
河南奶山羊线粒体D-loop区遗传多态性和遗传分化研究
2020-09-10 06:14:02韩浩园权凯闫志浩高慧军刘德奇王先宁尹慧茹赵金艳贾万里李红霞余世锋王拥庆李君
河南农业大学学报 2020年4期
韩浩园, 权凯, 闫志浩, 高慧军, 刘德奇, 王先宁, 尹慧茹,赵金艳, 贾万里, 李红霞, 余世锋, 王拥庆, 李君
(1.河南牧业经济学院动物科技学院,河南 郑州 450046; 2.西峡县畜牧局,河南 南阳 474500;3.南阳市兽药监察所,河南 南阳 473068; 4.南阳市畜牧局,河南 南阳 473068;5.沈丘县农牧科技研发中心,河南 周口 466300; 6.河南健羊牧业有限公司,河南 南阳 473000)
线粒体DNA(mtDNA)是除细胞核内遗传物质外,唯一存在于胞质细胞器内的一种遗传物质。mtDNA遵循母系遗传,mtDNA的D环控制区(D-loop)常被作为研究起源进化和遗传多样性的有效分子标记[1-2],用于探究母系亲缘关系、遗传多样性以及进化发育等。利用线粒体DNA研究山羊多态性和起源进化取得了一些有意义的成果,但同其他物种相比较,有关羊线粒体基因组及线粒体DNA的研究相对较少[3-7]。山羊的mtDNA D-loop区由16,640个碱基构成[8],D-loop控制区大小为1 212 bp,目前对山羊D-loop区的研究大多选取高变区作为研究对象。基于mtDNA的研究结果,山羊群体遗传多样性丰富[9],具有多母系起源并且在不同时间经历过不同的群体扩张。山羊线粒体DNA存在7个高度分化的单倍型分支(A, B1, B2, C, D, F, G)[10-11]。CHEN等研究了中国山羊线粒体DNA序列变异情况,分析得出中国山羊具有丰富的遗传多样性,并且首次报道了中国家山羊具有A、B、C和D 4个支系[12],而且没有明显的地理遗传结构[13-16]。山羊线粒体DNA D-loop序列多态性较高,张红平等[17]对中国16个家养山羊品种的308个个体的线粒体DNA D-loop区的研究结果表明,中国山羊的遗传多样性较为丰富。西藏山羊群体的线粒体DNA研究中共发现106个多态位点[18-19]。HOU等[20]对3个中国黑山羊群体研究也发现丰富的遗传多样性。
河南奶山羊是河南省地方奶羊品种,主要分布在河南省的开封、南阳、洛阳和郑州等地区。河南奶山羊是1904年意大利天主教神父潭维新(Noe-Joseph Tacconi,1873—1942年)将萨能奶山羊引入河南省开封和南阳地区,与当地地方山羊品种杂交,并经过长期选育形成的乳用山羊品种。该品种体型中等,耐粗饲、对农区秸秆资源利用率高,繁殖率高,适合中部平原地区的气候环境。目前,对于河南奶山羊遗传关系的研究尚为空白,为了解河南奶山羊群体构成与遗传多样性,本研究以河南奶山羊、萨能奶山羊、关中奶山羊和伏牛白山羊作为研究对象,通过分析mtDNA D-loop区全序列研究其遗传多样性,并利用mtDNA D-loop区的遗传特性及序列,分析河南奶山羊与萨能奶山羊、关中奶山羊和伏牛白山羊的遗传关系及遗传分化,了解河南奶山羊的遗传资源状况,对河南奶山羊品种资源的保护、开发和合理利用提供理论依据。
