
贵州羊肚菌的分子系统发育分析
2020-08-25 10:02李银双刘作易谭玉梅任锡毅刘永翔黄永会
山地农业生物学报 2020年3期
关键词:贵州
李银双 刘作易 谭玉梅 任锡毅 刘永翔 黄永会

摘 要:本研究以贵州省贵阳市、六盘水、遵义和铜仁等地采集的羊肚菌样本中挑选出24份子实体样本,采用五个基因片段(ITS、LSU、EF1-a、RPB1、RPB2)联合分析,结合子实体形态特征对其进行了物种鉴定。研究结果表明:贵州省境内共分布有8个野生羊肚菌物种,包括黑色羊肚菌和黄色羊肚菌两个类群,黑色羊肚菌有梯棱羊肚菌(M. importuna)、Mel-19、Mel-21,黄色羊肚菌有Mes-15、Mes-19、Mes-22、Mes-23、Mes-27。野生羊肚菌的发生时间为3—5月,具有喜湿喜阴的生长特性,Mes-15是首次报道具有火烧地偏好性的黄色种。
关键词:贵州;野生羊肚菌;系统发育;多基因联合
中图分类号:Q-9文献标识码:A
文章编号:1008-0457(2020)03-0052-09 国际DOI编码:10.15958/j.cnki.sdnyswxb.2020.03.009
Molecular Phylogenetic Analysis of Morchella in Guizhou
LI Yinshuang.1,3 , LIU Zuoyi.4* , TAN Yumei.2,3 , REN Xiyi.2,3 , LIU Yongxiang.2,3 ,HUANG Yonghui.2,3
(1. College of Life Sciences, Guizhou University, Guiyang, Guizhou 550025, China; 2. Institute of Biotechnology, Guizhou Academy of Agricultural Sciences, Guiyang, Guizhou 550006, China; 3. Guizhou Key Laboratory of Agricultural Biotechnology, Guiyang, Guizhou 550006, China;4. Guizhou Academy of Agricultural Sciences, Guiyang, Guizhou 550006, China)
Abstract:Combined with morphological characteristics of fruiting body of Morchella, 24 samples collected from Guiyang, Liupanshui, Zunyi and Tongren regions , Guizhou Province were identified using five gene fragments (ITS, LSU, EF1-a, RPB1, RPB2) . The results showed that there existed 8 wild Morchella species in Guizhou Province, including two groups of black and yellow Morchella. Black morel are M. importuna, Mel-19, Mel-21, and yellow one containing Mes-15, Mes-19, Mes-22, Mes-23, Mes-27. The occurrence of wild morels in Guizhou was from March to May and the species prefered moisture and shade. Mes-15 was first reported to prefer to growing in burnt land.
