
高寒地区钙果叶片表形特征差异分析
2020-09-17 13:44张巍刘运伟徐宜彬李阳任伟超
森林工程 2020年5期
关键词:遗传变异
张巍 刘运伟 徐宜彬 李阳 任伟超

摘 要:针对从吉林引进的钙果在小兴安岭伊春地区的栽培试验,通过分析叶片表形特征,对钙果在高寒地区的表形多样性表现进行探讨,为优选、改良寒地浆果类品种及开发利用提供技术支持。本文采用露地栽培和塑料大棚栽培两种设计,结合叶长、叶宽和叶面积等调查数据,对钙果叶片的比叶面积、遗传变异系数和叶片干物质含量等指标进行分析。结果表明,定植后的钙果叶片表形性状变异幅度为21.87%~34.44%,各品系均表现出较为丰富的遗传变异特征,这与环境因子的差异有直接关系。在不同立地条件下,不同品系表现出了不同的遗传程度,表现最为明显的是钙果4号品系。结论:在引进伊春地区后,钙果不同品系间遗传丰富度差异确实存在;各品系表形特征产生的差异性均取决于环境因子;而各品系遗传活跃度相近也证明了钙果品系内遗传性状相对稳定。
关键词:钙果;叶片变异;表形差异;遗传变异
中图分类号:S662 文献标识码:A 文章编号:1006-8023(2020)05-0040-05
Abstract:In this paper, Cerasus humilis introduced from Jilin Province was cultivated in Yichun area of Xiaoxingan Mountains. By analyzing leaf surface characteristics, the phenotypic diversity of Cerasus humilis in alpine regions was discussed, which provided technical support for the optimization and improvement of berry varieties in cold region and their development and utilization. The experimental sample plots were established by two designs of land cultivation and plastic greenhouse cultivation respectively. The leaf area, genetic variation coefficient and dry matter content of the leaves were analyzed by combining with the data of leaf length, leaf width and leaf area. Results showed that: the variation range of phenotypic traits of leaves ranged from 21.87% to 34.44%, and all strains showed abundant genetic variation characteristics, which was directly related to the differences of environmental factors. Different strains showed