1 材料和方法
1.1 样本采集
本研究采集河南奶山羊(n=20)、萨能奶山羊(n=12)、伏牛白山羊(n=12)和关中奶山羊(n=4),共48个山羊个体的外周血样本(表1)。血样均为随机抽测,利用 EDTA 抗凝管颈静脉采血5 mL,-20 ℃保存备用。河南奶山羊样品采自河南健羊牧业有限公司,萨能奶山羊和关中奶山羊样品采自洛阳妙妙牧场有限公司,伏牛白山羊样品由河南省内乡县畜牧站提供。
1.2 基因组DNA提取及PCR扩增
利用动物基因组DNA提取试剂盒(北京艾德莱生物科技有限公司)提取血样基因组DNA,4 ℃保存备用。由生工生物工程(上海)股份有限公司合成山羊D-loop区特异性引物,F:5′-AACCACTATTAACCACATCTA-3′;R:5′-CACTTACCATGTAAAAGACCC-3′。PCR 扩增反应体系为25 μL:DNA模板(100 ng·μL-1)1μL,正向引物和反向引物(10 μmol·L-1)各为1 μL,Taq PCR Master Mix 12.5 μL(1 U Taq Polymerase、5 μmol·L-1dNTP、0.2 mmol·L-1Tris-HCl、1 mmol·L-1KCl、0.03 mmol·L-1MgCl2),ddH2O 9.5 μL。PCR扩增条件为:95 ℃预变性3 min;95 ℃变性30 s,53.5 ℃退火30 s,72 ℃延伸45 s,34个循环;72 ℃延伸1 min 30 s,4 ℃保存。利用琼脂糖凝胶电泳对PCR产物进行检测,确认产物大小与目的片段一致,送往生工生物工程(上海)股份有限公司测序。
1.3 数据分析
经生工生物工程(上海)有限公司双向测序,利用Lasergene软件包中的SeqMan软件将上下游测序序列拼接,去除测序质量差的杂峰部分;利用MEGA6.0软件[21]自带的ClustalW多重比对对拼接好的序列进行分析,利用截齐后的1 140 bp序列构建UPGMA系统进化树,更改自举检验重复数为1000;分析河南奶山羊和伏牛白山羊、萨能奶山羊以及关中奶山羊间的遗传关系,利用K2P(Kimura 2-parameter)模型[22]计算品种间的遗传距离;导出MEGA 6.0比对后的FASTA格式序列,并导入DnaSP V5[23]分析4个山羊品种D-loop区的DNA多态性:单倍型多样度(Hd)、核苷酸多样度(Pi)和中性检验[24],以及品种间基因流和遗传分化:种群间核苷酸平均差异数(Kxy)、核苷酸歧义度(Dxy)、遗传分化系数(GST)和固定系数(FST)等遗传学参数;并利用DnaSP V5进行单倍型分析,生成单倍型数据,导入NETWORK 10.0.0.0软件(Fluxus Technology Ltd., Kiel, Germany)中构建单倍型网络图。