Keywords:Guizhou; wild morel; phylogeny; multigene combination
羊肚菌(Morchella)隸属于子囊菌亚门(Ascomycotina)、盘菌纲(Discomycetes)、盘菌目(Ezizales)、羊肚菌科(Morchellaceae)[1]。作为一种世界公认的珍稀食药用菌,羊肚菌不仅味道鲜美,而且对降血脂、调节机体免疫力、抗疲劳、抗菌、抗病毒、抑制肿瘤、减轻放化疗引起的毒副作用等具有显著功效[2-3]。在羊肚菌的分类研究早期,有学者依据羊肚菌子实体形状、大小、颜色、脉棱分布和凹坑深浅等形态特征将羊肚菌属划分为四大类群(黑色羊肚菌类群、黄色羊肚菌类群、半开羊肚菌类群和变红羊肚菌类群)[4-6]。然而,世界范围内羊肚菌属的形态学分类一直存在较大争议[7-9]。随着分子生物学技术在羊肚菌属分类研究中的引入,特别是多基因谱系一致性系统发育学物种识别法(genealogical concordance phylogenetic species recognition,GCPSR)的应用,为羊肚菌属的分类研究带来了革命性的变革[8,10]。2011年,ODONNELL等[11]首次利用4个基因(LSU、EF1-a、RPB1、RPB2)联合的方法对羊肚菌属进行了系统发育分析,认为羊肚菌属是由黑色羊肚菌支系(Elata Clade)、黄色羊肚菌支系(Esculenta Clade)、变红羊肚菌支系(Rufobrunnea Clade)三个支系构成,将基于形态学鉴定的“半开羊肚菌”划分到黑色羊肚菌支系中,提出采用所在类群缩写+数字的方法来进行羊肚菌属内物种命名(Esculenta支系:Mes-n;Elata支系:Mel-n),该研究很好的规避了羊肚菌分类过程中的主观性影响,解决了羊肚菌属分类研究中的诸多争议。已有研究证明,羊肚菌属内一共包括61个系统发育物种[12]。目前,部分黑色支系羊肚菌物种在我国已经成功实现了人工栽培,但生产中使用的栽培菌种主要来自野生子实体的人工分离驯化,因此,分离菌株的准确鉴定已成为羊肚菌栽培成功与否的关键[8,13-14]。
贵州温润多雨的气候条件孕育了丰富的野生羊肚菌资源。然而,有关贵州野生羊肚菌资源的种类、分布、生境等调查研究相对较少。在《贵州大型真菌资源》一书中记录有两种羊肚菌,分别为尖顶羊肚菌(Morchella conica)和羊肚菌(Morchella esculenta)[15] 。汪建文等[16]基于对现有资料整理与调查认为贵州分布有6个羊肚菌物种,分别为黑脉羊肚菌(Morchella angusticeps)、尖顶羊肚菌(Morchella conica)、粗柄羊肚菌(Morchella crassipes)、羊肚菌(Morchella eseulenta)、高羊肚菌(Morchella elata)、普通羊肚菌(Morchella vulgris)。