层根系致密,分布深;再者高寒草甸的土壤水分和养分状况优于高寒草原和草甸草原,这些因素都可能造成高寒草甸根系养分密度量大。在下一步研究中,将探究土壤养分与植物根系养分含量间的关系。
4 结论
(1)3种高寒草地根系C、N、P密度呈现出“T”型垂直分布和明显的季节变化。3种草地0—10 cm的根系C、N、P密度显著高于其他深度的C、N、P密度。生长季高寒草原C、N密度和高寒草甸根系C、N、P密度显著增加,而草甸草原显著降低。由于3种草地所处的生长阶段和养分分配策略的差别,3种草地根系C、N、P密度的季节变化不同。
(2)高寒草甸的根系C、N、P密度显著高于其它两种草地。高寒草甸、高寒草原及高寒草甸草原的物种组成不同,土壤养分含量差异及土壤水分状况的不同可能是导致3种草地根系养分密度差异的原因。在以后的研究中,会将这些因素对根系养分密度的影响考虑进来。
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
海外文摘·文学版(2021年3期)2021-05-19
今日农业(2020年19期)2020-12-14
幼儿100(2020年31期)2020-11-18
疯狂英语·初中版(2019年4期)2019-09-10
活力(2019年21期)2019-04-01
小太阳画报(2018年6期)2018-05-14
中国科技信息(2015年2期)2015-11-16
植物营养与肥料学报(2014年1期)2014-03-11
