
纳豆激酶溶栓功效与酶活测定研究进展
2020-09-06 05:35李五林
科学导报·学术 2020年79期
关键词:结构
李五林
【摘 要】纳豆是日本传统发酵食品,纳豆含有多种活性成分,其中纳豆激酶对于血栓的去除有极强的功效,但纳豆在中国的的推广进展缓慢,民众对于纳豆的结构、功效、酶活的判定等不够了解。本文概述了纳豆激酶的结构特性,纳豆激酶的溶栓机理、抗血栓实验效果与酶活力的测定方法,并对纳豆激酶的研究做出展望,以期增进公众对纳豆激酶的认知,并为进一步的研究奠定基础。
【关键词】纳豆激酶;结构;抗血栓效果;酶活性测定方法
1 引言
血栓类疾病对患者的个人生活影响极大,如中风患者常常突然昏倒、言语障碍或失语、偏身麻木甚至半身不遂,心肌梗塞容易造成心脏骤停、心源性休克甚至心脏破裂。根据世界卫生组织WHO统计,截止2018年,我国以血栓为主要诱因的心血管病患者数量达2.9亿,其发病死亡总数占居民总死亡数的40%,而在全球,直接或间接由血栓栓塞类疾病造成的死亡人数已达总死亡人数的51%,远高于癌症等其他疾病。因此抗血栓成为了公众和科学界关注的焦点问题,而直接具有溶栓作用的相关生物酶的研究也成为了领域热点。目前已开发应用的生物溶栓酶有链激酶和尿激酶两种,但其皆存在不可避免的缺陷。链激酶易激活全身纤溶系统,造成人体纤维蛋白原含量大幅下降和全身出血,同时链激酶存在抗原性,部分病人会产生过敏反应。尿激酶是我国最主要的抗血栓药物,但其对血栓骨架纤维蛋白特异性较低,不专门溶解血栓纤维蛋白,存在出血倾向,同时给药剂量难以控制,另外尿激酶价格昂贵,而血栓患者往往需要长期服药,绝大部分患者根本无法承受。1987年,日本须见洋行博士灵光一现,想到其本土食品纳豆的发酵过程中降解的主要物质为纤维蛋白,而这正是血栓的核心骨架成分。在后续实验中,他果然发现了一种有极大溶栓活性的物质,其活性相当于尿激酶的四倍之多,须见洋行将其命名为纳豆激酶。此后,一系列关于纳豆激酶更深入的研究得以展开,结果表明,纳豆激酶的溶栓活性大大高于已有生物酶,同时其具有溶栓特异性,专一降解血栓骨架中的纤维蛋白,引起出血风险极低。同时其价格低廉、稳定性好、对人体安全性高,有望替代已有生物酶,从而受到更广泛关注,日本更是倡议国民从小学起便开始坚持食用纳豆。本文从活体抗血栓实验、结构与生化特性、溶栓机理和活力测定方法几个方面对纳豆激酶的相关研究进行系统总结,并对一些问题进行分析,以期更全面的了解纳豆激酶,为后续进一步的研究奠定基础。
2纳豆激酶的结构与生化特性
2.1 纳豆激酶的结构
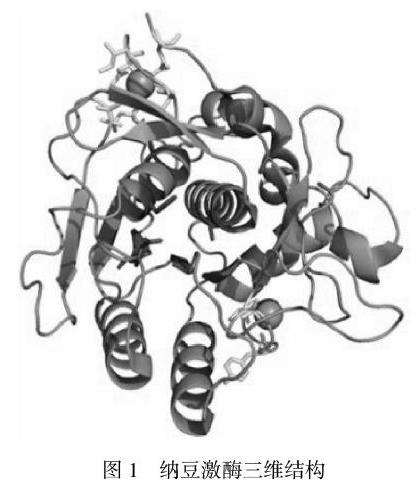