但是,无论2种或者6种,这些调查研究仍停留在形态学水平,没有采用分子系统学的方法对其进行物种鉴定,并且对贵州地区野生羊肚菌的地域分布、发生时间和生活环境等也没有进行过深入研究。基于以上分析,本研究在2018—2019年期间对贵阳市、六盘水、遵义和铜仁等地区开展野生羊肚菌采集工作,系统调查了贵州当地野生羊肚菌的发生时间与物种分布地域情况,同时,采用五个基因(ITS、LSU、EF1-a、RPB1、RPB2)联合分析的方法对采集样本进行了种类鉴定和物种多样性分析,进而为野生羊肚菌的栽培驯化、杂交育种与长期保护利用提供科学依据。
1 材料与方法
1.1 样本及培养基
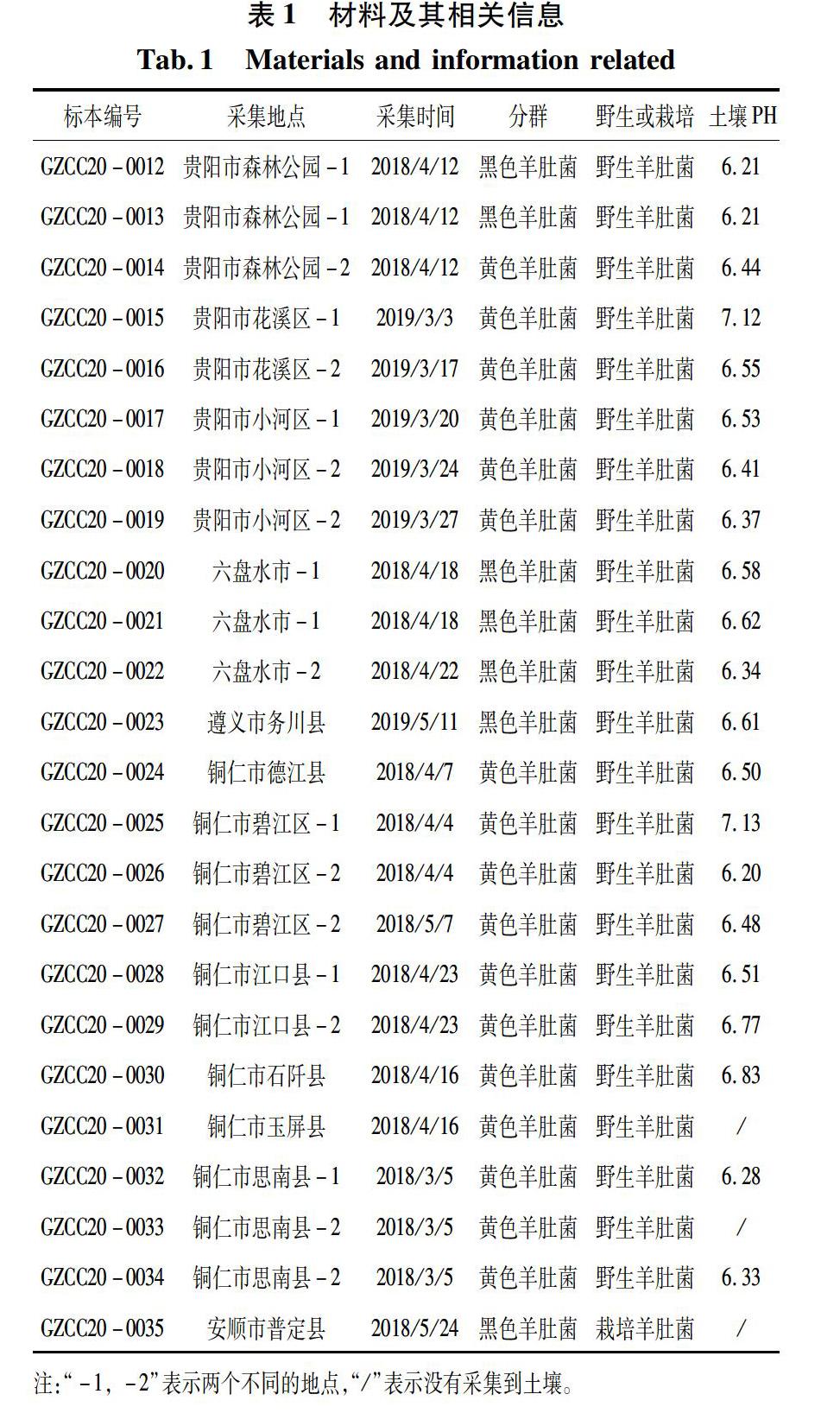
样本来源:2018—2019年期间,在贵州境内的贵阳、六盘水、遵义和德江等地共采集到78份野生羊肚菌子实体样本,在安顺市普定县羊肚菌栽培基地采集到2份栽培羊肚菌子实体样本。所有样本带回实验室后,自然风干-20℃保存于贵州省菌种中心。从采集样本中挑选出24份具有典型形态特征、采集时间、地域差异和涉及每个采集地点的子实体样本,编号GZCC20-0012~ GZCC20-0035,用于分子系统发育分析。24份样本的采集信息见表1。参照图书《中国大型真菌彩色图谱》[17],初步将24份采集样本分为黑色羊肚菌和黄色羊肚菌两个类群。
1.3.4 构建多基因系统发育树
参照文献[9]对所有样本序列和参考序列作拼接处理,将样本序列和参考物种的ITS、LSU、EF1-a、RPB1、RPB2单个基因序列集合分开在Bioedit Sequence Alignment Editor(Version5.09)软件中进行Clustalw Multiple Alignment处理,所有参数均为系统默认值;删除序列两端非对位的排列序列,数据中的空位“gap”作为缺失数据“missing date”处理,保证样本和参考物种每个基因的长度一致,所得结果保存为fas格式,命名为fas1~fas5。得到ITS、LSU、EF1-a、RPB1、RPB2单个序列集合的切除拼接结果后,在Edit模式下利用Bioedit sequence Alignment Editor(Version5.09)软件将五个序列集合以“ITS+LSU+EF1-a+RPB1+RPB2”的顺序拼接起来,所得结果依然保存为fas格式,命名为fas6。五个序列集合整理过程中,同时下载基部类群变红羊肚菌(Morchella rufobrunnea)的基因序列,将其作为系统发育分析的外类群。最后,把fas6文件输入系统发育软件MEGA6.0中,选择ML法(Maximum Likehood),Bootstrap重复1000次,其他数值均为系统默认值,构建系统发育树。由系统发育结果进行物种鉴定,分析贵州野生羊肚菌的物种多样性。
1.4 形态学描述及生境分析
依据每个采集地点抽取样本的多基因分子鉴定结果,结合鉴定菌株形态特征与每份样本的采集信息,详细记录每个羊肚菌物种子实体的菌盖和菌柄大小范围、颜色深浅、楞脉分布等形态特征,记录每个物种羊肚菌子实体的生长分布方式、生长环境、分布区域等,分析贵州当地野生羊肚菌的发生时间及规律。
2 结果与分析
2.1 多基因系统发育鉴定
2.1.1 ITS、LSU、EF1-a、RPB1和RPB2扩增测序