different degree of genetic activity under different site conditions, and the most obvious one was No. 4. After the introduction into Yichun area, the differences of genetic richness between different strains of Cerasus humilis did exist and the differences in the phenotypic characteristics were all from environmental factors. The similarity of genetic activity among the strains also proved that the genetic characters in the Cerasus humilis strains were relatively stable.
Keywords:Cerasus humilis; leaf variation; phenotypic difference; genetic variation
0 引言
植物的表型特征是其本身各種形态特征的组合,是植物自身遗传因素和环境因子共同作用过程中,经过长期适应,群体和个体间均产生不同程度形式的变异,以适应不同的环境[1],而植物叶面积的大小及其分布, 直接影响着植物对光能的截获及利用 ,进而影响种群生产力。叶片性状对气候变化非常敏感, 反映了植物对环境的高度适应能力和在复杂生境下的自我调控能力[2], 故以植物叶片为研究对象, 更能反映植物对气候变化的响应与适应机制。如曹红梅等[3]对蓝莓叶面积进行了测定并建立了相关回归方程。陈丽君等[4]分析了苦楝10个种源的叶片性状变异情况,表明各种源间叶片性状变异明显。
钙果(Cerasus humilis)为蔷薇科樱桃属多年生落叶小灌木,是从我国野生欧李中选育出的一种新型果树。钙果具有多种价值,果实富含钙和铁元素,其含量分别为0.60 mg/g和0.15 mg/g,是苹果的6.7倍和6.3倍。目前对于钙果的研究主要以引种适应性、有效物质提取和精深加工等为主,对于钙果异地引种后如何通过叶片水平上的表型变异测定群体内或群体间水平的遗传差异方面研究较少,同时这也是钙果生态模拟研究领域中的一个理论空白。
课题组于2018年引进钙果3个品系在小兴安岭伊春林区统一建立试验样地进行适生性试验。试验设计塑料大棚栽培和田间露地栽培两种栽培方式,通过对定植后钙果叶片的遗传表现进行分析,试图探讨钙果在高寒地区定植后的表形遗传变异表现,并对钙果在高寒地区不同立地条件下叶片功能性状是否产生差异及对植株生殖生长的影响进行观测,为今后钙果在高寒地区的引种及育种工作做好前期工作。
1 材料与方法
1.1 试验地概况
试验样地位于黑龙江省友好林业局万亩蓝莓园内,总面积1.3 hm2。试验地平均海拔347.1 m,年积温2 000~2 300 ℃,日照时数2 430.4 h,无霜期117 d,年降水量629.6 mm。年平均气温为0.4 ℃,1月份平均气温-23 ℃,7月份平均气温为20.5 ℃,冬季平均气温-16.2 ℃,平均年降水量为630 mm,早霜9月上旬,晚霜翌年5月中下旬消失,无霜期110 d,属北温带大陆性湿润季风气候区,土壤为暗棕壤。样地内土壤pH为5.51,有机质含量为11.23%,全效氮含量33.67 mg/kg,全效磷含量201.70 mg/kg,全效钾含量193.78 mg/kg。
