
极小种群野生植物浙江安息香的生态位和种间关系
2020-09-17 01:02:42吴霖东李婷婷傅国林吴初平余裕龙王志高盛卫星朱锦茹江波焦洁洁文香英袁位高
浙江大学学报(农业与生命科学版) 2020年4期
吴霖东,李婷婷,傅国林,吴初平,余裕龙,王志高,盛卫星,朱锦茹,江波,焦洁洁,文香英,袁位高*
(1.浙江农林大学林业与生物技术学院,杭州311300;2.浙江省林业科学研究院,杭州310023;3.浙江省建德市建德林场,杭州311600;4.浙江省建德市林业技术推广中心,杭州311600;5.中国科学院华南植物园,广州510650)
浙江安息香(Styrax zhejiangensis)为安息香科安息香属的一个种,是浙江省特有的极小种群野生植物,分布区域极为狭窄,目前仅发现于浙江省建德林场泷江林区桃花坞;其数量稀少,濒临灭绝,2011年调查发现仅有37 丛,属极其濒危,已被列为浙江省珍稀濒危野生植物及省级重点监测物种。浙江安息香树形优美,花朵芳香且盛开时下垂美丽,是珍稀的观赏植物,其花、果实、木材均具有重要的经济价值,可用于园林绿化、药用保健以及能源利用等方面[1]。目前,国内外对于安息香属植物保护方面的研究大多处于资源现状调查阶段,特别是对于一些国内特有种的濒危原因等方面的研究较为缺乏,严重影响了该属植物资源的保存和利用[2-3]。
极小种群野生植物因其分布地域狭窄或呈间断分布并长期受到外界因素胁迫干扰,导致种群退化和数量持续减少,目前,种群及个体数量都极少,已经低于稳定存活界限的最小可存活种群而随时处于濒临灭绝状态[4-5]。研究极小种群的生态位特征和种间联结性能够量化极小种群植物与环境之间的关系以及其在群落中所处的地位,并反映群落中物种之间的竞争关系[6]。生态位理论在植物种群研究中有重要而广泛的应用,通过对植物种群之间生态位重叠及生态位宽度的计算,可以定量地反映物种之间的关系以及物种和环境之间的关系[7-9]。极小种群植物在种群竞争间会处在劣势地位,比如极小种群植物玉龙蕨(Sorolepidium glaciale)在种群间竞争中处于劣势,数量稀少,对资源利用率低,不能形成优势种群[10]。但有些时候造成极小种群植物濒危的主要原因并不是生态位因素,比如极小种群野生植物圆叶玉兰(Oyama sinensis)[11]和重庆特有极小种群植物缙云秋海棠(Begonia jinyunensis)[12]都具有较强的生态适应能力,在群落中都具有重要的生态地位和作用。另外,对群落种间联结性进行研究,能反映不同物种之间的相互作用及物种与生境因子间的耦合关系,为物种多样性的保护提供种间关系依据[13]。胡理乐等[14]对濒危植物毛柄小勾儿茶(Berchemiella wilsonii var. pubipetiolata)和黄祥童等[15]对珍稀植物对开蕨(Phyllitis scolopendrium)与其伴生物种联结性进行研究,提供了相应物种保护和回归实践的理论依据。
本文研究了浙江安息香野生植物群落的物种组成、种群密度、生态位宽度、生态位重叠和种间联结性等,即从生态位特征和种间关联方向分析其是否存在生态适应能力弱以及与周围树种发生资源竞争等可能存在的濒危原因,旨在为浙江安息香的就地保护与迁地保护提供理论参考,并为该物种的科学保护积累资料。
1 材料与方法
1.1 研究区概况
浙江安息香种群主要分布在浙江省建德林场,该林场拥有经营面积8 000 多hm2,森林蓄积量达65 万多m3,森林覆盖率95%以上,是国家级森林公园,地处钱塘江上游的富春江水库两岸。浙江安息香种群保护区所在的建德林场泷江林区桃花坞,地形复杂,植被资源丰富,境内分布的木本植物有642种,隶属91 科、276 属,其中许多是国家珍稀保护植物,非常适合浙江安息香种群保护区的建设。
1.2 研究方法
参考方精云等[16]的方法,采用样地记录法进行群落调查。由于目前发现的浙江安息香共有8个分布点,因此,根据这8个分布点的位置分为8个调查样地,即在每个分布点设置1个样地,其位置分布如图1 所示。样地面积设置为20 m×20 m,观测记录每个样地海拔、坡向、坡度、坡位、郁闭度等,获得生境原貌;对样地中的所有浙江安息香予以调查,记录树高、基径、冠幅、枝下高等;将每个样地分为4个10 m×10 m 的小样方并调查乔木层的树木,记录种名、株数、胸径、树高、冠幅等;同时,在每个样方沿对角线方向设置5个5 m×5 m的小样方并测量灌木层和草本层的种名、盖度、高度,获得群落的基本信息。样地的环境特征如表1所示。
1.3 数据处理
1.3.1 种群密度
Nd=S/A.