24份样本的ITS扩增结果中,7份黑色羊肚菌样本(GZCC20-0012、GZCC20-0013、GZCC20-0020、GZCC20-0021、GZCC20-0022、GZCC20-0023、GZCC20-0035)的ITS片段长度在750 bp左右,17份黄色羊肚菌样本(GZCC20-0014、GZCC20-0015、GZCC20-0016、GZCC20-0017、GZCC20-0018、GZCC20-0019、GZCC20-0024、GZCC20-0025、GZCC20-0026、GZCC20-0027、GZCC20-0028、GZCC20-0029、GZCC20-0030、GZCC20-0031、GZCC20-0032、GZCC20-0033、GZCC20-0034)的ITS片段長度在1100bp左右,两个类群的ITS片段长度存在较大差别。其LSU片段在1100bp左右,EF1-a片段大小在1100bp左右,RPB1片段大小在750bp左右,RPB2片段在800bp左右,该四个片段大小在两个羊肚菌类群中则无明显差异。所有扩增结果送至上海生物工程有限公司进行测序。
2.1.2 ITS、LSU、EF1-a、RPB1、RPB2序列比对分析
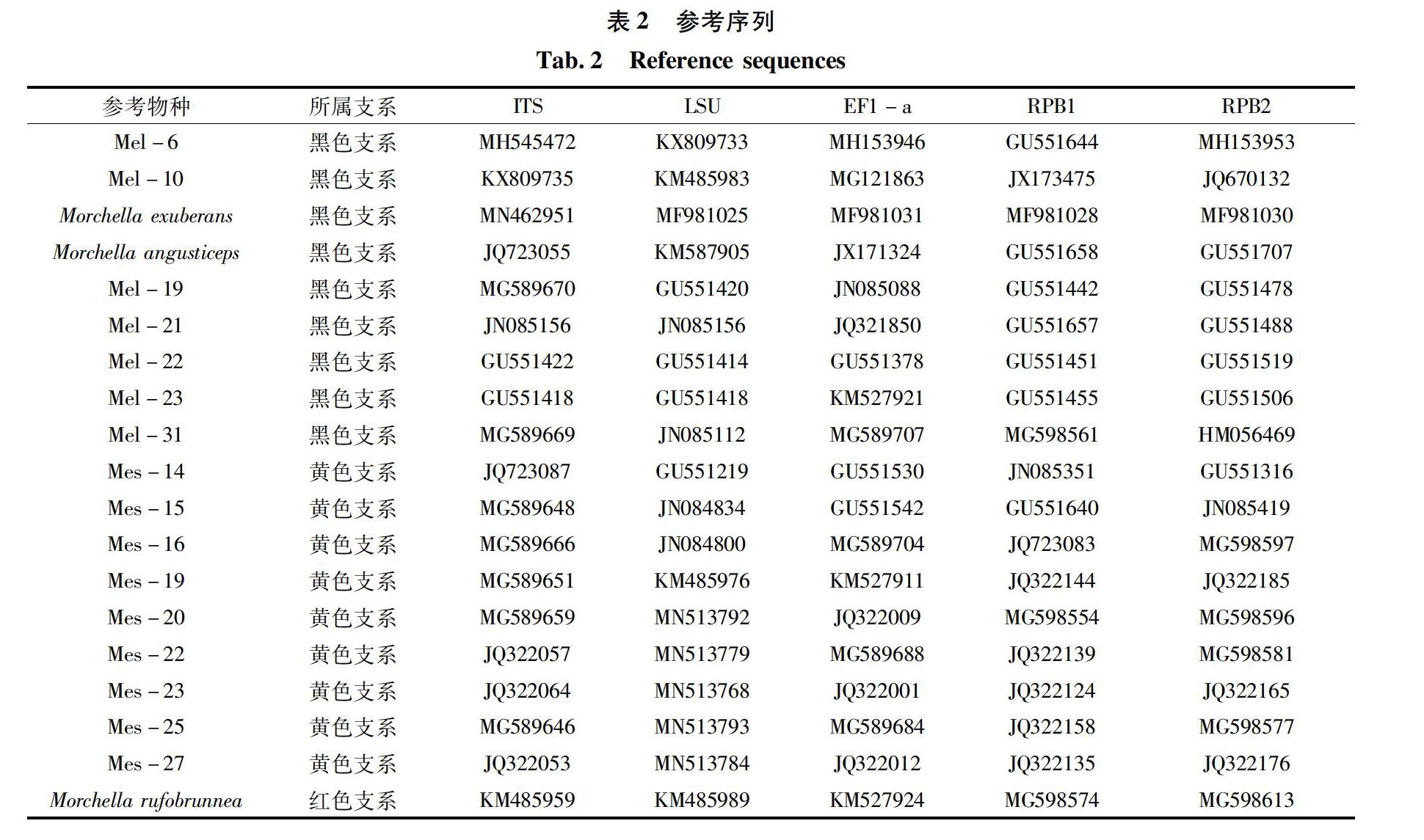
获得所有序列信息后,切除引物片段输入NCBI blast比对分析,下载并筛选出相似性较高的羊肚菌序列作为参考序列(表2)。120条样本序列整理上传至NCBI数据库后,得到每个基因的序列登录号如表3。
2.1.3 构建多基因系统发育树
由于黄色羊肚菌类群的ITS片段较长,为保证序列中的关键分析位点不被切除,故均以变红羊肚菌(M. rufobrunnea)作为外类群,将黑色羊肚菌和黄色羊肚菌两个类群分开分析。
黑色羊肚菌类群中,所有样本序列及参考序列经过对齐处理后,ITS序列的保留长度为580bp,LSU序列的保留长度为650bp,EF1-a序列的保留长度为760bp,RPB1序列的保留长度为610bp,RPB2序列的保留长度为600bp。将所有序列按照“ITS+LSU+EF1-a+RPB1+RPB2”的顺序比对拼接后,全序列长度为3200bp,采用MEGA6.0构建系统发育树(图1)。以变红羊肚菌(Morchella rufobrunnea)作为外类群(图1),构建黑色羊肚菌类群的系统发育树后,7份黑色羊肚菌样本序列与其他参考序列被聚类在一个大的分支上。其中,GZCC20-0012、GZCC20-0013、GZCC20-0022与Mel-21以99%的Bootstrap值聚为一类;GZCC20-0023与Mel-19以100%的Bootstrap值聚为一类;GZCC20-0035与Mel-6以100%的Bootstrap值聚为一类;GZCC20-0020、GZCC20-0021与Mel-10以100%的Bootstrap值聚为一类。而梯棱羊肚菌(M. importuna)、六妹羊肚菌(M. sextelata)对应的系统发育物种分别是Mel-10和Mel-6。由此可知,黑色羊肚菌类群由Mel-21、Mel-19、梯棱羊肚菌(M. importuna)、六妹羊肚菌(M. sextelata)四個物种构成。其中,鉴定为六妹羊肚菌(M. sextelata)的GZCC20-0035为栽培样本。