课题组于2018年分别引进钙果4号、6号和7号3个品系。当年在友好林业局万亩蓝莓园内统一建立试验样地。样地共分棚式栽培和露地栽培两种设计。其中,棚式栽培采用连体大棚栽培,连体大棚总面积1.3 hm2,内建立试验样地一处,面积0.67 hm2。为防止边缘效应,各品系采取不同定植设计,两次重复。露地栽培试验样地1处,面积0.27 hm2,栽培品系为钙果4号和6号两个品系,两次重复。两种栽培设计造林密度均为1 m×1 m。
1.2 研究方法
叶片采集于2019年8月进行,共分为塑料大棚栽培和露地栽培两个设计。大棚内以钙果4号、6号和7号3个品系为指标进行采集。露地栽培以4号和6号为主进行采集。因叶片不同位置变异较大,所以在采集叶片时,以每单株的上、中、下部分各取5片叶片进行测量,采集枝段上已完全展开的叶片,未完全展开叶片、发黄衰老和不完整叶片不采集。依据不同品系分开保存在自封袋中,袋面上做编号,并单独记录编号带回实验室。
将新采集叶片洗净后,依据各部位叶片进行分别测量。测量指标主要有叶鲜重、叶长和叶宽。计算叶面积、长宽比和叶形系数等。测量后将叶片统一编号装入密封袋带回,当日不能进行测量的样品放入冰箱冷冻保存。叶片洗净后,置于80 ℃干燥箱中烘干48 h至恒重,称量样品干重[6]。运用公式(1)—公式(3)进行变异系数、比叶面积(specific leaf area,SLA,面积与干重的比例在公式中用S(SLA)表示)和叶片干物质含量(leaf dry matter content,LDMC,干重与鲜重的比率在公式中用l(LDMC)表示)的计算[7],公式如下:
l(LDMC)=叶干重/叶湿重。 (1)
S(SLA)=叶面积/叶干重。 (2)
CV=(SD/MN)×100%。 (3)
式中:CV为变异系数;SD為标准偏差;MN为平均值;S(SLA)为比叶面积;l(LDMC)为叶片干物质含量。
叶面积测量用杭州大吉光电仪器有限公司的YMJ-A叶面积仪进行测量。数据分析用SPSS19.0完成,其他数据用WPS-Excel 2019进行相关分析。
2 结果与分析
2.1 不同品系叶片分析
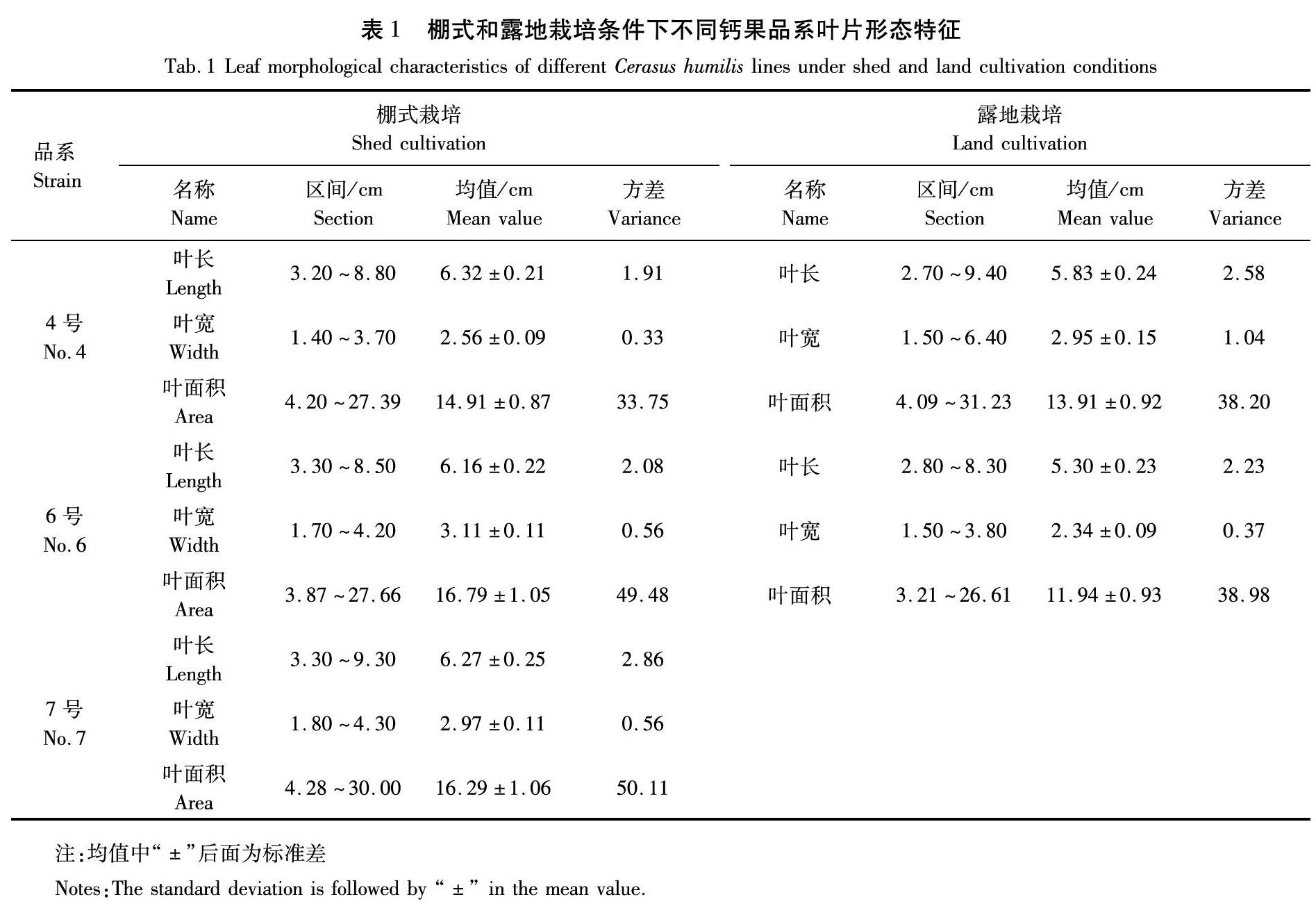
叶片受控于植物遗传特性,其表形是植物遗传变异和环境差异的共同反映。而叶面积指数(LAI)作为一个重要生理指标,在气体交换、光合产量和水分利用等方面都是必不可少的参数[8]。综合分析可知,棚式定植的钙果叶片长度总体平均值为6.25 cm,宽度总体平均值为2.88 cm,叶面积总体平均系数为16.00 cm2。露地定植钙果叶片长度总体平均值为5.56 cm,宽度总体平均值为2.64 cm,叶面积总体平均系数为12.92 cm2。表明棚式栽培钙果叶片交错程度明显高于露地栽培苗木。
分别对钙果4号、6号及7号进行叶面积分析,结果表明,钙果4号叶片长度取值范围为3.20~8.80 cm,叶片宽度取值范围为1.40~3.70 cm。叶面积总体平均值为14.91 cm2,稍小于其他两个品系。而6号和7号两个品系叶面积指数相近,分别为16.79 cm2和16.29 cm2。相比之下,露地栽培钙果4号和6号品系叶面积分别小于棚式栽培钙果叶面积7.19%和40.62%(表1)。