纳豆激酶(Nattokinase简称NK)又名枯草杆菌蛋白酶NAT,屬碱性丝氨酸蛋白酶,由枯草杆菌所属的纳豆菌代谢时所分泌,在细胞外进行降解作用。酶分子为单链多肽,无二硫键,共含275个氨基酸,分子量为27.7 kDa。其与底物结合的中心由三个氨基酸(Ser125、Leu126、Gly127)构成,催化活性中心为(Asp32、His64、Ser221)。其与枯草杆菌蛋白酶E在氨基酸序列的同源性达99.5%,仅仅在85和192位的氨基酸残基存在差异,与枯草杆菌蛋白酶BPN的同源性达86%,但除NK外其他枯草杆菌蛋白酶均未发现纤溶活性,分析原因可能与蛋白质的三级结构差异有关。2010年,NK的X-射线晶体结构被发现,如图1。由晶体结构分析得出,NK中存在7个α-螺旋结构和9个β-折叠结构,它们相互交错,呈现出NK整体的球状结构,其活性位点位即于球状蛋白表面的沟槽内。另外,
NK结构中存在2个Ca2+结合位点,与蛋白质三级结构的稳定性有关。在NK肽链中还存在发光氨基酸色氨酸、酪氨酸和苯丙氨酸,能为蛋白质的光谱研究提供基础。
2.2 纳豆激酶的生化特性
NK适宜pH为5-12,最佳pH为7-9,适宜温度为60 oC以下,最佳温度为35-45 oC,而人体血液pH 约为7.35-7.45,血液温度近似37.5-38 oC,这与NK的最佳环境极其一致,十分有利于NK表现最佳活性。但酸性环境和高温都会让NK迅速失活,因此一般纳豆激酶产品利用肠衣包裹,避免其受到胃酸破坏,另外,有研究表明,单纯食用纳豆时,其中的纳豆激酶在纳豆粘液、食物中的蛋白质、油脂等物质的包裹下,能在酸性环境中仍保持部分活性。低温对NK的影响不大,在-20 oC的环境下,反复冻融NK五次,其仍存留95%以上的活性。因此低温冷藏可以作为纳豆的有效储存方式。经Svunsson柱型电泳法,等电聚焦测定出NK的等电点为8.6±0.3,为NK的分离纯化奠定了基础。金属离子对NK活性有一定影响,Na+、K+、Mg2+、Co2+、Ca2+、Tb3+等能提高其活性,Cu2+、Hg2+、Zn2+、Fe3+、Al3+则会不同程度的抑制其活性。这可能与金属离子影响了NK的聚集性质有关。
3 纳豆激酶的溶栓机理
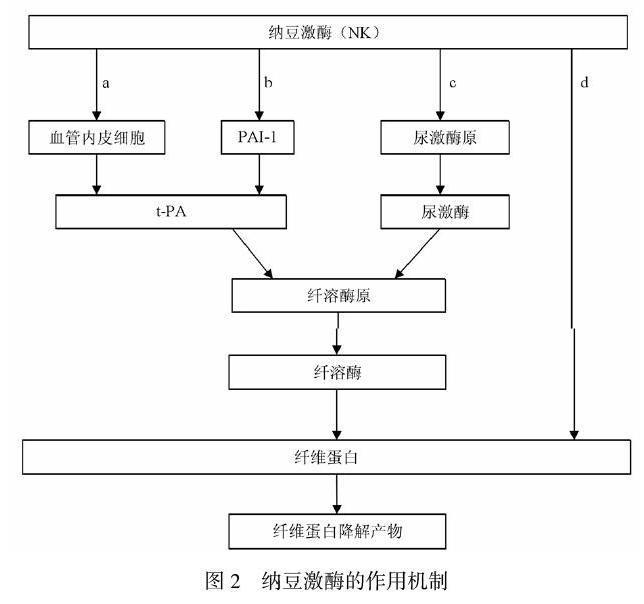
正常人体存在对立调控的凝血溶血系统,凝血系统保证血管壁破裂时及时填堵,溶血系统则保证血液畅通。当凝血系统过强或溶血系统过弱时,便会形成血栓。当人体血因外部物理因素等造成血管内皮损伤后,体内凝血系统激活,激活各种凝血因子,使血小板大量聚集,同时促进纤维蛋白原转化为纤维蛋白,并形成交联结构作为血栓骨架,与聚集的血小板一起最终构成血栓。人体内部的纤溶酶是唯一能降解纤维蛋白从而促进血栓溶解的物质,大多数溶栓药物如尿激酶、链激酶等皆通过促进纤溶酶的形成发挥溶栓作用。而纳豆激酶除激活纤溶酶发挥作用外,还能直接作用与交联纤维蛋白,发挥溶栓作用。NK作用机制见图2,主要有四种途径:a,直接刺激血管内皮细胞,促进纤溶酶前体的调控因子的释放,间接促进纤溶;b,抑制a途径中调控因子的抑制因子,避免纤溶受到阻碍;c,刺激尿激酶前体转化为尿激酶,再由尿激酶激活纤溶酶前体转化为纤溶酶,以此促进纤溶;d,直接作用于纤维蛋白,降解血栓骨架,完成纤溶,这也是NK最主要的作用途径。