2 结果与分析
2.1 线粒体D-loop扩增产物检测
4个山羊品种48个样本经由DNA提取和PCR扩增得到图1所示琼脂糖检测结果,显示扩增效果良好,条带清晰明亮,无杂带,条带大小约为1 200 bp,与预期目的条带大小吻合,可用于后续测序及分析。
2.2 线粒体D-loop遗传多样性及单倍型分析
测序拼接后,将48个个体D-loop序列截为1 140 bp,其中单态位点为1 078个,突变位点为43个,缺失位点为19个。遗传多样性分析表明,4个山羊品种总单倍型多样度为0.903,各品种单倍型多样度范围为0.391~0.814,总核苷酸多样度为0.007 50,4个品种核苷酸多样度的范围为0.000 35~0.004 64,河南奶山羊均表现为最高的单倍型多样度和核苷酸多样度。4个山羊品种的核苷酸平均差异数为8.402(表1)。单倍型分析共发现16个单倍型,其中河南奶山羊有9个单倍型(Hap 4~12),萨能奶山羊有3个单倍型(Hap 1~3),关中奶山羊和伏牛白山羊分别有2个单倍型(Hap 13~14和Hap 15~16)。单倍型网络图明显分为4个单倍型组,分别代表河南奶山羊、萨能奶山羊、关中奶山羊和伏牛白山羊(图2)。河南奶山羊Tajima’s D 中性检验P<0.05,显著偏离中性,表明河南奶山羊经历过群体扩张。萨能奶山羊、关中奶山羊和伏牛白山羊Tajima’s D 中性检验均不显著(P>0.1),符合平衡发展,未出现群体大规模扩张或瓶颈现象,群体大小保持相对稳定。
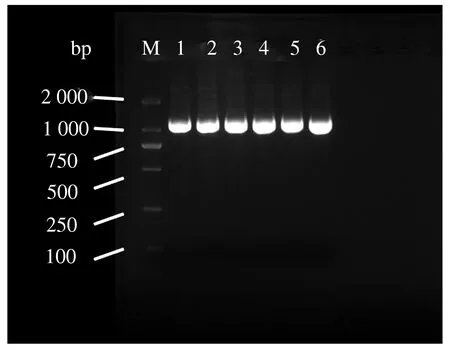
注:M.Marker;1、2.河南奶山羊、3、4.萨能奶山羊;5.关中奶山羊;6.伏牛白山羊。

表1 4个山羊品种遗传多样度及单倍型Table 1 Genetic diversity of four goat breeds
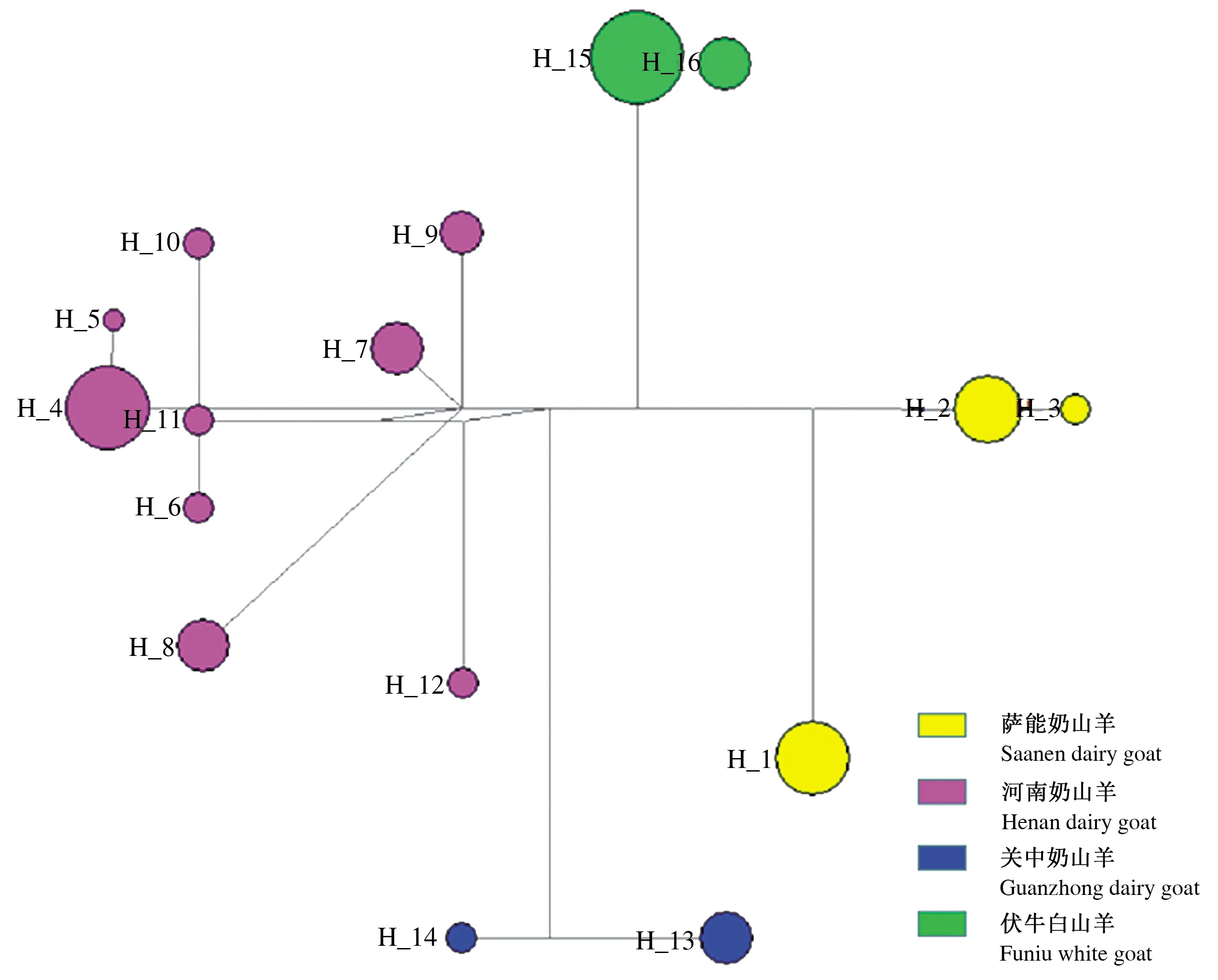
注:H_1~16代表单倍型Hap 1~16。
2.3 基因流与遗传分化分析
品种间基因流与遗传分化分析结果表明,核苷酸平均差异数范围为9.333 33~13.666 67,核苷酸岐异度变异范围为0.008 33~0.012 19,萨能奶山羊与伏牛白山羊间的核苷酸平均差异数和歧异度最低,而萨能奶山羊和河南奶山羊之间的最高。河南奶山羊与伏牛白山羊的核苷酸平均差异度和歧异度最小,其次是萨能奶山羊,与关中奶山羊差异最大(表2)。遗传分化系数范围为0.135 65~0.360 10,固定系数范围为0.583 23~0.935 50,关中奶山羊与伏牛白山羊间的遗传分化系数和固定系数最高(表2)。河南奶山羊与关中奶山羊分化系数最小,代表两者间遗传多态性差异最大,而与伏牛白山羊间的分化系数最大,代表两者之间核苷酸差异最小,与前述核苷酸平均差异数和歧异度的结果相符,也与固定系数结果基本一致。