式中:Nd为种群密度;S为样地内浙江安息香的总株数;A为样地面积[17]。
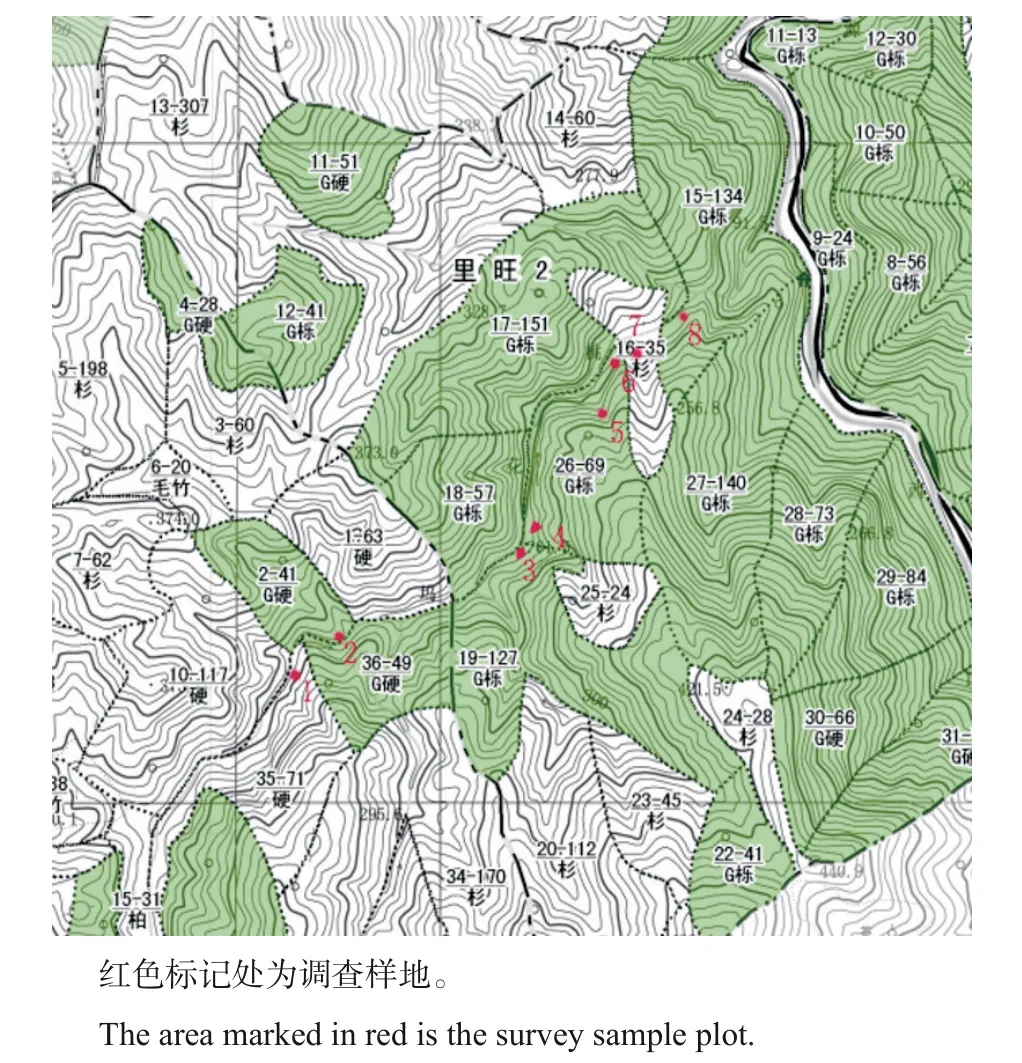
图1 浙江安息香样地分布图Fig.1 Sample plot distribution of Styrax zhejiangensis
1.3.2 重要值
重要值能反映各个种群在群落中的地位和作用[18],浙江安息香群落中各树种重要值的计算公式如下:

表1 样地的环境特征Table 1 Environmental characteristics of sample plots
相对密度=某一物种在样方中的株数/该样方中所有物种的总株数;
相对优势度=某一物种在样方中的胸高断面积和/该样方所有树种的胸径平方和;
灌木和草本相对优势度=某一物种在样方中的盖度/样方中所有物种的盖度和;
相对频率=某一物种在样方中出现的频率/该样方中所有树种出现的频率和;
重要值=(相对密度+相对优势度+相对频率)/3×100%。
1.3.3 生态位宽度
生态位宽度可以反映物种对环境资源利用状况以及物种的生态适应性和分布幅度[19]。本文分别采用Levins和Shannon法的生态位宽度计算公式[20-21]。

式中:BL和BS分别为Levins和Shannon法中物种i的生态位宽度;Pij为物种i在资源位j中的重要值占该物种在整个资源中总重要值的比例;r 为资源等级数(本文为样地数);nij为物种i在资源位j上的优势度(本文为样方中物种的重要值);Ni为物种i的总量(本文为物种i在所有样地的重要值的总和)[12]。
1.3.4 生态位重叠
生态位重叠是指在一定资源序列上,2 个物种利用同等级资源而互相重叠的情况[19]。本文采用Pianka指数来表示生态位重叠,其计算公式[22]为:

式中:Oik为生态位重叠值;pij和pkj分别为物种i与物种k 在资源位j 中的重要值占该物种在整个资源中总重要值的比例;nij和nkj分别为物种i和物种k在资源位j 上的优势度(本文为样方中物种的重要值)。Oik具有值域[0,1]的特点:当Oik为1 时,表明物种i与物种k的生态位完全重叠;当Oik为0时,表明2个物种没有共同资源状态,其生态位完全不重叠[23]。
1.3.5 种间联结
以χ2检验为基础,建立2×2联列表,利用结合点相关系数(point correlation coefficient, PCC)、联结系数(association coefficient, AC)、关联度指数(Ochiai index, OI)、共同出现百分率(percentage of co-occurrence,PC)进行种间联结分析[24-25]。
1)χ2检验可以确定物种间联结性是否显著。由于本研究取样的非连续性,χ2值采用Yates的连续校正公式进行计算[15,26]。