4 活体抗血栓实验效果
4.1 直接抗血栓实验
研究者通过大鼠静脉直接注射纳豆激酶,发现NK具有明显溶栓效果,且溶栓活性为血纤维蛋白溶酶的4倍以上。在小鼠颈部动脉注射醋酸生成血栓的实验中,分别静脉注射等摩尔数的纳豆激酶、纤溶酶及弹性蛋白酶,1h测定的小鼠血流再通率分别为(62.0±5.3)%、(15.8±4.5)%及0%,NK的血流再通率大大高于纤溶酶和蛋形蛋白酶,且绝大部分实验组实现血流再通,体现了NK极高的体内溶栓活性。
另外,在突发性血栓实验中,NK表现出了极佳的预防功效。研究者通过肠衣包裹纳豆激酶口服给药,确保其直达肠道吸收,持续进药15000IU/Kg/d,7天后通过FeCl3诱导模仿突发性血栓,比较实验组和生理盐水给药的对照组的血栓重量(2.8mg)发现,实验组的突发性血栓重量(1.4mg)大幅度降低,表明NK能有效抵抗和减缓突发性血栓的形成。
4.2 间接抗血栓实验
在研究小鼠对纳豆激酶的肠道吸收实验中,通过小鼠十二指肠给药,在3-5h后即在血液中检测到纳豆激酶,表明NK可经肠道吸收。研究者以大鼠为实驗动物研究NK的体内溶栓作用底物,发现纳豆激酶发挥作用时直接降解血栓骨架-交联纤维蛋白,但对纤维蛋白前置物纤维蛋白原不敏感,这种直接作用十分有利于NK发挥其溶栓效果,且不易引起出血倾向。以大鼠为实验动物研究NK的抗动脉粥样硬化作用实验中,在大鼠血管内皮损伤的前三周到后三周持续给与与含NK的纳豆提取物,比较血管损伤开始和三周后的血管内膜/中层厚度值,发现实验组(0.06±0.01)较对照组(0.15±0.03)数值明显减小(P<0.05),且尤球蛋白凝块溶解时间缩短,这表明NK能有效抑制大鼠血管壁血栓的形成。
另有研究者利用血液中的血栓骨架-纤维蛋白的降解产物D-dimmei和FDP的含量表征纳豆激酶的抗血栓效果,通过注射角叉菜胶在小鼠体内形成血栓,再将纳豆激酶分低、中、高剂量组连续灌胃一周,同时设置生理盐水对照组,结束后取腔静脉血分析血液中FDP、D-dimmer浓度。结果显示,低剂量组FDP、D-dimmer含量与对照组相近,未表现明显差异,而中剂量、高剂量组FDP、D-dimmer含量分别为对照组的2倍和3倍多,差异明显。这表明中高剂量的纳豆激酶降解了血栓,有很好的抗血栓疗效,且其抗血栓作用表现出明显的剂量效应关系。
4.3人体临床效果
通过对心血疾病患者的研究发现,纳豆激酶能够降低患者出现心肌缺血的概率,减小心机梗死的面积。在一项志愿者参与的实验中,研究者利用肠溶胶囊给药避免NK受到胃酸腐蚀,对照组服用煮熟大豆,对比实验组和对照组的优球蛋白溶解时间(ELT)、溶解活性(EFA)和纤维蛋白降解产物(FDP),结果表明实验组志愿者血液中ELT明显缩短,EFA大大提高,同时FDP显著增加增加,这表明NK促进了人体中血栓的降解,显现出较强的抗血栓活性。2015年,国家药品批准纳豆激酶作为一种功效成分上市,纳豆激酶得到官方认可,继而市场上出现大量纳豆激酶产品,其中以胶囊、软胶囊、微囊等最为多见。其利用肠溶外衣结构,避免纳豆激酶受到胃部极酸环境(pH约 0.9-1.8)的破坏,从而保证NK有效的肠道吸收。