对数据进行不同品系叶片系数T检验结果表明,叶片指数在置信度为95%时,6号和7号品系组间概率P值为0.502,大于0.05,证明组间叶片指数无明显差异。同样,钙果4号与钙果7号两个品系组间概率P值为0.072,大于0.05,表明组间差异不明显,但钙果4号和6号两个品系组间概率P值为0.031,表明两个品系间叶片变异系数存在显著差异。
综上分析表明,棚式栽培的钙果叶片明显大于露地栽培的钙果叶片。这是由于相对于露地栽培,塑料大棚栽培方式不但可以为钙果在生长过程中提供更为有利的自然因子条件,且通过温、湿度及光照的调控,对钙果的生殖生长产生了积极作用,从而也证明,钙果本身对外部环境刺激较为敏感。
2.2 叶片变异性状关联性分析
对各品系数据进行统计分析,由表2可知,在对棚式栽培各品系叶片鲜重、干重统计中,6号品系统计平均值大于其他两个品系。而在比叶面积(SLA)和干物质含量(LDMC)两个指标计算中,7号品相关系数均高于其他两个品种。露地栽培统计中,4号品系鲜重、干重两项指标系数相对高于6号品系。变异系数方面棚式栽培7号品系和露地栽培4号品系的遗传系数较高,表明不同品系在环境因子作用下表现有所差异。对各样本进行T检验结果表明,叶片指数在置信度为95%时各品系系数均小于0.05,表明各品系组内高度相关,不存在明显差异。
比叶面积(SLA)为叶面积与其干重之比。在同一个体或群落内,一般受光越弱比叶面积越大。分别对钙果4号和6号进行棚式栽培和露地栽培的比叶面积比较。表2显示4号和6号钙果品系其棚式栽培比叶面积均稍大于露地栽培苗木。对两个品系的两种栽培方式分别进行相关性分析,结果表明,棚式栽培4号、6号钙果和露地栽培4号、6号钙果相关系数分别为0.57和0.65。在置信度为95%前提下,钙果6号品系检测值为0.00,表明虽然两组叶面积有所差异,但总体相对稳定,并没有受到环境因子改变的影响。而钙果4号品系棚式栽培区组和露地栽培区组检测值为0.23(P>0.05),表明4号品系苗木叶片因不同环境因子刺激而产生一定差异。这也表明相对于其他品系,4号品系遗传变异更为丰富。干物质含量(LDMC)是判断植物光合作用及分解作用的重要指标。通过钙果对干物质量进行计算证明,各品系叶片干物质含量主要分布于0.48~0.55区间,而相同品系在不同生境定植后,其叶片干物质量基本处于同一区间,并无明显差异。
分别对6个钙果区组进行叶片的遗传变异分析,结果表明种群的表形性状变异幅度为21.87~34.4,平均变异系数为 26.01%。以叶片长度和宽度为计算单位进行遗传系数计算,结果表明,不同种组间同一表形性状的变异范围不同,同一组内的表形变异也因不同性状而有差别。其中露地栽培钙果4号品系叶片宽度指数遗传变异系数为34.44,是各组最大的。叶片长度遗传变异指数为27.53,与其他区组遗传系数差异不大。由此表明露地栽培的钙果4号品系叶片遗传差异主要是由于其叶片宽度的变异,而叶片长度的变异与其他区组差异不大。
3 讨论
植物表形的遗传变异程度是植物适应不同环境因子的外在表现[8-11]。而叶片是植物制造光合产物的重要器官,是影响植物生产力大小的决定性器官之一[12],对植物的生殖生长具有重要意义。目前一般将变异系数≥30 的规定为高度变异,在15~30 的为中度变异,<15 的为低度变异。通过对3个钙果品系及两种不同栽植方式的对比分析表明,3个品系组内及组间平均变异系数小于 20,属中度变异。从而表明钙果在被引进后,因环境因子发生巨大改变使各品系总体发生一定程度的表形变异情况,这也证明钙果这一品种表形多样性较为丰富。