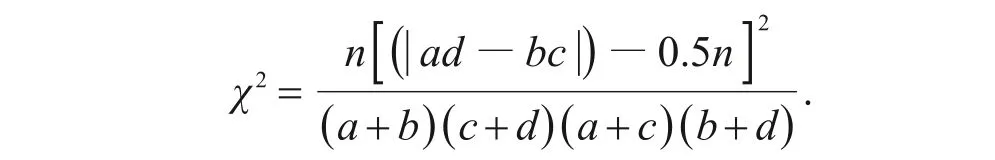
当ad-bc>0时,种对间呈正联结关系,反之则为负联结。种对间联结性强弱由χ2值决定:若χ2<3.841(P>0.05),则种对间联结不显著;若3.841<χ2<6.635(0.01<P<0.05),则种对间联结显著;若χ2>6.635(P<0.01),则种对间联结极显著。
2)点相关系数(PCC)计算公式为:
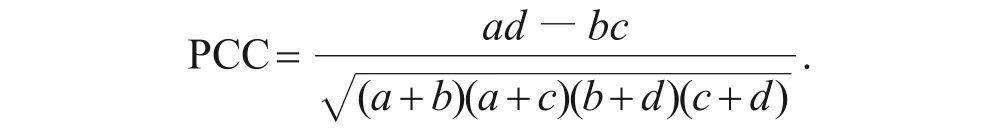
3)联结系数(AC)表示种对间的联结程度。计算公式为:

式中:n 为样方总数;a 为A、B 2 个物种都存在的样方数;b为A物种不存在而B物种存在的样方数;c为A物种存在而B物种不存在的样方数;d为A、B 2个物种都不存在的样方数[18,25]。
在计算联结系数(AC)时,为避免a、b 或d 为0而导致AC=±1,即出现无法比较关联程度的情况时,将a、b、c、d为0的数值都加权为1[15,25]。
2 结果
2.1 群落物种组成
浙江安息香主要生长于生境相对湿润的溪流边。根据8 个样地调查结果,浙江安息香群落内有约91 种植物(表2~4)。其中乔木层和灌木层的植物约65种,隶属于33科49属,总数达2 422株,包括安息香科、冬青科、壳斗科、山茶科、樟科、榆科等。在乔木层和灌木层中,植物数量超过20株的物种占其总物种数的24.62%,其林木数量占其植物总数量的86.94%,其中:箬竹(Indocalamus tessellatus)数量最多,约1 181 株,占植物总数量的48.66%;而浙江安息香仅有114株(66丛),占植物总数量的4.71%;另外有35 个物种的林木数量不超过10 株,占总物种数的53.85%,占植物总数量的5.08%。
种群密度是反映种群自我数量调节的作用系统指标,也是影响种群数量动态的主要因子[17]。从表2~4中各物种种群密度可知:箬竹的总体种群密度最大,为3 691株/hm2,其次为青冈(Quercus glauca)、油茶(Camellia oleifer)、浙江安息香、柃木(Eurya japonica)等。可见,浙江安息香的种群密度排在第4位,为356株/hm2。
2.2 重要值特征
由表2~4 可以看出:乔木层中,青冈的重要值最大,其次为杉木(Cunninghamia lanceolata)、木荷(Schima superba)、蓝果树(Nyssa sinensis)、红楠(Machilus thunbergii)等。灌木层中,箬竹的重要值最大,为26.45%;浙江安息香的重要值为10.74%,位居第3,是所在群落灌木层中的优势种群之一;重要值前5的物种还包括油茶、柃木、青冈。在实际调查中,占据灌木层第1位的箬竹在各个样地分布均匀,种群数量也多。与箬竹相比,浙江安息香的优势并不明显,种群数量也相对较少。