5 纳豆激酶活力的测定方法
纳豆激酶活力测定主要基于其三种特性,纤溶特性、降解特定枯草杆菌底物特性和抗原特性。相应其活力测定主要有三类方法:a,通过溶解模拟血栓的纤维板从而形成溶圈,比较样品溶圈面积和标准酶溶圈面积,根据面积与活力关系得到样品酶活的纤维板法及其改进的各种方法;b,通过直接降解几种特定底物,根据生成产物的速率定义得出酶活,产物定量方式有表观明显现象如气泡、通过产物测定吸光度或经显色反应后测定吸光度等;c,根据NK的抗原特性用特异性抗体捕获,与另一抗体二次结合后以过氧化物酶与二次抗体的反应确定一次结合的抗体量,进而确定NK的活力的酶联免疫吸附法。
5.1 纤维板法及其改进
作为NK发现者验证其纤溶活性的经典方式,纤维板法及其改进的各种活力测定方法能直接反应NK的纤溶活性,且其操作简单易行,并可以同时测定多种样品,因此是目前最为常用的NK活力检测方法。该方法利用凝血酶与牛纤维蛋白原反应,在平板中生成纤维蛋白,打孔后将一定体积适宜酶活的样品酶注入孔内,37 oC恒温孵育18h测定溶圈面积,再将溶圈面积带入面积-酶活标准曲线中计算样品酶活。其中标准曲线的建立方法为:配置不同浓度的尿激酶,再分别于纤维平板中恒温孵育出对应的不同溶圈面积,面积与酶活成线性关系,从而得到溶圈面积-酶活标准曲线。此法也存在许多缺陷,由于溶圈面积是表观值,而一般形状又不完全规则,因此受个人测量误差影响较大,而溶圈面积受孵育条件与酶纯度干扰,孵育时间、温度、孵育平板、酶杂质都会对溶圈面积造成影响。另外,纤维平板所用纤维蛋白原价格昂贵,实验成本较高。这些因素导致纤维板法数据波动性大,且具有一定人为主观性,难以多次重复等问题。
纤维板法的改进主要有:a,增加琼脂糖基底的琼脂糖-纤维板法;b,增加聚丙烯酰胺基底并用特定模具代替玻璃或塑料皿,辅以指示剂放大蛋白降解效果的聚丙烯酰胺纤维蛋白胶板法;c,直接测定纤维蛋白形成初始加入NK前后,吸光度变化的血清板法。增加琼脂糖基底能有效避免塑料平皿底部差异和不平现象,且有效减少纤维蛋白用量,聚丙烯酰胺和特定模具则进一步提高了制板精度,能一定程度提高数据稳定性。血清板法在血清板小孔中形成微型血栓,在加入NK反应前后测定OD655,操作更加简便快捷,但一般紫外分光光度计无法直接测定血清板样品,设备要求较高。
5.2通过降解特定底物确定酶活的方法
NK作为一种纤维蛋白降解酶,可以作用于纤维蛋白,同时NK属枯草芽孢杆菌,能降解部分适应于枯草杆菌蛋白酶的底物,如四肽底物、甲基苯磺酸-L-精氨酸甲酯(TEAM)、酪蛋白等,根据底物命名的方法对应为纤维块溶解时间法(CLT)、四肽底物法、TEAM法和酪蛋白法。