对组内和组间的测定表明,各组组内遗传变异系数总体处在同一合理区间,证明虽然因地理环境改变而产生一定表形变异,但总体性状变异均处于同一区间,相对稳定,并没有表现出因环境因子改变而产生组内分化现象。叶片性状遗传大致可分为数量性状的遗传和质量性状的遗传。其区别为数量性状的遗传主要由多基因控制,加上外部环境等因素影响,可能会发生不同性状间的变异系数有较大差异的现象。此次钙果引进在不同环境定植后,组间的性状差异是存在的,其中组间差异表现最为明显的是钙果4号品系,这也与周文才等[13]的观测结果相近。
钙果总体遗传稳定性较强,环境因子在一定范围内的波动不会对群落产生较大影响。但在环境因子波动较大时,钙果叶片的表形性状会发生变异,而这种表形變异的产生与环境因子呈正相关关系。这表明钙果叶片在寿命期内的表现取决于其生境因子和其本身适应性的双重生理制约机制,叶片所表现出来的表形变异与环境因子的影响之间存在权衡关系,这也与王常顺等[14]的研究结果相近。表形变异是由遗传因子和环境因素共同作用的结果,表形变异必然蕴藏遗传变异,一般来说表形变异越大,遗传变异也就越大。本文以不同品系钙果叶片表形性状进行分析表明其表形性状自然变异丰富,这同时也表明钙果具有广阔的遗传改良前景。
【参 考 文 献】
[1]卢欣石,何琪.种群遗传变异及基因多样度分析[J].草业学报,1999,8(3):76-82
LU X S, HE Q. The genetic variation of population and the analysis for gene diversity[J]. Acta Prataculturae Sinica, 1999, 8(3): 76-82.
[2]张深梅,奚建伟,洪俊彦,等.大别山山核桃果实与叶片性状的表型多样性研究[J].林业科学研究,2020,33(1):152-161.
ZHANG S M, XI J W, HONG J Y, et al. Phenotypic diversity of fruit and leaf traits in Dabie Mountain[J]. Forest Research, 2020, 33(1):152-161.
[3]曹红梅,宋世鑫,谢兆森.蓝莓叶面积测定回归方程的建立[J].北方园艺,2014(24):1-6
CAO H M, SONG S X, XIE Z S. Establishment of regression equation for blueberry leaf area determination[J]. Northern Horticulture, 2014(24):1-6.
[4]陈丽君,刘明骞,廖柏勇,等.苦楝不同种源叶片性状变异研究[J].西南林业大学学报,2016,36(5):16-20.
CHEN L J, LIU M H, LIAO B Y, et al. Study on the variation of leaf characters of different provenances of Melia azedarach[J]. Journal of Southwest Forestry University, 2016, 36(5):16-20.