黄色羊肚菌类群中,所有样本序列及参考序列经过对齐处理后,ITS序列的保留长度为860bp,LSU序列为550bp,EF1-a序列为800bp,RPB1序列为600bp,RPB2序列为655bp。将所有序列按照“ITS+LSU+EF1-a+RPB1+RPB2”的顺序拼接后,全序列长度为3465bp,采用MEGA6.0构建系统发育树(图2)。以变红羊肚菌(Morchella rufobrunnea)作为外类群(图2),构建黄色羊肚菌类群的系统发育树后,17份黄色羊肚菌样本序列与其他参考序列被聚类在一个大的分支上。其中,GZCC20-0014、GZCC20-0018、GZCC20-0017、GZCC20-0016、GZCC20-0015、GZCC20-0019、GZCC20-0025、GZCC20-0029与Mes-15以98%的Bootstrap值聚为一类,GZCC20-0015和GZCC20-0016亲缘性较近;GZCC20-0026、GZCC20-0027、GZCC20-0028、GZCC20-0032、GZCC20-0034与Mes-19以100%的Bootstrap值聚为一类,GZCC20-0028、GZCC20-0032、GZCC20-0034亲缘性较近;GZCC20-0030与Mes-23以100%的Bootstrap值聚为一类;GZCC20-0031、GZCC20-0033与Mes-22以100%的Bootstrap值聚为一类;GZCC20-0024与Mes-27以100%的Bootstrap值聚为一类。由此可知,黄色羊肚菌类群由Mes-15、Mes-19、Mes-22、Mes-23、Mes-27五个物种构成。
2.2 形态学描述及生境分析
2.2.1 形态学描述
依据多基因分子鉴定结果,结合每份样本的采集信息,将9个羊肚菌物种的形态特征,生长分布等描述如下:
Mel-21:属黑色羊肚菌支系,单生或双生。采集时间为4月份,分布于贵阳市森林公园,六盘水市。基部土壤pH6.14~6.57,子实体周围环境潮湿,植被类型为阔叶林。菌盖圆锥形,黑色,长4.8cm~6.9cm,宽3.1cm~4.7cm,顶端或尖或钝。菌柄长2.9cm~5.1cm,粗2.1cm~3.7cm,污白色,大小均匀(图3:GZCC20-0012)。
Mel-19:属黑色羊肚菌支系,单生。采集时间为5月份,分布于遵义市务川县。基部土壤pH=6.51~6.77,子实体周围环境潮湿,分布苔藓和草类植物,植被类型为阔叶林与针叶林混交林。子实体粗壮,菌盖圆锥形,褐色、黑褐色,长5.0cm~6.7cm,宽3.7cm~4.8cm,表面表面棱脊曲折,凹坑呈不规则形状,密集形似蜂窝。菌柄长6.3cm~6.7cm,粗3.3cm~3.7cm,灰白色、灰黄色,表面平滑,与菌盖连接处较细,基部略膨大(图3:GZCC20-0023)。
梯棱羊肚菌(Mel-10):属黑色羊肚菌支系,单生。采集时间4月份,分布于六盘水市。基部土壤pH=6.51~7.22,子实体周围环境潮湿、分布苔藓和蕨类植物,植被类型为阔叶林。菌盖圆锥形,灰色或灰褐色,长5.3cm~8.7cm,宽3.9cm~6.0cm,顶端尖,表面凹坑内部有横隔。菌柄长2.9cm~5.2cm,粗2.5~4.0cm,污白色;表面分布有分布有浅纵沟,伴有少量微绒毛和颗粒物(图3:GZCC20-0020)。
六妹羊肚菌(Mel-6):栽培羊肚菌,属黑色羊肚菌支系,单生、双生或丛生。子实体较大,菌盖长圆锥形或长圆柱形,长6.5cm~8.2cm,宽4.4cm~6.3cm,颜色褐色、红褐色、朱砂红。菌柄长5.2cm~7.2cm,粗2.7cm~5.5cm,白色,表面分布有大量纵沟与颗粒物质,基部膨大。该种可从子实体大小和颜色上和其他物种明显区分开(图3:GZCC20-0035)。
