
含水量对超低温保存后马尾松种子生理生化特性的影响
2020-09-18 08:24张晓宁肖玉菲姚瑞玲陈博雯覃子海刘海龙
种子 2020年8期
张晓宁,张 烨,肖玉菲,姚瑞玲,陈博雯,覃子海,刘海龙
(1.广西壮族自治区林业科学研究院,南宁 530002;2.广西优良用材林资源培育重点实验室,南宁 530002)
超低温保存(cryopreservation)是指在不高于-100 ℃条件下保存生物材料,常用液氮(-196 ℃)作为冷源[1];在此温度下,能最大限度地抑制生理代谢活动,降低劣变频率。超低温保存是目前为止集简便、实用、低成本、占地少于一体的一种长期保存种质资源的方法[2],在农作物[3]、果蔬[4]、中药材[5]及林木种子[6-7]方面都有研究。
马尾松(PinusmassonianaLamb)从种子发芽到进入繁殖期时间间隔较长(实生植株约在第6年才进入正常的开花结实期)[8],而常温开放保存时间较短(3~4年几乎全部丧失发芽力),即便在2~4 ℃低温密闭条件下也只能维持10年左右[9-10]。因此,开展超低温保存研究对延长马尾松保存期限具有现实和必要意义,除了资源安全外也为其种质资源的交流、遗传改良和良种推广等生产实践和科学研究提供了可靠的物质保障[11]。
项目组前期对马尾松种子进行了超低温保存研究,结果表明,含水量是影响其冻存效果的关键影响因子,在含水量为6.1%时获得高达91.33%的冻后发芽率[12]。超低温保存不仅对种子生活力产生影响,还会对种子的生理生化特性产生影响[13],许多研究通过测量材料生理生化特性判断超低温保存效果[14,15]。
本研究在此基础上,对不同含水量马尾松种子冻存后REC、MDA、抗氧化物酶、非酶抗氧化剂、渗透调节物质的变化进行了研究,旨在从生理角度揭示不同含水量下各生理指标的变化规律,阐述其损伤/保护的生理机制,为进一步提高马尾松保存效果提供理论依据。
1 材料与方法
1.1 试验材料
成熟马尾松(PinusmassonianaL.)种子于2016年11月采自广西藤县,千粒重为11.4 g,发芽率为90.25%,初始含水量为9.56%。利用浸种和硅胶干燥的方式获取27.4%、22.7%、16.8%、10.7%、6.1%、4.8%、3.2%等7个含水量梯度的种子,不同含水量的种子冻存后发芽率见表1(见参文献[12])。设对照组和试验组,对照组直接测定生理指标,试验组经超低温保存后测定其生理指标。
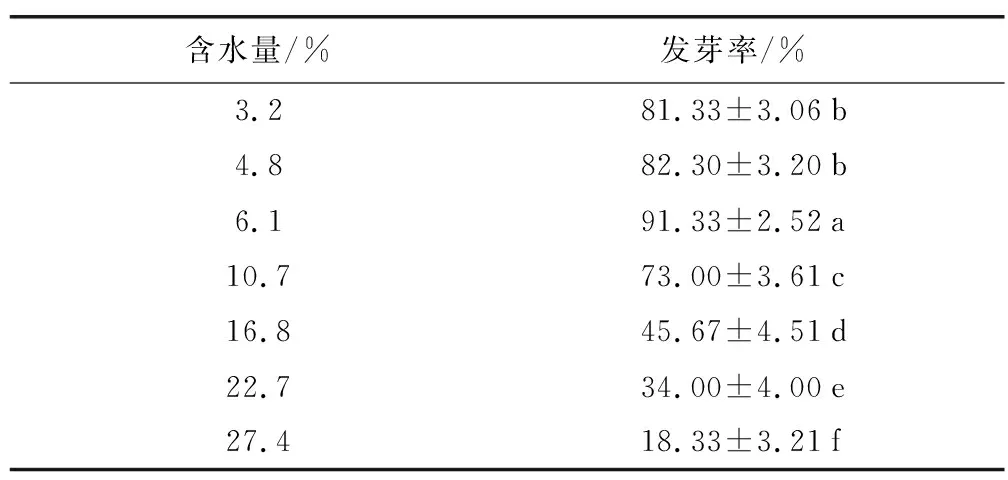
表1 不同含水量马尾松种子冻存后发芽率
1.2 测定方法
相对电导率(REC)的测定参见任淑娟等[16]的方法;丙二醛(MDA)质量摩尔浓度采用硫代巴比妥酸(TBA)法测定;超氧化物歧化酶(SOD) 活性采用黄嘌呤氧化酶法测定,过氧化物酶(POD)活性采用愈创木酚比色法测定,过氧化氢酶(CAT)活性采用磷钼酸法测定;AsA(Vc)含量采用菲咯啉比色法测定;可溶性糖、可溶性蛋白含量分别采用蒽酮法和考马斯亮蓝法测定。
1.3 数据分析
