
添加硫和锰对长白山森林土壤与腐殖质顽固性有机碳矿化的影响*
2020-09-16 04:54王玉哲张凤丽郑兴波韩士杰郑俊强
林业科学 2020年8期
王玉哲 张凤丽 郑兴波 韩士杰 郑俊强,
(1. 福建农林大学林学院 福州 350002; 2. 中国科学院森林生态与管理重点实验室 中国科学院沈阳应用生态研究所 沈阳 110016; 3. 河南大学生命科学学院 开封 475004)
土壤碳库是陆地生态系统中最大的碳库,其中森林土壤有机碳约占全球土壤总有机碳的70%(Panetal., 2011)。根据周转时间不同,土壤有机碳可分成不同组分,其中蛋白质、淀粉和碳水化合物等容易被微生物利用和降解,称为活性碳库(Bergetal., 2007)。木质素、腐殖质类和多酚化合物等顽固性碳组分的周转速度较慢,容易在土壤中累积,故称为顽固性有机碳,鉴于其在全球碳循环中的重要性,对其矿化的关注愈来愈多(Davidsonetal., 2006)。
近年来,工业生产将大量的二氧化硫和氮氧化物排入大气,之后不同形态的硫和氮随降水(湿沉降)或颗粒物(干沉降)进入环境,引起一系列生态环境问题(Duanetal., 2016)。我国已成为继欧美之后的世界第三大酸沉降(包括氮沉降和硫沉降)区(Liuetal., 2013)。硫沉降往往在酸沉降中占据很大比例,会引起土壤酸化,并通过改变微生物的丰度和活性从而影响土壤有机碳矿化(Vileetal., 2003)。此外,硫酸盐的还原过程也会产生CO2(Chambersetal., 2011)。目前国内关于硫对土壤有机碳矿化影响的研究主要集中在热带和亚热带森林(吴建平等, 2015; Wuetal., 2016),而对温带森林的研究相对较少。
Mn在土壤中含量丰富(Boudissaetal., 2006),自然本底浓度能达到5 mg·g-1(Hernandez-Sorianoetal., 2012)。锰过氧化物酶(MnP)是一种依赖Mn的过氧化物酶,有助于白腐菌将木质素降解为CO2和水(Hofrichter, 2002)。有研究报道凋落物在分解后期的质量损失与其Mn2+浓度呈正相关(Bergetal., 2007; Hofrichter, 2002; Virzo De Santoetal., 2009)。此外,Trum等(2011)对欧洲山毛榉(Fagussylvatica)林土壤腐殖质层的研究发现,Mn2+对土壤有机碳矿化的影响因腐殖质类型及分解程度而异。