Mes-15:属黄色羊肚菌支系,单生、双生或丛生。采集时间3月份、4月份,该种分布较广,分布于贵阳市森林公园、贵阳市花溪区、贵阳市小河区、铜仁市碧江区、铜仁市江口县。基部土壤pH=6.27~7.36。子实体周围环境潮湿、分布苔藓和草类植物,采集于阔叶林、绿化草坪。在该物种的两个采集地出现了大量出菇现象,周围伴有火烧地痕迹,推测该物具有火烧地偏好性。菌盖卵圆形、圆锥形或无规则形状,颜色黄棕色、淡黄色,长3.0cm~5.5cm,宽2.7cm~4.8cm,顶端或尖或钝,表面凹坑较深,凹坑内部有横隔。外部脉棱上分布有少量刺突,肉质特别脆,这是该种的一个显著标志。菌柄长2.3cm~6.4cm,粗1.7cm~2.9cm,颜色淡黄色,表面光滑,大小均匀(图3:GZCC20-0014)。
Mes-19:属黄色羊肚菌支系,单生。采集时间3月、4月、5月,分布于铜仁市碧江区,铜仁市江口县,铜仁市思南县。基部土壤pH=6.20~7.18,实体周围环境潮湿、分布苔藓和草类植物,采集于阔叶林、绿化草坪。菌盖近圆柱形或圆球形,形似花朵,颜色浅灰色、灰黄色,长4.0cm~5.9cm,宽3.8cm~4.2cm,表面凹坑较深,顶端圆钝。菌柄长4.5cm~5.7cm,粗2.2cm~3.5cm,颜色淡黄色,表明光滑(图3:GZCC20-0027)。
Mes-22:属黄色羊肚菌支系,单生。采集时间3月份、4月份,分布于铜仁市玉屏县,铜仁市思南县。未采集到土壤,子实体周围潮湿,分布有大量草类植物,采集于河滩草丛中。菌盖长圆锥形,颜色淡黄色、灰白色,长4.8cm~6.7cm,宽3.8cm~4.1cm,顶端或尖或钝,肉质脆,部分棱脊上分布有小裂痕。菌柄长3.7cm~5.1cm,粗1.8cm~2.9cm,颜色白色,表面光滑(图3:GZCC20-0031)。
Mes-23:属黄色羊肚菌支系,单生,双生。采集时间4月份,分布于铜仁市石阡县,基部土壤pH=6.45~7.27,子实体周围环境潮湿、分布苔藓和草类植物,植被类型为阔叶林。菌盖圆柱形、圆锥形、圆球形皆有,长4.8cm~5.7cm,宽4.2cm~4.6cm;表面凹坑较深,肉质脆,幼小时深褐色,成熟后颜色黄灰色、黄褐色;菌肉表面有少许刺突。菌柄长4.3cm~5.9cm,粗1.9cm~2.5cm,污白色、灰黄色,部分表面分布有少量颗粒物质(图3:GZCC20-0030)。
Mes-27:属黄色羊肚菌支系,单生,双生或丛生。采集时间4月份,分布于铜仁市德江县。基部土壤pH=6.50~7.51,子实体周围潮湿,分布有苔藓,植被类型为阔叶林与针叶林混交林。菌盖近卵圆锥形,表面凹坑较深,顶端或尖或钝,长4.2cm~5.7cm,宽3.6cm~4.4cm,颜色灰色、黄灰色。菌柄长4.2cm~5.0cm,粗1.9cm~2.6cm,白色、污白色,表面光滑,大小均匀(图3:GZCC20-0024)。
2.2.2 生境分析