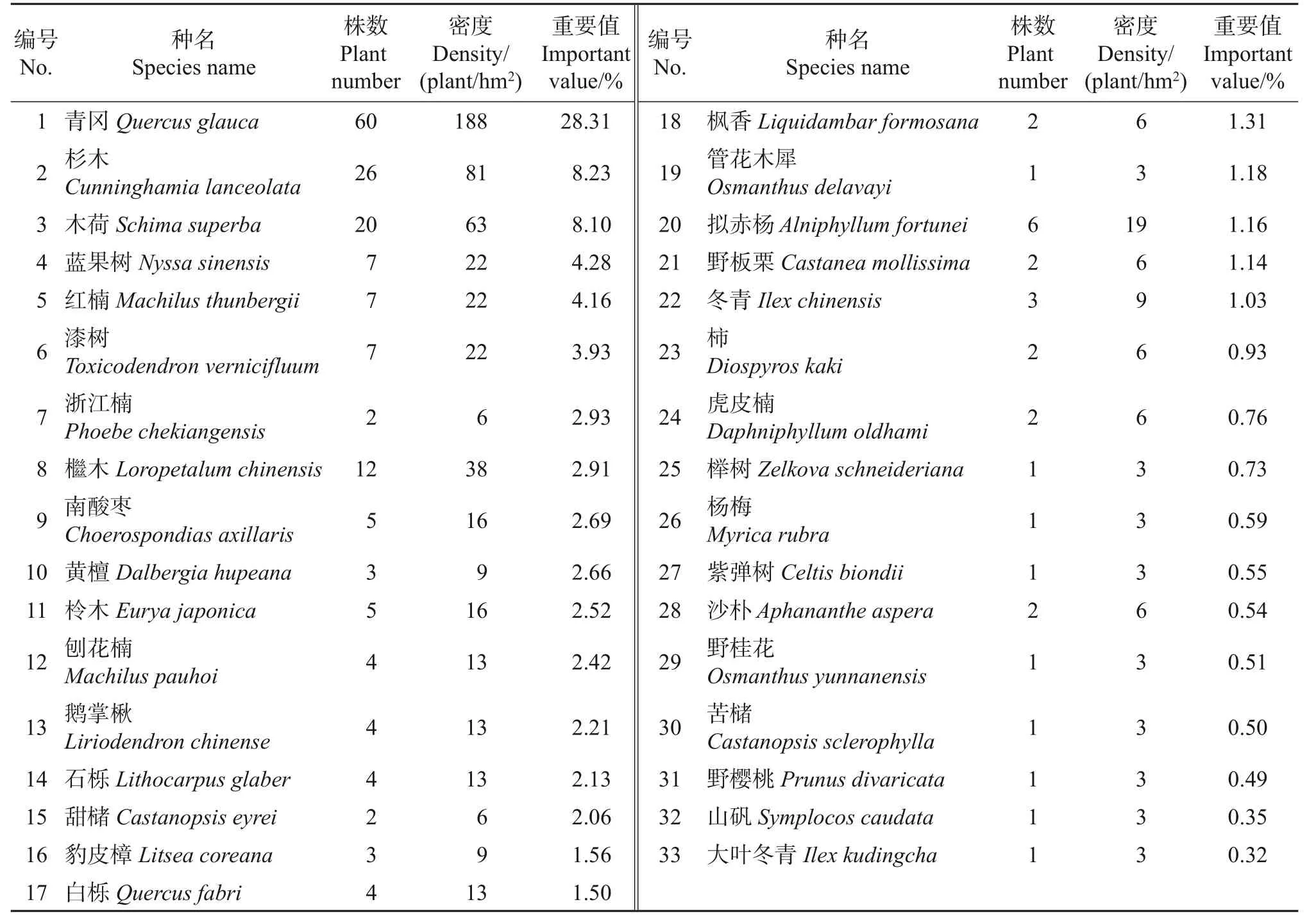
表2 乔木层物种组成Table 2 Species composition of tree layer

表3 灌木层物种组成Table 3 Species composition of shrub layer
2.3 生态位宽度特征
生态位宽度可以衡量植物种群对环境的适应能力和资源利用能力。种群生态位宽度越大,则表明该种群对环境的适应能力强,对资源的利用能力也强[12]。本文分别采用Levins 和Shannon 法计算浙江安息香群落中木本植物的生态位宽度,二者的计算结果与物种重要值的变化趋势基本一致(表5)。总体上,物种的重要值越大,生态位宽度也越大。
在乔木层,青冈的重要值最大,其生态位宽度也最大,分别为6.126和1.904,表明其生态适应能力相对其他树种强,且在实际调查过程中,青冈在各个样地均有出现,其竞争能力较强。而杉木、蓝果树和南酸枣(Choerospondias axillaris)等树种的重要值排序虽然较其他树种相对靠前,但其只出现在个别样地中,使其生态位宽度相对靠后。相反,红楠、柃木、刨花楠(Machilus pauhoi)和虎皮楠(Daphniphyllum oldhami)等树种的重要值虽然相对靠后,但其出现的样地数量较多,频度较大,使其生态位宽度相对靠前。
在灌木层,虽然浙江安息香重要值排第3,但其生态位宽度最大,分别为6.557和1.977,说明浙江安息香在所调查的阴湿生境中具有较强的适应能力,能够较强地利用该地的环境资源,可与其他物种形成比较平衡的竞争关系。此外,箬竹、油茶、柃木、青冈、乌药(Lindera aggregata)、构树(Broussonetia papyrifera)的生态位宽度较大,在各个样地基本都有分布。而山苍子(Litsea cubeba)、大叶冬青(Ilex latifolia)、漆树(Toxicodendron vernicifluum)、乌饭子(Vaccinium bracteatum)、檵木(Loropetalum chinensis)等树种的生态位宽度较为狭窄,仅分布于1 个样地中。
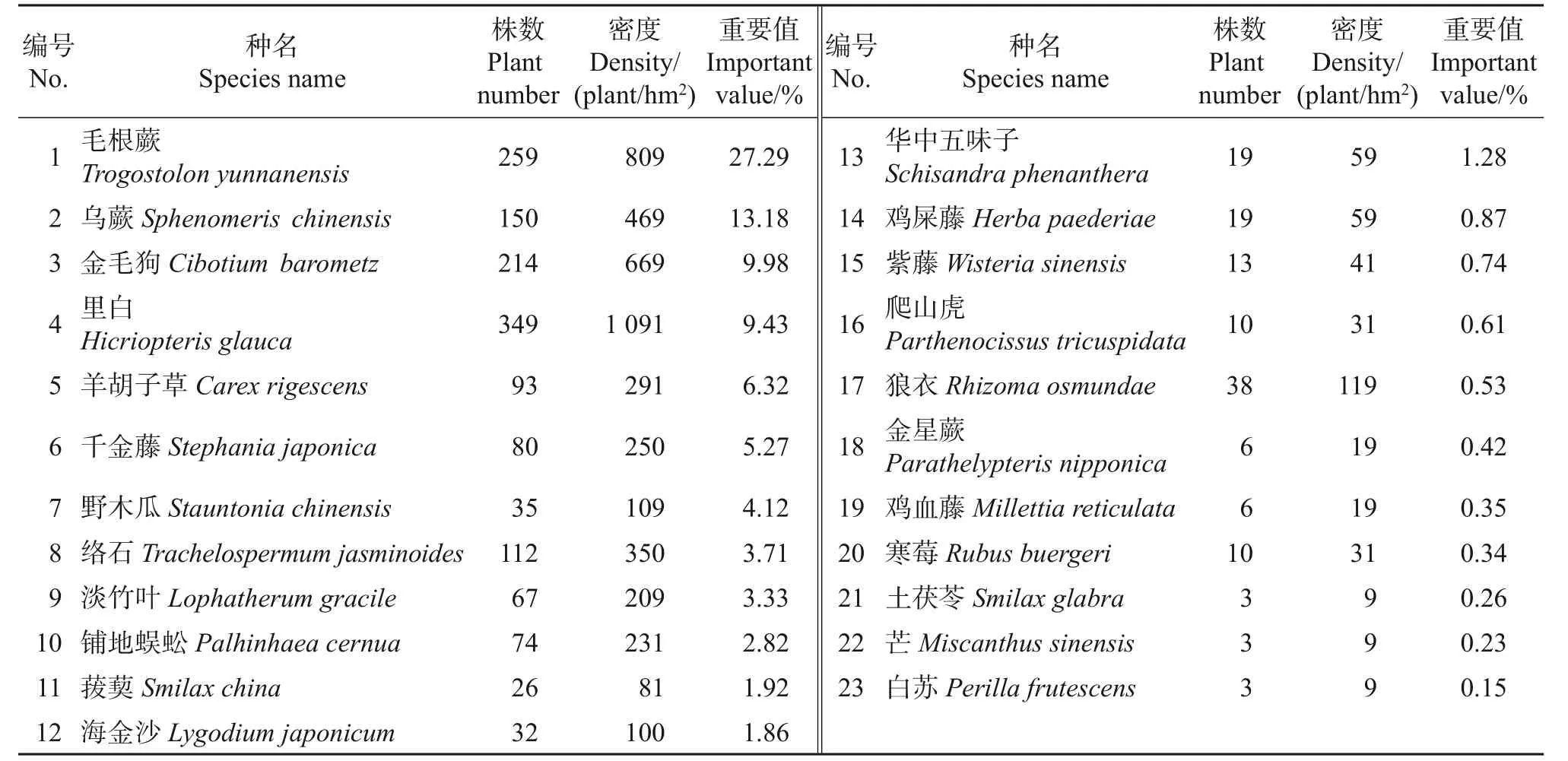
表4 草本层物种组成Table 4 Species composition of herb layer
2.4 生态位重叠特征
表6列举了乔木层中重要值≥0.5%物种的生态位重叠值,其中:生态位重叠值在0.8~0.9 的有5对,分别是木荷和白栎(Quercus fabri)、檵木和南酸枣、檵木和沙朴(Aphananthe aspera)、刨花楠和豹皮樟(Litsea coreana)以及冬青(Ilex chinensis)和沙朴;生态位重叠值大于0.5 的共有75 对,占总对数的17.24%;多数种对的生态位重叠值偏低,其中生态位重叠值小于0.1 的最多,共有266 对,占总对数的61.15%。总之,乔木树种间的生态位重叠值总体偏低。