在未来全球变化背景下,大气CO2浓度升高和酸沉降导致的土壤酸化也会引起土壤Mn2+浓度升高(Bowmanetal., 2008; Ohetal., 2004; Watmoughetal., 2007)。然而,土壤Mn2+浓度增加是否能促进土壤顽固性有机碳的分解,还需进一步研究(Trumetal., 2011)。
鉴于此,本研究以长白山自然保护区的阔叶红松(Pinuskoraiensis)林、白桦(Betulaplatyphylla)和山杨(Populusdavidiana)为主的杨桦林和高山苔原的土壤以及阔叶红松林和杨桦林的腐殖质为对象,探讨硫和锰添加对土壤顽固性有机碳矿化及其温度敏感性的影响,以期为评估长白山森林碳元素的生物地球化学循环对大气硫输入的响应提供科学依据。
1 材料与方法
1.1 供试土壤来源
供试土壤采自长白山自然保护区的阔叶红松林、杨桦林和高山苔原。该区域属于典型的温带大陆性季风气候,年均气温3.8 ℃,年降水量600 ~ 900 mm,水热同期,夏季短暂多雨,冬季漫长寒冷。长白山气象观测场监测数据表明,冬季(10月至翌年2月)降水中的硫酸根月均含量从2015的3.0 mg·L-1增加到2017年的4.0 mg·L-1,说明长白山森林地区硫沉降有一定程度的增加趋势。阔叶红松林位于中国科学院长白山森林生态系统定位站一号标准地(128°05’E,42°24’N),海拔766 m,主要树种为红松、蒙古栎(Quercusmongolica)、水曲柳(Fraxinusmandsurica)和紫椴(Tiliaamurensis),优势树种的平均年龄为300年。杨桦林与阔叶红松林毗邻,于20世纪40年代林地干扰后形成。阔叶红松林和杨桦林土壤为砂壤土,属于暗棕壤,由松散的火山灰岩发育而成,0~5 cm土层的土壤有机碳含量约为13%,阔叶红松林和杨桦林0~20 cm土层土壤的有效锰含量分别为1.20~1.59和0.81~1.22 mg·kg-1。高山苔原(128°04’E,42°02’N,海拔2 150 m),坡向朝南,有机层厚10~15 cm,透水性好,pH值为5.9。有机层下矿质土壤非永久冻土,主要植被为越桔属(Vacciniumspp.)矮灌和苔藓。
1.2 土样采集