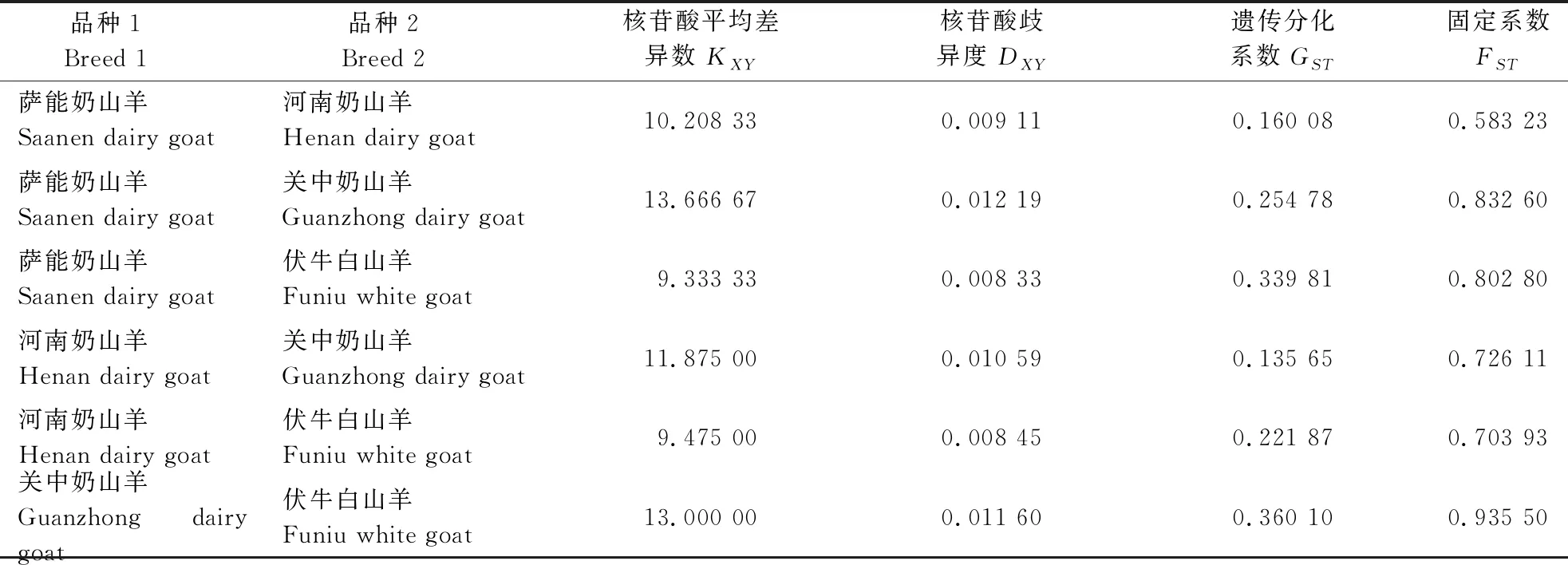
表2 遗传分化与基因流相关参数Table 2 Genetic differentiation and gene flow of four breeds
2.4 系统发育树构建与遗传距离分析
4个山羊品种间系统发育树(图3)表明,河南奶山羊首先与伏牛白山羊聚为一支,其遗传距离为0.008 553 6;接着河南奶山羊与萨能奶山羊聚类,其遗传距离为0.009 136 8;最后与关中奶山羊聚类,其遗传距离为0.010 649 1。系统发育树聚类结果与遗传距离分析结果相符(表3),证明了河南奶山羊与伏牛白山羊遗传关系最近,接着是萨能奶山羊,离关中奶山羊最远。该结果与遗传分化分析结果一致。

注:H.河南奶山羊;SN.萨能奶山羊;G.关中奶山羊;F.伏牛白山羊。
3 结论与讨论
3.1 河南奶山羊遗传多样度
核苷酸多样度和单倍型多样度是评价群体遗传分化的重要指标,数值越大,表明群体遗传多样性越丰富。本研究的遗传多样性分析表明,3个奶山羊品种单倍型多样度为0.429~0.814,核苷酸多样度为0.001 14~0.004 64。本研究结果与皮秀霜[25]对关中奶山羊、文登奶山羊和崂山奶山羊遗传多样性研究结果相一致,关中奶山羊、文登奶山羊和崂山奶山羊的平均核苷酸多样度为0.009 8±0.000 9,平均单倍型多样度为0.865±0.010。中国奶山羊品种核苷酸多样度低于云贵川地方山羊品种,川渝黔地区山羊线粒体D-loop遗传多样性

表3 河南奶山羊、伏牛白山羊、萨能奶山羊和关中奶山羊间的遗传距离Table 3 The genetic distance of Henan dairy goat, Funiu white goat, Saanen dairy goat and Guanzhong dairy goat
研究表明核苷酸多样度范围为0.004~0.040[26-28],云南地方山羊单倍型多样性为0.985,平均核苷酸多样性为0.019 15[29-30],显示出川渝黔地区山羊品种较高的遗传多样性。中国奶山羊品种均以萨能奶山羊为父本,经过与地方品种长期的杂交改良和选育形成,血统来源相对单一,可能是造成中国奶山羊多样度较低的原因。
本研究结果表明,河南奶山羊单倍型多样度和核苷酸多样度最高,依次为萨能奶山羊、关中奶山羊和伏牛白山羊,从品种形成来看,河南奶山羊母本来源包括槐山羊、伏牛白山羊和尧山白山羊,品种数量多、群体大,而萨能奶山羊、关中奶山羊和伏牛白山羊母本来源相对单一、数量小。因此,河南奶山羊单倍型多样度和核苷酸多样度远高于其他3个品种。