以纤维蛋白为底物的CLT法能粗略测定NK酶活,是纤溶酶、尿激酶等抗血栓酶常用的酶活测定方法。在37 oC恒温水浴环境下,利用凝血酶与一定量纤维蛋白原生成纤维蛋白,从加入酶后体系产生气泡起,到无气泡冒出作为样品反应时间。先测定已知活力的不同尿激酶对应的溶解时间,得到酶活-溶解时间标准曲线,再测定酶样品的溶解时间,带入标曲中得出酶活。此法方便快捷,但是精度较低,同时无法同时测定多个样品。
四肽底物法源于与NK同源性达99.5%的枯草杆菌蛋白酶E的活力测定方法,NK能将人工合成的小分子四肽Suc—Ala—Ala—Pro—Phe—pNA水解生成显色物质对硝基苯胺(p-NA),在410nm出测定吸光度,根据p-NA生成的量得出NK活力。该方法简便迅捷,精度较高,但不能完全反映纤溶活性。
TEAM可被NK切割生成甲醇,甲醇被高锰酸钾氧化为甲醛,后能与变色酸在沸水中发生显色反应,产物在574nm处有强吸收,根据吸光度和反应关系从而得出NK样品酶活。此法操作简便、灵敏度高,但易受样品中杂质的影响,另外其酶活结果不能完全代表纤溶活性。
酪蛋白法来源于经典的枯草杆菌蛋白酶活力测定方法,NK与酪蛋白40 oC水浴生成酪氨酸,酪氨酸与福林酚试剂30 oC下能发生显色反应,产物在680nm处有强烈吸收,以已知不同濃度的酪氨酸与福林酚反应,得到浓度吸光度标准曲线,再由酶活定义得出酶活-吸光度标准曲线,由最终吸光度即可得出NK样品酶活。该法简便易行、灵敏度高、可同时测定多个样品等,同时有研究表明此法得出的酶活结果与纤溶活性呈现一定的线性关系,故此法有望成为新的NK活力检测方法。但由于NK特殊性,此法实际应用于NK时也存在一些问题,如酶样品中杂质的影响等。
5.3 酶联免疫吸附法(ELISA)
根据NK的抗原性分别制备其单克隆和多克隆抗体,先将NK与单克隆抗体结合,将多克隆抗体与过氧化物连接,再将NK结合体与多克隆复合体结合形成类似三明治的结构,过氧化物酶能与底物发生反应,生成显色产物,在490nm出有强吸收,有吸光度即可确定NK样品的活性。此法灵敏度极高,但操作极为繁琐,价格昂贵,自报道以来未见后续研究。
6 结论与展望
到目前为止,纳豆激酶的抗血栓效果已经得到了初步的验证,在小鼠、犬、兔等模型中观察到了明显的溶栓效果,在人体临床实验中也得到了积极的从侧面反映的抗血栓疗效,但缺乏更直观的临床实验结果,鉴于血栓疾病的长期性,外界环境因素的复杂性,这可能是极具挑战性的问题。NK结构特征和作用机理已经基本明晰,基因工程研究正尝试使NK在植物或者其他机体中直接表达出来,同时优化改进其基因以期得到更强更适应于人体的抗血栓产品。NK具体活力的测定已经有较多依据,市面上的相关产品也十分丰富,NK正越来越为人们熟知,有望为更多血栓疾病患者带来希望。
References:
[1]Chen H,et al. Nattokinase:A Promising Alternative in Prevention and Treatment of Cardiovascular Diseases. Biomark Insights,2018. 13:p. 1177271918785130.