2010年5月,在阔叶红松林和杨桦林分别设置20个采样点,在高山苔原设置16个采样点。在每个采样点,分别于1 m2的区域用直径5 cm的取土器采集3个0~10 cm土层土柱,将每个采样点的3个土柱装入同一塑料密封袋,放入冰盒中冷藏保存。土壤运回实验室后,移去根系和石块等杂质后,过2 mm筛,将每个植被类型所有土壤样品混合均匀,取部分土壤样品风干后用于测定基本理化性质,其余土壤置于4 ℃下冷藏保存用于培养试验。在阔叶红松林和杨桦林长期监测标准样地内随机选择20个1 m×1 m样方,移去样方中心处地表凋落物层后,用10 cm×10 cm的不锈钢采用框采集腐殖质层样品(Fisheretal., 2000),过4 mm筛后分成2份,1份自然风干用于全碳和全氮含量测定,另一份置于4 ℃下冷藏保存以备室内培养试验使用。供试土壤和腐殖质层的基本性质见表1。

表1 供试腐殖质和0~10 cm土层土壤基本理化性质
1.3 试验设计
称取每种植被类型4 ℃冷藏保存的土壤(20 g)和腐殖质(7 g)样品各50份,置于180 mL的培养瓶中,在黑暗条件下预培养90天。为保持好氧条件,分别将土壤和腐殖质的含水率调至其生长季的野外平均水平,分别为40%和70%。预培养是为了消耗样品中的活性碳组分(Hoosbeeketal., 2007),在预培养过程中通过定期称质量来维持样品含水率。预培养结束后,用微型喷雾器将2 mL MnCl2、NaCl、MnSO4和Na2SO4溶液均匀喷洒在培养瓶中土壤和腐殖质样品表面,每种溶液处理设10个重复。本研究供试土壤和腐殖质的Mn2+含量为1.12~3.90 mg Mn·g-1有机碳,根据土壤和腐殖质样品中Mn2+添加量为3.00 mgMn·g-1有机碳的标准(Bergetal., 2007; Virzo De Santo, 2009),计算MnCl2和MnSO4溶液浓度,然后根据MnCl2和MnSO4溶液中Cl-和SO42-的摩尔浓度分别推算出NaCl和Na2SO4溶液浓度。阔叶红松林、杨桦林和高山苔原土壤样品添加的MnCl2溶液浓度分别为8.96、6.88和6.97 g·L-1、添加的NaCl溶液浓度分别为16.67、12.80和12.96 g·L-1、添加的MnSO4溶液浓度分别为10.76、8.26和8.36 g·L-1、添加的Na2SO4溶液浓度分别为10.12、7.77和7.86 g·L-1。