表7列举了灌木层中重要值≥0.3%物种的生态位重叠值,其中:生态位重叠值在0.8~0.9 的有13对,分别为油茶和柃木、油茶和山矾(Symplocos caudata)、浙江安息香和柃木、柃木和山矾、柃木和桂皮等种对;生态位重叠值大于0.5 的有120 对,占总对数的27.59%;而生态位重叠值小于0.1 的也最多,共有134对,占总对数的30.80%。
一般而言,生态位宽度相对较大的物种之间生态位重叠值也较高,如:木荷和红楠,柃木和油茶,柃木和浙江安息香等,其生态位重叠值均大于0.8。但并非种群生态位宽度小的其生态位重叠值也小,如:南酸枣和冬青,刨花楠和豹皮樟,大叶紫珠(Callicarpa macrophylla)和虎皮楠等,其生态位重叠值也均在0.8 以上。这与它们自身的生态学特性、对环境的适应能力以及对资源位的要求存在一定关系[11,27]。另外,从表7 还可以看出,同属种群间生态位重叠值较小,如青冈和白栎(Quercus fabri),红楠和刨花楠,柃木和大叶柃(Eurya japonica)等,其生态位重叠值均小于0.5。
浙江安息香与柃木的生态位重叠值最大,为0.809,其次为箬竹、油茶、乌药、构树、冬青、石栎(Lithocarpus glaber)、青 冈、紫 珠(Callicarpa formosana),其值分别为0.734、0.729、0.681、0.670、0.646、0.643、0.640、0.626。而与浙江安息香生态位重叠值较小的有山苍子、漆树、大叶冬青,大小分别为0.184、0.217、0.217。

表5 木本层树种的生态位宽度Table 5 Niche breadth of species in the woody layer

表6 乔木层重要值≥0.5%物种的生态位重叠值Table 6 Niche overlap value of species with important value ≥0.5%in tree layer

表7 灌木层重要值≥0.3%物种的生态位重叠值Table 7 Niche overlap value of species with important value ≥0.3%in shrub layer
2.5 种间联结特征
浙江安息香与乔木层、灌木层的每一物种的χ2检验、结合点相关系数、联结系数、关联度指数及共同出现百分率的计算结果如表8和表9所示。
从表8 可知:浙江安息香与乔木层种对的χ2<3.841(P>0.05),说明其与所有乔木层物种联结不显著。从联结系数(AC)、结合点相关系数(PCC)看,与浙江安息香正联结的种对仅有8种,占乔木层物种数的24.24%,负联结的有23种,占69.70%。而|AC|>0.4的只有2种,其中木荷为正联结,豹皮樟为负联结。而与浙江安息香-0.40≤AC<-0.60的有17种,占乔木层物种数的51.52%;除了豹皮樟,其余|PCC|都小于0.3,也说明浙江安息香与绝大部分乔木的关联性不大或稍倾向于负联结。