[2]杨勤军,罗梦曦与李佩佩,中风后遗症中医分型及治疗研究进展. 江西中医药大学学报,2016. 28(01):第113-117页.
[3]尹咪咪,心电信号分析处理及心肌梗塞疾病模型的建立,2016,郑州大学. 第 79页.
[4]胡盛寿等,《中国心血管病报告2018》概要. 中国循环杂志,2019. 34(03):第209-220页.
[5]邵荣军,溶栓酶活性检测方法的比较研究及高效溶栓菌株的筛选,2012,重庆师范大学. 第 45页.
[6]Wu H,et al. Acute toxicity and genotoxicity evaluations of Nattokinase,a promising agent for cardiovascular diseases prevention. Regul Toxicol Pharmacol,2019. 103:p. 205-209.
[7]杨亚楠等,纳豆激酶稳定性研究. 辽宁中医药大学学报,2018. 20(11):第41-44页.
[8]Sumi H,et al. A novel fibrinolytic enzyme(nattokinase)in the vegetable cheese Natto;a typical and popular soybean food in the Japanese diet. Experientia,1987. 43(10):p. 1110-1.
[9]Wu H,et al. Nattokinase-heparin exhibits beneficial efficacy and safety-an optimal strategy for CKD patients on hemodialysis. Glycoconj J,2019. 36(2):p. 93-101.
[10]Guo H,et al. Comparative anti-thrombotic activity and haemorrhagic adverse effect of nattokinase and tissue-type plasminogen activator. Food Sci Biotechnol,2019. 28(5):p. 1535-1542.
[11]Han L,et al. Development of a novel strategy for robust synthetic bacterial promoters based on a stepwise evolution targeting the spacer region of the core promoter in Bacillus subtilis. Microbial Cell Factories,2019. 18(1).
[12]张杰等,纳豆激酶高产菌株的选育及固态发酵技术. 食品科学,2016. 37(03):第151-156页.
[13]高泽鑫等,纳豆激酶的研究进展与展望. 中国酿造,2017. 36(08):第11-15页.
[14]王天琦,纳豆激酶体内药效和毒性研究,2018,辽宁大学. 第 66页.
[15]张锋,金杰与解成骏,纳豆激酶最新研究进展. 中国调味品,2008(01):第29-34页.
[16]Fujita M,et al. Transport of Nattokinase across the Rat Intestinal Tract. Biological and Pharmaceutical Bulletin,1995(9).
[17]Y S,et al. Dietary supplementation with fermented soybeans suppresses intimal thickening. Nutrition,2003. 19(3):p. 261-264.
[18]舒敏,纳豆激酶在毕赤酵母中的表达及在大鼠体内溶栓作用的研究,2016,湖北大学. 第 67页.
[19]刘瑞娟,张叶与帖卫芳,纳豆激酶的研究进展. 黑龙江科学,2019. 10(02):第40-41页.
[20]法芸等,纳豆激酶分离纯化和酶活性测定的研究进展. 色谱,2019. 37(03):第274-278页.
[21]杨亚楠等,纳豆激酶稳定性研究. 辽宁中医药大学学报,2018. 20(11):第41-44页.
[22]Ju S,et al. Isolation and Optimal Fermentation Condition of the Bacillus subtilis Subsp. natto Strain WTC016 for Nattokinase Production. Fermentation,2019. 5(4):p. 92.
[23]Cui,W,et al. Stepwise modifications of genetic parts reinforce the secretory production of nattokinase in Bacillus subtilis. Microb Biotechnol,2018. 11(5):p. 930-942.