[5]張巍, 李金禹,李妍,等.宽叶蓝靛果叶片不同海拔和郁闭度的遗传变异研究[J].森林工程,2018,34(3):14-18.
ZHANG W, LI J Y, LI Y, et al. Study on genetic variation in different altitude and crown density of the wide-leaf Lonicera turczaninowii leaves[J]. Forest Engineering, 2018, 34(3): 14-18.
[6]黄海侠,杨晓东,孙宝伟,等.浙江天童常绿植物当年生与往年生叶片性状的变异与关联[J].植物生态学报,2013,37(10):912-921.
HUANG H X, YANG X D, SUN B W, et al. Variability and association of leaf traits between current-year and former-year leaves in evergreen trees in Tiantong, Zhejiang, China[J]. Chinese Journal of Plant Ecology, 2013, 37(10): 912-921.
[7] 高祥斌,张秀省,蔡连捷.观赏植物叶面积测定及相关分析[J].福建林业科技,2009,36(2):231-234.
GAO X B, ZHANG X S, CAI L J. The determination and correlation analysis of leaf area of ornamental plant[J]. Journal of Fujian Forestry Science and Technology, 2009, 36(2): 231-234.
[8]陈丽君,刘明骞,廖柏勇,等.苦楝不同种源叶片性状变异研究[J].西南林业大学学报,2016,36(5):16-20.
CHEN L J, LIU M Q, LIAO B Y, et al. Variation of leaf morphological characters of Melia azedarach from different provenances[J]. Journal of Southwest Forestry University, 2016, 36(5): 16-20.
[9]汪娟,张华,张杰,等.紫叶李叶片春季成熟过程中花青素含量和光合功能变化研究[J].林业科技,2018,43(3):13-17.
WANG J, ZHANG H, ZHANG J, et al. The changes of anthocyanidin content and photosynthetic function of Prunus cerasifera leaves during the development process in spring[J]. Forestry Science & Technology, 2018, 43(3):13-17.
[10]胡玉玲,姚小华,张山,等.油茶叶片生理生化指标动态变化[J].林业科技,2018,43(1):17-21.
HU Y L, YAO X H, ZHANG S, et al. Changes of physiological and biochemical indexes of oil-tea Camellia oleifera leave[J]. Forestry Science & Technology, 2018, 43(1):17-21.
[11]罗传文.均匀论[M].北京:科学出版社,2014.
LUO C W. Uniform Theory[M]. Beijing: Science Press, 2014.
[12]李守勇,赵永军,孙明高,等.11个黑杨无性系叶片性状变异研究[J].山东林业科技,2002,32(2):5-7.
LI S Y, ZHAO Y J, SUN M G, et al. Study on lamina character of 11 clone of Populus nigra[J]. Journal of Shandong Forestry Science and Technology, 2002, 32(2): 5-7.
[13]周文才,巢军,孙颖,等.不同钙果品系引种试验[J].南方林业科学,2017,45(1):14-16.
ZHOU W C, CHAO J, SUN Y, et al. Introduction experiment of different Cerasus humilis[J]. South China Forestry Science, 2017, 45(1): 14-16.
[14]王常顺,汪诗平.植物叶片性状对气候变化的响应研究进展[J].植物生态学报2015,39(2):206-216.
WANG C S, WANG S P. Research progress in response of plant leaf characters to climate change[J]. Chinese Journal of Plant Ecology, 2015, 39 (2): 206-216.
猜你喜欢
湖北农业科学(2017年22期)2017-12-25
中国医药导报(2017年30期)2017-11-16
成长·读写月刊(2017年10期)2017-10-15
江苏农业科学(2016年1期)2017-05-17
魅力中国(2016年19期)2017-04-18
电子技术与软件工程(2017年1期)2017-03-06
吉林农业(2016年12期)2017-01-06
中学生物学(2016年7期)2016-11-02
江苏农业科学(2014年10期)2014-11-22
江苏农业科学(2014年9期)2014-11-15