阔叶红松林腐殖质样品添加的MnCl2、NaCl、MnSO4和Na2SO4溶液浓度分别为12.55、23.34、15.06和14.16 g·L-1; 杨桦林腐殖质样品添加的MnCl2、NaCl、MnSO4和Na2SO4溶液浓度分别为10.89、20.25、13.07和12.29 g·L-1。对照处理中加入等量的双蒸水。所有溶液的pH值调至7后再加入到土壤和腐殖质样品中。添加溶液后,立即用Parafilm 封口膜覆盖培养瓶来避免水分蒸发。将每个处理的10个重复分为两组(每组5个),一组放至25 ℃培养,另一组在35 ℃下培养。分别于第1、3、6、10、15、21、30天用注射器采集气体,每次气体采集前用橡胶塞密封培养瓶 4 h。在第10天往所有培养瓶中加入1 mL的双蒸水,以便去除水分对碳矿化的限制。用气相色谱仪(Agilent 7 890 A,美国)测定所采集气体的CO2浓度,并计算出单位时间CO2释放量,用以表示有机碳矿化速率。用相邻2次采样时间的有机碳矿化速率均值代表这一时段的平均矿化速率,将平均矿化速率乘以培养时间得到该时间段的CO2释放量,将所有采样时间段的CO2释放量相加得到CO2累积释放量,用以表示土壤和腐殖质顽固性有机碳累积矿化量。采用磷脂脂肪酸生物标记(PLFA)法测定培养结束时(第30天)土壤和腐殖质样品的磷脂脂肪酸总量,用于表征土壤微生物生物量。
1.4 数据处理
温度敏感性系数Q10的计算公式如下(Kirschbaum, 1995):
Q10=(R2/R1)10/(T2-T1)。
式中:R2和R1分别为T2和T1培养温度下土壤和腐殖质顽固性有机碳平均矿化速率。
采用重复测量方差分析方法检验锰和硫添加对土壤和腐殖质有机碳矿化速率的影响,用双因素方差分析测定植被类型和锰、硫添加对土壤和腐殖质顽固性有机碳累积矿化量和Q10的影响,用单因素方差分析检验锰和硫添加对同一植被类型土壤或腐殖质顽固性有机碳累积矿化量、Q10和磷脂脂肪酸总量的影响,采用LSD方法进一步对比两两处理之间的差异。统计分析在SPSS 22.0(SPSS Inc. Chicago, IL, USA)软件中完成,显著性水平设为α=0.05。
2 结果与分析
2.1 土壤和腐殖质顽固性有机碳矿化速率