[24][2019]纳豆芽孢杆菌发酵产纳豆激酶条件优化及酶品质分析_葛芸.
[25][2018]纳豆激酶热稳定性及pH稳定性的改造研究_赵菡.
[26]Yanagisawa,Y,et al. Purification,crystallization and preliminary X-ray diffraction experiment of nattokinase from Bacillus subtilis natto. Acta Crystallogr Sect F Struct Biol Cryst Commun,2010. 66(Pt 12):p. 1670-3.
[27]NguyenT T,T D,Quyen and Le HT. Cloning and enhancing production of a detergent- and organic-solvent-resistant nattokinase from Bacillus subtilis VTCC-DVN-12-01 by using an eight-protease-gene-deficient Bacillus subtilis WB800. Microb Cell Fact,2013. 12:p. 79.
[28][2019]纳豆激酶体内药效和毒性研究_王天琦.
[29]牟小超,纳豆激酶活性和稳定性的化学研究,2017,山西大学. 第 82页.
[30]Chen H,et al. Nattokinase:A Promising Alternative in Prevention and Treatment of Cardiovascular Diseases. Biomarker Insights,2018. 13:p. 117727191878513.
[31]Weng Y,et al. Nattokinase:An Oral Antithrombotic Agent for the Prevention of Cardiovascular Disease. International Journal of Molecular Sciences,2017. 18(3):p. 523.
[32]Kumar S S and A Sabu. Fibrinolytic Enzymes for Thrombolytic Therapy. Adv Exp Med Biol,2019. 1148:p. 345-381.
[33]张莉,纳豆加工工艺的研究和产品开发,2012,山东轻工業学院. 第 72页.
[34][2016]纳豆激酶产品研究现状及其(测定)进展_梁一博.
[35]Yuki Y,et al. A Sandwich Enzyme-linked Immunosorbent Assay for Nattokinase. Bioscience,Biotechnology,and Biochemistry,1994. 58(2):p. 366-370.
[36]《食品与发酵工业》网络首发论文.
[37]谢平会等,纤维蛋白平板法测定保健食品中纳豆激酶活力. 中国卫生检验杂志,2010. 20(07):第1658-1659页.
[38]赵明等,薄层聚丙烯酰胺纤维蛋白平板法对纤溶酶活性的体外检测. 四川师范大学学报(自然科学版),2005(02):第228-230页.
[39]法芸等,纳豆激酶分离纯化和酶活性测定的研究进展. 色谱,2019. 37(03):第274-278页.
[40]邵荣军,溶栓酶活性检测方法的比较研究及高效溶栓菌株的筛选,2012,重庆师范大学. 第 45页.
[41][GB]蛋白酶制剂.
[42][2015]基于分光光度法构建纳豆激酶纤溶活性快速测定方法(2).
[43]张俊杰等,双菌株联合发酵提高鹰嘴豆纳豆激酶活性的研究. 轻工学报,2019:第1-10页.
[44][2011]基因工程受体表达细节-纳豆激酶研究进展_敬俊锋.
[45]Han L,et al. Development of a novel strategy for robust synthetic bacterial promoters based on a stepwise evolution targeting the spacer region of the core promoter in Bacillus subtilis. Microbial Cell Factories,2019. 18(1).
猜你喜欢
智富时代(2019年5期)2019-07-05
智富时代(2019年5期)2019-07-05
进出口经理人(2018年1期)2018-01-24
中学生数理化·高一版(2017年1期)2017-04-25
初中生世界·七年级(2017年2期)2017-01-20
中学化学(2016年10期)2017-01-07
新高考·高三数学(2016年4期)2016-08-10
科技知识动漫(2016年5期)2016-06-04
作文大王·低年级(2016年3期)2016-03-11
中学数学杂志(高中版)(2016年1期)2016-02-23