从多基因系统发育结果与样本采集信息可知,不同羊肚菌物种在形态特征、生长分布方式和分布地域等存在较大差异。在生境上,阔叶林、混交林地、绿化草坪和河滩湿地均能采集到羊肚菌子实体,其周围一般都有苔藓或草类等禾本科植物,环境相对阴湿,表明羊肚菌具有喜湿喜阴的生长特性。测定基部土壤pH表明,羊肚菌的生长偏中性,两个类群无明显差别。由样本采集信息(表1)及其他人的经验,只在3月、4月和5月份里采集到了羊肚菌子实体,而每年的3—5月份为贵州的无霜期,大部分地区气温回升、降雨增加,推测该时间段为贵州地区野生羊肚菌子实体高发生期。值得一提的是,已有研究表明部分羊肚菌物种具有火烧地偏好性,具体表现为:在发生过火烧的林地或耕地的第一年会出现羊肚菌大量出菇的现象[11,19]。迄今為止,共发现6个(Mel-1、Mel-6、Mel-7、Mel-8、Mel-9、Mel-10)偏好火烧迹地生境物种,全为黑色支系物种[8,11]。本研究在贵阳市的两个采集地发现:黄色支系物种Mes-15子实体大量出现在上一年的火烧林地中,火烧地特征与其他报道相符[8,11,19],表明Mes-15这一物种也具有火烧地偏好性。
3 结论与讨论
羊肚菌的形态学分类体系一直存在较大争议,分子生物学分析技术在羊肚菌属分类研究中的引入,解决了羊肚菌属分类研究中的诸多争议。然而,目前羊肚菌属内物种是根据多基因谱系进行界定的,各系统发育物种的形态学研究还不够深入,形态学记录相当有限,这给羊肚菌商品信息和科学信息的准确交流带来了很多麻烦[8,20]。本研究通过对采集样本进行形态学初步分析也发现,羊肚菌属内各系统发育物种的形态学记录非常之少,只能将采集样本初步分为黑色羊肚菌和黄色羊肚菌两个类群。
野生羊肚菌资源在我国许多省份都有分布,武冬梅[9]采用4个基因(ITS、EF1-a、RPB1、RPB2)联合的方法对我国新疆地区的野生羊肚菌分布物种进行分析,认为新疆地区共分布有7个羊肚菌物种,新增了两个中国新纪录种(Mel-2、Mes-17)。除上述报道外,国内关于我国野生羊肚菌资源分布的多基因谱系研究还不多。贵州良好的气候类型,孕育了丰富的野生羊肚菌资源。但是,以往贵州野生羊肚菌资源的调查研究,只停留在形态学水平。另外,野生羊肚菌出菇期短,分布数量有限,大量采摘导致野生资源逐渐消失,如何科学保护和利用这一野生真菌资源成为一个亟需解决的问题。本研究通过对贵州境内采集的24份羊肚菌样本进行多基因谱系分析,将所有样本鉴定为9个不同的羊肚菌物种。其中,GZCC20-0035为采自安顺市普定县的栽培样本,属于黑色羊肚菌类群中的六妹羊肚菌(M. sextelata),是目前国内主要栽培物种之一。其余23份野生羊肚菌样本鉴定为8个野生羊肚菌物种,包括黑色羊肚菌和黄色羊肚菌两个类群,其中,黑色羊肚菌有Mel-21、Mel-19、梯棱羊肚菌(M. importuna),黄色羊肚菌有Mes-15、Mes-19、Mes-22、Mes-23、Mes-27。在准确的分子鉴定的基础之上,本研究给予了每个物种正确的科学名称,纠正了以往研究中错误命名的现象,详细记录了不同物种的形态特征、分布地域、生长分布方式等。并通过样本采集信息分析,认为贵州当地野生羊肚菌的发生时间为3—5月份,生长土壤pH接近中性,具有喜湿喜阴的生长特性,首次发现Mes-15这一黄色支系物种具有火烧地偏好性。这不仅丰富了贵州真菌资源信息库,也为当地野生羊肚菌资源的长期保护利用提供了科学依据,有望下一步加快野生菌种资源的栽培驯化和良种选育工作,促进羊肚菌产业的健康长远发展。
参 考 文 献:
[1] 黄年来,林志彬,陈国良,等.中国食药用菌学[M].上海:上海科学技术文献出版社,2010:1370-1351.
[2] 徐永强,张明生,张丽霞.羊肚菌的生物学特性,营养价值及其栽培技术[J].种子,2006,25(7):97-99.
[3] 万春艳.羊肚菌的药用研究近况[J].中国林副特产,2008(4):91-92.