采用Microsoft Office Excel(2016)软件对试验数据进行整理作图,采用SPSS 19.0软件对实验数据进行方差分析、多重比较、生理生化指标的相关性及主成分分析。试验结果为3个重复的平均值±标准误。
2 结果与分析
2.1 MDA和种子浸出液REC的变化
如图1, REC和MDA含量受含水量影响显著,随含水量增加,变化趋势大体相同,均为先降后升,在含水量为6.1%时,REC和MDA含量最低,分别为15.33%和5.61 nmol·g-1,表明该含水量下质膜电解质外渗量少,脂质过氧化程度低,受到的损伤小。与MDA不同的是,REC表现出一个较广的适应范围,在含水量为4.8%~10.7%时,电导率值均较小,无显著差异。
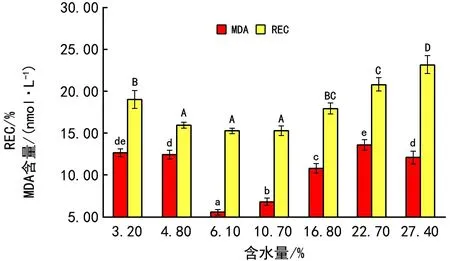
图1 含水量对马尾松种子冷冻保存后浸出液REC和MDA的影响
2.2 抗氧化物酶活性及非酶抗氧化剂含量的变化
不同含水量POD活性(图2)变化呈升—降—升—降的双峰型,在4.8%,6.1%和22.7%时最高,在10.7%时最低。CAT活性(图2)随含水量增加总体呈下降的趋势,在含水量为3.2%~10.7%的范围内活性较高,同样表现出一定程度的广适性。当含水量大于10.7%时,CAT 活性逐渐下降。SOD活性(图3)总体呈先升后降的趋势,在6.1%时达到最高值(577.92 U·g-1),后逐渐下降,直至含水量为16.8%时,达到最低值(414.31 U·g-1),含水量继续增加,SOD活性小幅上升。AsA含量变化与POD类似,呈双峰形状,在含水量为6.1%和22.7%时含量较高,低于4.8%时含量较低。

图2 含水量对马尾松种子冷冻保存后POD,CAT活性的影响

图3 超低温保存对不同含水量马尾松种子SOD活性,AsA含量的影响
2.3 可溶性糖(SS)和可溶性蛋白(SP)含量的变化
SS和SP含量变化如图4所示,SS在含水量为4.8%~10.7%时较高,在含水量小于4.8%或大于10.7%时显著降低。SP随含水量升高波动性变化,含水量为4.8%~6.1%时,达最大值。
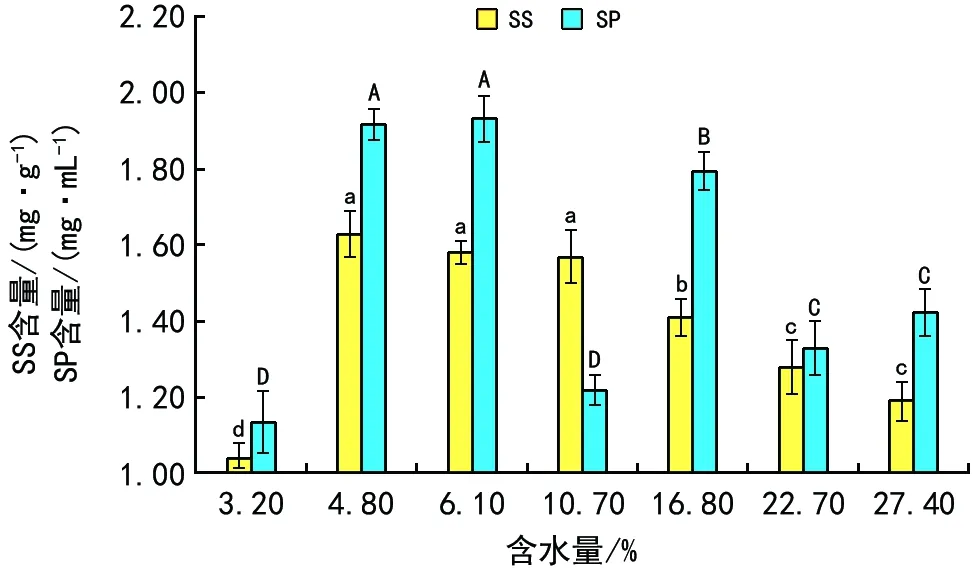
图4 超低温保存对不同含水量马尾松种子SS和SP含量的影响
3 结论与讨论
植物材料在超低温保存过程中,易受到代谢胁迫、脱水胁迫、结冰伤害及由此带来的机械损伤等,从而使其不同部位受到不同程度的损伤[17]。种子在低温逆境时,最先受损伤的是细胞膜系统[18]。脱水、冷冻和降温都可能会使细胞膜的脂质层发生相变(由液晶态转变为凝胶态),膜系统收缩,出现裂缝或通道,导致膜透性增加,细胞吸涨时,胞内电解质外渗,溶液电导率(REC)升高,因此,REC与膜受损伤的程度呈正相关[17]。本研究中,REC在含水量为6.1%时,达最低值,表明在此含水量下膜通透性较小,细胞膜系统受到的损伤较轻[19]。含水量增加或减少时,REC均增加,含水量太高容易导致冷冻和化冻阶段结冰使膜系统受到机械损伤,含水量太低在回复培养时会因快速复水产生吸涨伤害,太低的含水量也会降低种质活性,最终影响超低温保存效果[13,17]。