本研究在河南奶山羊、关中奶山羊、萨能奶山羊和伏牛白山羊界定16个mt DNA D-loop单倍型,每个品种均存在特异单倍型,该结果与王杰[31]、赵中权等[32]的报道相一致,说明中国奶山羊不同群体都具有独特的mt DNA D-loop单倍型,不同品种间共享单倍型较少,类群单倍型丰富。
3.2 河南奶山羊群体扩张
本研究中河南奶山羊Tajima’s D值为0.190 07,P<0.05,差异显著,表明河南奶山羊存在过群体扩张,ZHAO等研究发现中国的奶山羊品种支系A显著偏离中性,曾经历群体扩张[16,20]。历史对于线粒体DNA的遗传变异模式和遗传多样性都具有深远影响,对畜禽群体来说,环境条件的改变,可导致随机漂变和等位基因频率的改变,当群体数量发生大的变化时,可以检测到其是否发生过群体扩张。目前,河南奶山羊规模场主要分布在南阳市西峡县,西峡县政府在近些年来将河南奶山羊作为畜牧业主导产业进行大力发展,河南奶山羊得到了快速的扩张和发展。
3.3 河南奶山羊遗传距离和遗传分化
本研究发现河南奶山羊与伏牛白山羊遗传距离为0.008 553 6,与萨能奶山羊聚类,其遗传距离为0.009 136 8;最后与关中奶山羊聚类,其遗传距离为0.010 649 1,表明河南奶山羊和伏牛白山羊母系遗传关系最近,其次是萨能奶山羊,离关中奶山羊最远,本研究结果从遗传角度证实了河南奶山羊是以萨能奶山羊为父本,以伏牛白山羊为母本选育而成。皮秀霜[21]发现中国奶山羊品种间的遗传距离变异范围为0.004 7~0.013 8,本研究结果与该结果相近。本研究发现河南奶山羊与萨能奶山羊、关中奶山羊间存在单倍型特异性与明显的遗传分化,证明河南奶山羊与其他奶山羊间存在明显差别。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22 00:33:26
肝博士(2022年3期)2022-06-30 02:48:28
今日农业(2021年15期)2021-11-26 03:30:27
Journal of Sport and Health Science(2019年6期)2019-11-26 07:30:53
作文周刊·小学一年级版(2017年46期)2018-01-16 21:55:21
小天使·一年级语数英综合(2017年4期)2017-04-18 18:09:55
兽医导刊(2016年6期)2016-05-17 03:50:53
中国畜牧兽医文摘(2015年9期)2015-12-29 03:38:17
动物医学进展(2015年10期)2015-12-07 05:46:21
癌变·畸变·突变(2014年2期)2014-03-01 04:39:42