阔叶红松林、杨桦林和高山苔原3种土壤顽固性有机碳矿化速率存在显著差异(P<0.01,表2),而不同植被类型腐殖质顽固性有机碳矿化速率没有显著差异(P> 0.05,表2)。双因素方差分析结果显示,在MnCl2和NaCl处理之间,以及在 MnSO4和Na2SO4处理之间,土壤和腐殖质顽固性有机碳矿化速率均没有显著差异,因此,图1只呈现MnCl2、MnSO4和对照3种处理土壤和腐殖质顽固性有机碳矿化速率。MnCl2和MnSO4处理之间的土壤顽固性有机碳矿化速率存在显著差异(P<0.05),MnSO4处理能提高土壤顽固性有机碳矿化速率,而MnCl2对土壤顽固性有机碳矿化速率没有显著影响。
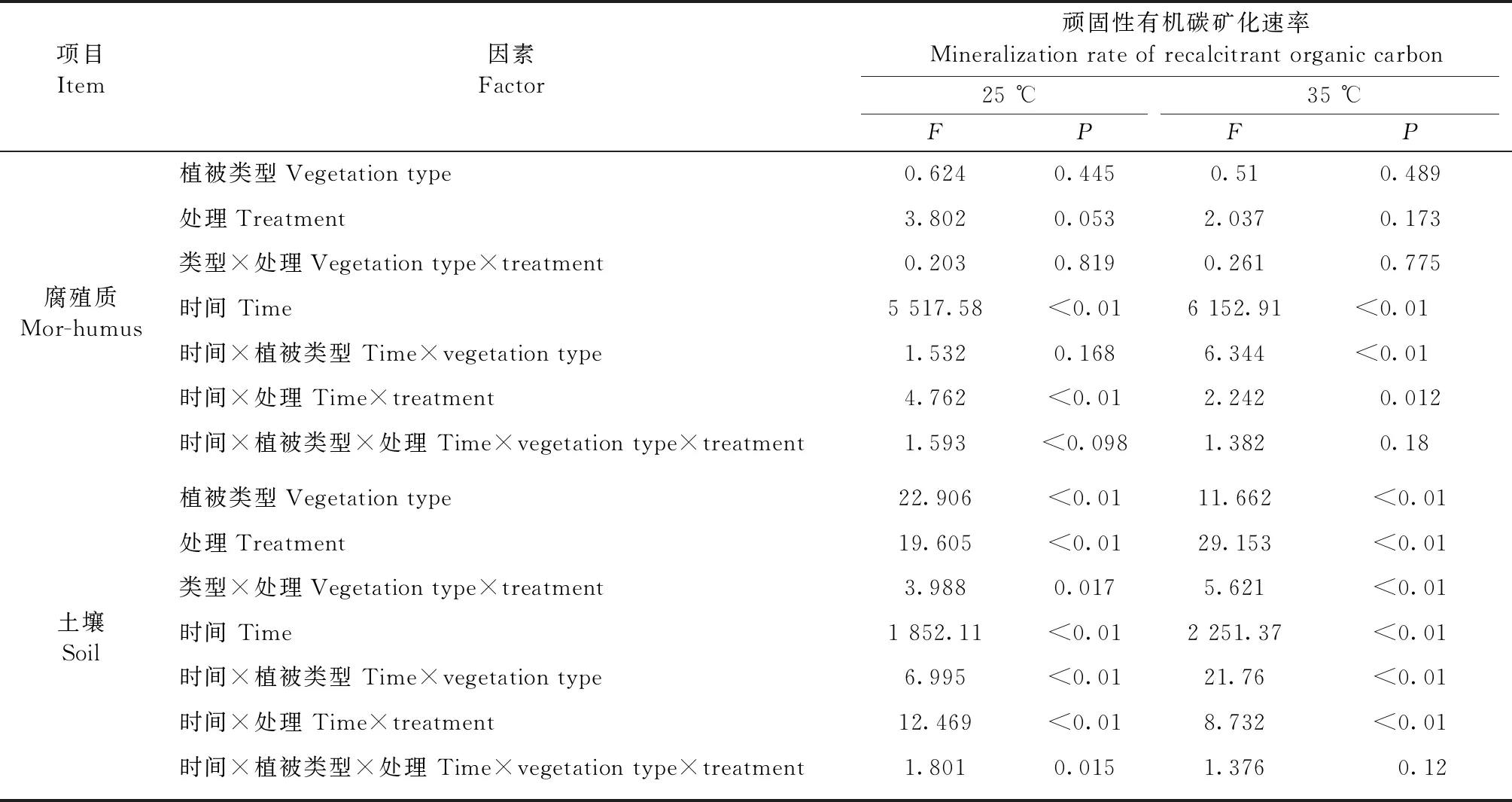
表2 土壤和腐殖质顽固性有机碳矿化速率的重复测量方差分析结果
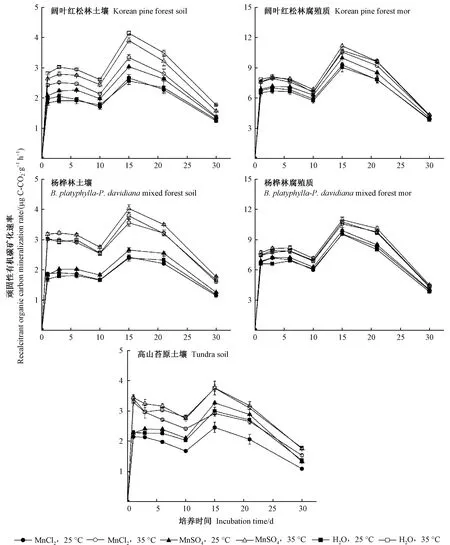
图1 长白山森林土壤和腐殖质顽固性有机碳矿化速率
2.2 土壤和腐殖质顽固性有机碳累积矿化量
在MnCl2和NaCl处理之间以及在MnSO4和Na2SO4处理之间,土壤和腐殖质顽固性有机碳累积矿化量均没有显著差异(P> 0.05,图2)。在培养温度为25 ℃时,MnSO4和Na2SO4处理显著提高了杨桦林的土壤和腐殖质顽固性有机碳累积矿化量(P<0.05,图2)。在培养温度为35 ℃时,MnSO4和Na2SO4处理显著提高杨桦林土壤的顽固性有机碳累积矿化量,相反降低阔叶红松林土壤的顽固性有机碳累积矿化量(P<0.05,图2)。

图2 长白山森林土壤和腐殖质顽固性有机碳累积矿化量
2.3 土壤和腐殖质顽固性有机碳矿化速率的温度敏感性
图3表明:土壤和腐殖质顽固性有机碳矿化速率的Q10为1.1~1.6; MnCl2、NaCl、MnSO4和Na2SO4添加对长白山阔叶红松林和杨桦林的腐殖质顽固性有机碳矿化速率Q10均没有显著影响(P> 0.05;MnCl2、NaCl、MnSO4和Na2SO4添加对土壤顽固性有机碳矿化速率Q10的影响因森林类型而异,其中显著降低阔叶红松林土壤顽固性有机碳矿化速率Q10(P<0.05),而对杨桦林和高山苔原的土壤顽固性有机碳矿化速率Q10没有显著影响; 对于高山苔原土壤,MnCl2添加处理的顽固性有机碳矿化速率Q10显著高于MnSO4和Na2SO4处理(P<0.05)。
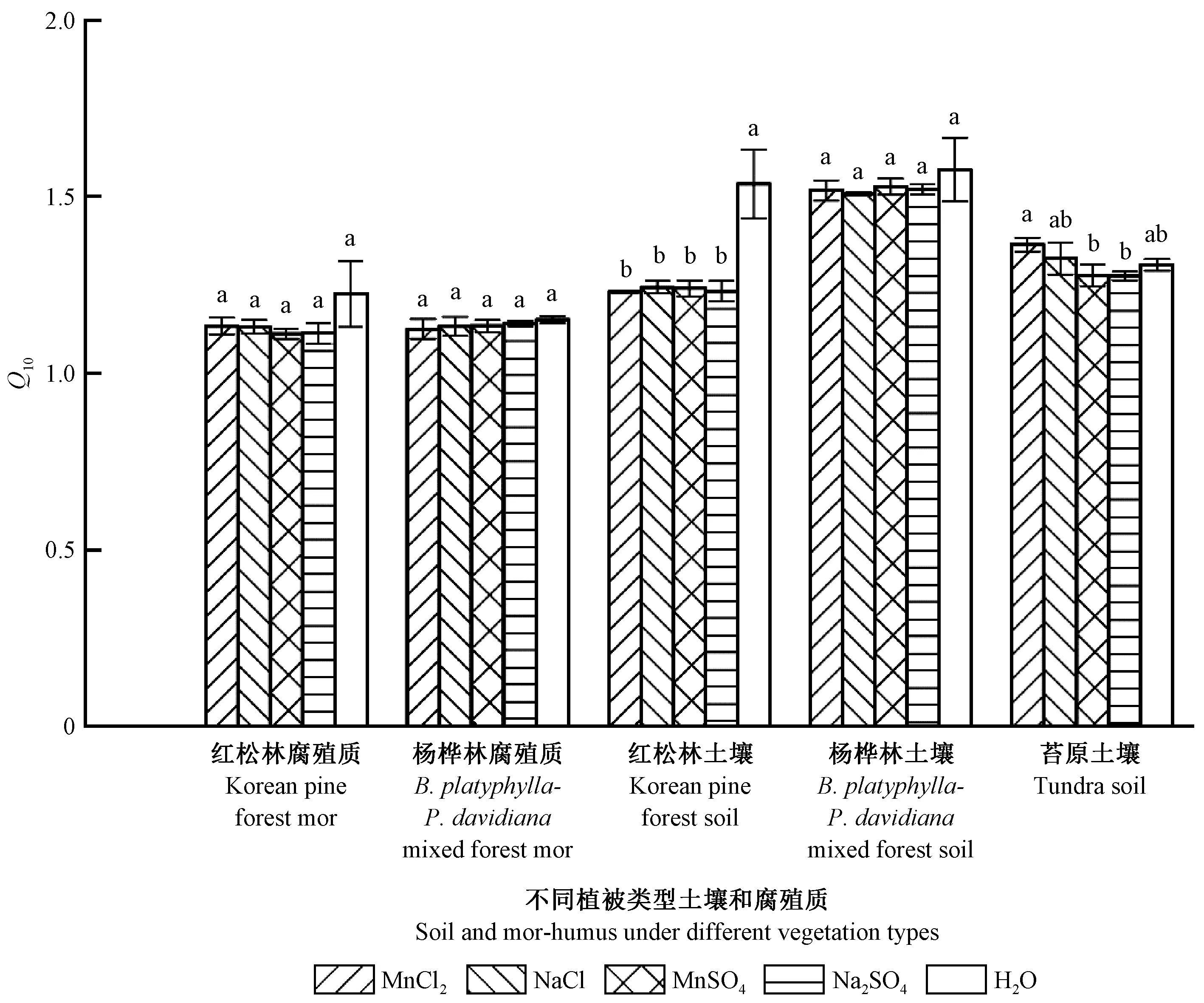
图3 长白山森林土壤和腐殖质有机碳矿化速率的温度敏感性(Q10)
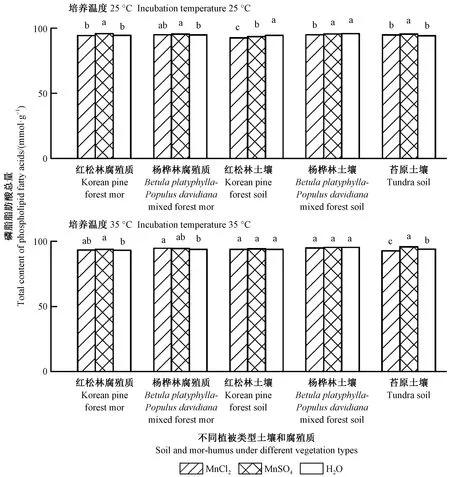