表8 浙江安息香与乔木树种联结性指标Table 8 Indices of interspecific association between Styrax zhejiangensis and tree species
而从表9可知:在灌木层中,浙江安息香与山矾、紫珠和桂皮的联结系数(AC)均大于0.40,而与浙江安息香的AC值在0.20~0.40之间的有26种,占灌木层总物种数的54.17%,说明浙江安息香与灌木层其他大多数树种都存在正联结倾向。而与浙江安息香生态位重叠值最大的柃木与浙江安息香的AC 值在-0.60~-0.40之间,为-0.503,浙江安息香与大青的联结系数也达到了-0.606,说明浙江安息香与柃木、大青之间存在负联结倾向。箬竹、冬青与浙江安息香的生态位重叠值也较大,也存在负联结倾向。综上所述,柃木、大青、箬竹和冬青等物种与浙江安息香对资源的竞争较为激烈。
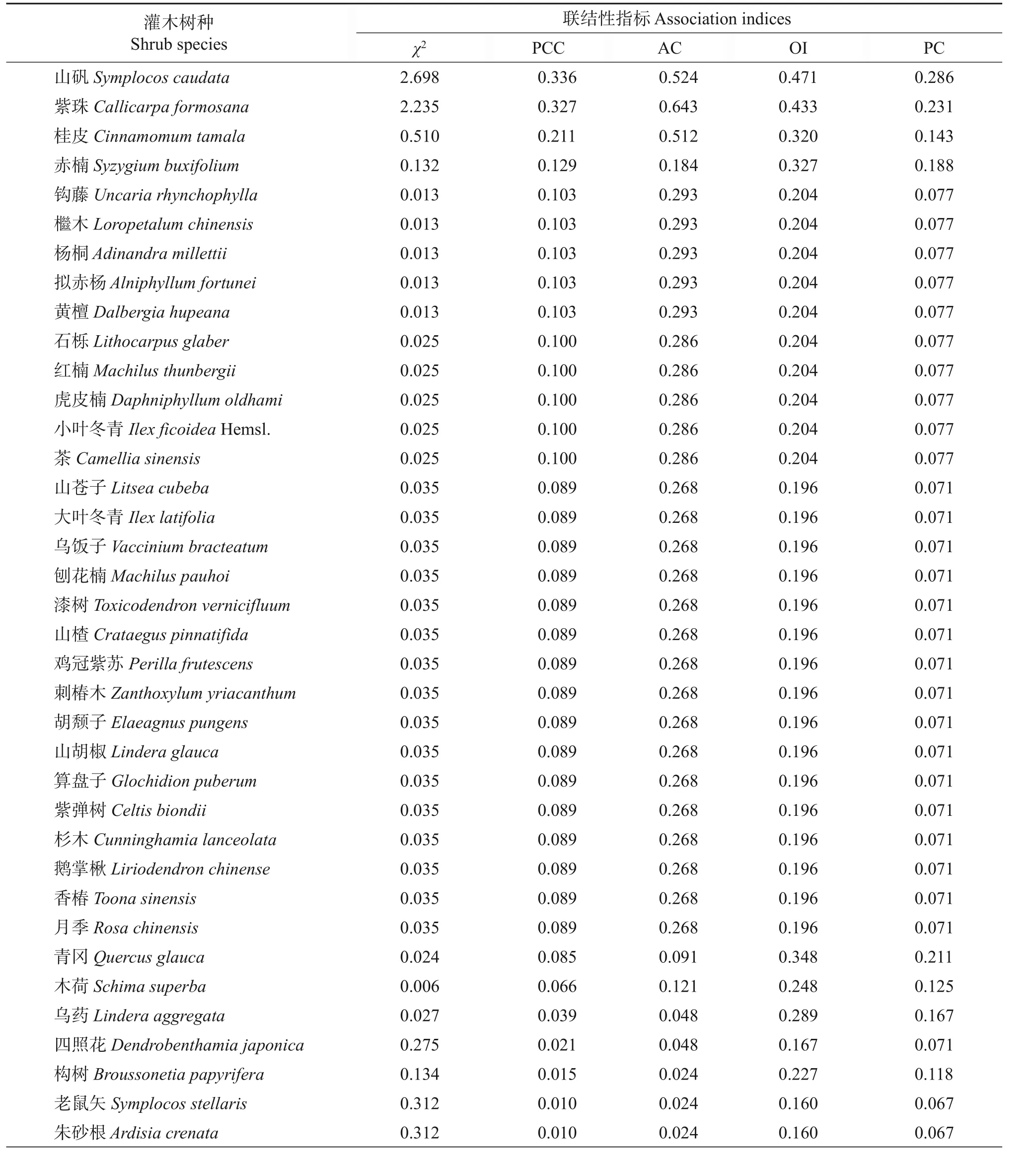
表9 浙江安息香与其他灌木树种联结性指标Table 9 Indices of interspecific association between Styrax zhejiangensis and other shrub species

表9(续) Continuation of Table 9
3 讨论
根据调查,浙江安息香所在的群落样地内乔木层和灌木层中共有植物33科49属65种2 422株,而浙江安息香数量仅占总数量的4.71%,相比2011 年的调查结果[2],其种群数量有相应的增加,新发现数量达40 丛左右。浙江安息香所在样地的群落类型为次生常绿阔叶林和针阔混交林,主要以次生常绿阔叶林为主,林分层次结构明显。其中:乔木层主要优势树种有青冈、杉木、木荷、蓝果树、红楠等;灌木层优势树种有箬竹、油茶、浙江安息香、柃木、乌药、大青等。在调查中发现,浙江安息香母树周围有少量实生小苗,高度20~120 cm,这说明浙江安息香有一定的自然繁殖能力,但由于其多生长于阴湿环境,分布区域较为狭窄,不利于开花坐果,且落花落果现象严重,种子饱满程度低,导致其种子自然繁殖能力相对较弱[2]。
种群的生态位宽度可以衡量物种对资源的利用能力和对环境的适应能力,可以间接反映种群在群落演替过程中的动态趋势[7,11]。对浙江安息香种群生态位研究发现,重要值与生态位宽度反映其在群落中的地位并非完全一致,说明物种的分布频度也可能影响生态位宽度[12,28]。重要值越大的物种,生态位不一定越宽。如在乔木层中,杉木的重要值和生态位宽度分别为8.23%、2.069,而红楠分别为4.16%和3.660。