氧化伤害是超低温保存过程中的次生伤害之一,逆境胁迫(包括低温和脱水)会导致细胞内活性氧(ROS)的产生和积累,过量的ROS可使膜脂的不饱和脂肪酸发生过氧化作用,最终生成MDA[20,21], MDA对细胞有毒害作用,可降低SOD,POD等抗氧化物酶的活性,使蛋白质的结构和功能发生变化[22]; SOD,POD,CAT及AsA是重要的保护酶和非酶保护剂,可将过量有毒的ROS转化成无毒害的O2和H2O等物质。在含水量为6.1%时,MDA含量最低,各种抗氧化物酶活性和非酶抗氧化剂含量均达到最大。表明在此含水量下抗氧化能力较强,自由基水平较低,细胞膜系统受到的损伤较轻[14,19]。POD活性在4.8%,6.1%和22.7%时达到最大,一方面体现了其保护效应,高活力种子具有高的抗逆酶活性[23]。另一方面推测高含水量下POD参与ROS的产生,表现出对细胞的伤害效应[24]。POD的伤害效应在金丝李种子的研究中有报道,当种子含水量超过10.7%时,金丝李种子POD活性迅速增加[25]。CAT活性随含水量增加呈下降趋势,这与珙桐离体胚轴结果一致,表明高含水量下冷冻材料受到了一定程度的损伤[26]。
植物的超低温耐性受植物体内保护性物质影响。可溶性糖和可溶性蛋白作为重要的保护性物质,可通过增加细胞液浓度,降低细胞渗透势,提高细胞保水能力,降低细胞冰点,提高植物超低温耐性[17,27]。本研究中,在含水量为6.1%时,可溶性糖和可溶性蛋白的含量均达到最大。宋红等对玉婵花的研究结果也表明,适当降低含水量,可溶性蛋白质量分数上升,有利于保持种子的低温耐性,冻后发芽率显著升高[15]。
值得指出的是,在本研究中许多生理指标的最优值(统计学上无显著性差异)都是一个相对较宽的含水量范围,如REC值在含水量为4.8%~10.7%时较低;POD和CAT活性分别在含水量为4.8%~6.1%和3.2%~10.7%时较高;SS和SP含量分别在含水量为4.8%~10.7%和4.8%~6.1%时较高,这体现了正常型种子相对较广的安全含水量范围[12]。
本研究通过对生理指标的测量,进一步从生理层面证明6.1%左右的含水量为超低温保存的最适含水量,揭示了影响超低温保存效果的生理机制,即细胞膜透性小、膜脂过氧化程度低、细胞抗氧化能力强、胞内渗透调节物质浓度高,为进一步提高马尾松种子超低温保存效果,从分子层面解析超低温保存机理提供了理论基础。
猜你喜欢
农业灾害研究(2022年6期)2022-12-02
导弹与航天运载技术(2022年2期)2022-05-09
特种结构(2021年6期)2021-12-31
特种结构(2021年6期)2021-12-31
新农业(2021年12期)2021-11-29
南方农业·中旬(2021年5期)2021-10-25
上海农业科技(2020年2期)2020-04-02
山西农业科学(2020年1期)2020-02-27
绿色科技(2019年5期)2019-11-29
农民致富之友(2018年9期)2018-06-27