图4 培养结束时土壤和腐殖质磷脂脂肪酸总量
2.4 土壤和腐殖质磷脂脂肪酸总量
图4表明: 与对照处理相比,MnSO4添加处理显著提高阔叶红松林和杨桦林腐殖质的磷脂脂肪酸总量(P<0.05,培养温度为35 ℃时杨桦林腐殖质未达显著性差异); 培养温度为35 ℃时,MnCl2添加处理的杨桦林腐殖质磷脂脂肪酸总量显著高于对照处理(P<0.05); 培养温度为25 ℃时,MnSO4添加处理红松林土壤磷脂脂肪酸总量显著低于对照处理(P<0.05),而MnSO4添加处理高山苔原土壤磷脂脂肪酸总量在25和35 ℃培养温度下均显著高于对照处理(P<0.05)。
3 讨论
3.1 硫添加对土壤和腐殖质顽固性有机碳矿化的影响
本研究发现硫添加提高了长白山森林土壤顽固性有机碳矿化速率,说明硫是长白山森林土壤顽固性有机碳矿化的限制因素。陈晶等(2016)在落叶松(Larixprincipis-rupprechtii)林的研究也发现,土壤呼吸速率与硫沉降水平正相关(P<0.05)。然而,硫输入易引起土壤酸化,从而影响土壤微生物群落的丰度和活性(Vileetal., 2003)。有学者对亚热带森林土壤的研究发现硫添加抑制土壤有机碳矿化(吴建平等, 2015; Wuetal., 2016),这可能是由于南方土壤呈酸性,持续硫添加加剧土壤酸化,从而抑制土壤微生物活性及其调控的有机碳矿化过程。本研究的供试土壤采自温带森林,与我国南方地区相比,该地区酸沉降程度相对较低(程念亮等, 2016)。因此,硫添加在一定程度上为土壤微生物提供了养分,提高了土壤和腐殖质层的微生物生物量(图4),从而促进土壤有机碳矿化。
此外,之前研究发现硫沉降对土壤有机碳矿化的影响还与土壤pH背景值和硫添加水平有关,本研究并未设置不同的硫添加梯度,因此,在何种硫添加水平下将会抑制土壤有机碳矿化,还需今后进一步研究。由于20世纪70年代开始大气硫沉降在减少,因而其对土壤有机碳矿化的影响易被忽略。然而,在有些地区硫沉降强度还在增加(程念亮等, 2016; Sorimachietal., 2007),因此,未来在土壤碳矿化模型中应将大气硫输入作为一个重要参数,以便提高模型的准确性。
3.2 锰添加对土壤和腐殖质顽固性有机碳矿化的影响