这是因为样地7为针阔混交林,杉木具有较高的重要值,说明其在样地7占有重要的资源,但其分布的样地较少;而红楠虽然在各样地所占的重要值相对较小,但其分布的样地比杉木多,导致其生态位宽度较大。在灌木层,浙江安息香、箬竹、油茶占有的资源位相同,虽然浙江安息香的重要值相对较小,但其生态位宽度最大。这是因为浙江安息香在各个样地中的重要值与箬竹、油茶相比较为平均。可见,占有较大资源量种群的生态位宽度不一定就高,而占有较大资源位或占有较大资源量且平均的种群,往往生态位宽度较大。本研究中,浙江安息香的生态位宽度较大,在研究区内分布较为均匀,表明其在该环境条件下具有较强的生态适应能力。对极小种群野生植物圆叶玉兰[11]、极小种群濒危植物黄山木兰[29]和极小种群植物缙云秋海棠[12]的研究表明,各极小种群植物的生态位宽度在群落中均较大,本研究结果与其一致。这说明在调查区域内的生态环境条件下生态位宽度大小与浙江安息香濒危原因没有必然联系,但也同时说明浙江安息香对环境要求的特殊性,即其在相对阴湿的溪流边具有较强的生态适应能力。
在群落中,2 个或更多的植物种群对某些资源的共同需求,使不同种群的生态位之间常处于不同程度的重叠状态[30]。生态位重叠值越大,物种之间对资源的利用程度越相似;相反,则表示物种之间对环境资源的需求差异大[11]。而生态位宽度与生态位重叠值之间存在一定的联系,通常,生态位宽度较大的物种之间可能出现较大的生态位重叠,这是因为生态位宽度较大的物种,其分布较广,适应能力强[31]。本研究计算结果表明,乔木层生态位重叠值大于0.5 的仅占总数的17.24%,灌木层仅占27.59%,说明浙江安息香所在的群落内大部分物种之间生态位重叠值均较低,群落物种之间对资源的需求不同,环境适应能力相似度低,有一定的生态位分化。浙江安息香与柃木、箬竹、油茶、乌药等物种有较高的生态位重叠,这反映出浙江安息香与这些树种要么对某一环境因子的要求存在互补性,要么对资源利用能力具有相似性[32];而其与山苍子、漆树、大叶冬青等物种的生态位重叠较小,这可能是浙江安息香与这些树种对生态因子的需求不同,共同资源状态也相对较少有关[12]。由于浙江安息香与柃木、箬竹、油茶分布样地多,且生态位宽度较大,因此,相应的生态位重叠值也较高。而在资源不足以满足物种的需求时,也可能因较强的竞争而表现出高的生态位重叠值[12]。根据《中国植物志》中记载,与浙江安息香生态位重叠值最大的柃木多生长于山坡、沟坎阴湿处及山坡路旁或溪谷边灌丛中[33]。在实际调查中发现,浙江安息香通常也生长于溪水边,而柃木、箬竹、油茶都喜湿润且宜生长于深厚肥沃的酸性土壤上,与实际调查中浙江安息香的生长环境较为相似。但种对之间的重叠值大小也只能反映出它们之间对资源的需求和利用的相似程度,不能完全反映其竞争关系[12,34]。