MnP能促进土壤和凋落物中木质素和胡敏酸的降解,锰是合成MnP的重要组分(Hatakka, 1994; Hofrichteretal., 1998; Bergetal., 2007; Virzo De Santoetal., 2009)。本研究假设锰添加能促进土壤和腐殖质顽固性有机碳的矿化作用,为此,首先进行了为期90天的预培养,以消耗土壤和腐殖质中的易分解碳组分(Hoosbeeketal., 2007),预培养结束时土壤和腐殖质的碳矿化速率约为初始碳矿化速率的10%(图1),说明大部分活性碳组分已消耗完,从而认为锰和硫添加处理时测得的矿化速率是来自顽固性有机碳。与预期不同,本研究并未发现添加锰能显著改变土壤和腐殖质顽固性有机碳的矿化速率。锰添加对森林土壤和腐殖层有机碳矿化的影响与环境理化条件、森林类型以及腐殖质分解程度有关(Trumetal., 2011)。所添加的Mn2+并不能保证参与合成MnP,锰在环境中的形态与pH值和氧化还原条件有关,在本研究供试土壤pH值大于6的环境下,锰的形态也可能是MnO2和Mn2O3等锰氧化物(Lindsay, 1979),这在一定程度上限制锰参与合成MnP。本研究中锰添加对长白山森林土壤和腐殖质有机碳矿化没有显著影响,说明锰不是该地区森林土壤和腐殖质层顽固性有机碳矿化的限制因子。
4 结论
本研究首次探讨了硫和锰添加对长白山森林土壤和腐殖质顽固性有机碳矿化速率及其温度敏感性的影响,发现硫添加显著提高森林矿质土层和腐殖质层顽固性有机碳矿化速率,这是由于硫添加通过提高土壤微生物的生物量而促进了有机碳分解。锰添加对土壤和腐殖层顽固性有机碳矿化没有显著影响。硫和锰添加对Q10的影响因森林类型而异,对阔叶红松林表现为显著降低,对杨桦林和高山苔原没有显著影响。建立土壤有机碳矿化模型时应将硫输入作为一个重要参数。要准确预测森林土壤碳矿化对硫输入水平的响应,还需开展硫添加梯度的野外定位研究。
猜你喜欢
北京林业大学学报(社会科学版)(2022年4期)2023-01-04
建筑与预算(2022年6期)2022-07-08
腐植酸(2022年2期)2022-07-08
新农业(2022年9期)2022-05-16
小哥白尼(野生动物)(2021年9期)2022-01-17
——低维护的小型观赏草阔叶山麦冬
花卉(2021年21期)2021-11-11
中国林副特产(2020年5期)2020-10-09
中国环境科学(2018年10期)2018-10-29
南方农业·下旬(2018年11期)2018-02-22
腐植酸(2016年1期)2016-08-13