种间联结不仅能反映不同物种在空间分布上的相互关联性,还能反映各物种在不同生境中相互影响所形成的联系[35]。研究发现,浙江安息香与所有乔木层物种联结不显著。而从AC、PCC结果看,与浙江安息香呈正联结的种对占乔木层物种数的24.24%,除木荷之外,正联结系数均不大,但呈负联结的占69.70%。由此可见,乔木层对浙江安息香的生长存在着一定的影响。在灌木层中,浙江安息香与山矾、紫珠和桂皮的联结系数(AC)较大(均大于0.40),且与灌木层大多数树种存在正联结倾向。而浙江安息香与柃木、大青、箬竹及冬青之间虽然生态位重叠较大,但存在着负联结倾向,其中:与生态位重叠值最大的柃木的负联结系数(AC)达到了-0.503,与大青更是达到了-0.606,表明浙江安息香与柃木、大青、箬竹及冬青之间的种间竞争较为激烈。而浙江安息香与所有的乔木层、灌木层植物关联不显著,种间联结关系较为松散并趋向于相互独立,有较大的随机性。这与珍稀植物对开蕨以及濒危植物毛柄小勾儿茶的种间联结性的结果[14-15]相类似。
4 结论
浙江安息香不像其他濒危植物存在生态位宽度小,与大部分物种之间存在激烈竞争等问题。但浙江安息香对生长环境的要求较为特殊,能较强地适应阴湿环境。浙江安息香数量少的原因主要包括:对阴湿环境的特殊要求;在资源不足的情况下,与箬竹、柃木、冬青、大青等产生资源竞争;其林下灌草盖度大,可能会影响其种子的着床,导致幼苗数量稀少等。因此,对浙江安息香的保护措施有:
1)保护其自然生境。浙江安息香对环境具有特殊的要求,喜阴湿凉爽、阳光不直射、腐殖质丰富的酸性土壤。
2)人为干预。浙江安息香群落中,箬竹数量较多,可以将分布在浙江安息香周围的箬竹进行适当铲除,降低因资源不足的情况下箬竹与浙江安息香之间可能的竞争。
3)迁地保护。对浙江安息香进行科学的迁地抚育,扩大其分布范围,从而提高其种群数量。
4)由于浙江安息香种子的自然繁殖能力相对较弱,寻找促进其种子自然萌发的途径可能对提高其数量有一定的帮助。
此外,还需对浙江安息香的种群动态、生殖生物学特性等作进一步的研究和探讨,为保护极小种群野生植物浙江安息香提供更多的理论科学依据。
猜你喜欢
世界竹藤通讯(2023年2期)2023-05-12 09:27:08
世界竹藤通讯(2022年4期)2022-11-28 09:55:26
建筑与预算(2022年6期)2022-07-08 13:59:18
食品工业科技(2022年9期)2022-04-26 10:33:28
热带亚热带植物学报(2020年5期)2020-10-09 14:32:02
化学教与学(2019年4期)2019-05-14 04:46:40
东北林业大学学报(2015年2期)2015-03-08 05:44:46
应用化工(2014年1期)2014-08-16 13:34:08
建筑与预算(2014年5期)2014-04-11 07:29:28
中国中药杂志(2014年6期)2014-03-20 21:56:07