[4] BUNYARD B A, ROYSE N D J. A systematic assessment of Morchella using RFLP analysis of the 28s ribosomal RNA gene [J]. Mycologia,1994,86(6):762-772.
[5] WIPF D, MUNCH J C, BOTTON B. DNA phylogenies in morels: complete sequences of the internal transcribed spacer of genes coding for rDNA in Morchella esculent(yellow morel) and Morchella conia(black morel) [J]. Applied and environmental microbiology, 1996,62(9):3541-3543.
[6] GUZMAN G, TAPIA F. The known morels in mexico, a description of a new blushing species, Morchella rufobrunnea, and new data on M.guatemalensis [J]. Mycologia,1998,90(4):705-714.
[7] 刘伟,冯丽,张亚平.新疆羊肚菌rDNA的ITS序列分析[J].石河子大学学报(自然科学版),2009,27(5):584-587.
[8] 杜习慧,赵琪,杨祝良.羊肚菌的多样性,演化历史及栽培研究进展[J].菌物学报,2014,33(2):183-197.
[9] 武冬梅.新疆野生羊肚菌分子系统学和遗传多样性研究[D].北京:中国农业大学, 2015:37-56.
[10] TAYLOR J W, JACOBSON D J, KROKEN S, et al. Phylogenetic species recognition and species concepts in fungi [J]. Fungal genetics and biology, 2000,31(1):21-32.
[11] ODONNELL K, ROONEY A P, MILLS G L, et al. Phylogeny and historical biogeography of true morels (Morchella) reveals an early Cretaceous origin and high continental endemism and provincialism in the Holarctic [J]. Fungal genetics and biology,2011,48(3):252-265.
[12] DU X H, ZHAO Q, O'DONNELL K, et al. Multigene molecular phylogenetics reveals true morels (Morchella) are especially species-rich in China [J].Fungal genetics and biology, 2012,49(6):455-469.
[13] 趙琪,康平德,戚淑威,等.羊肚菌资源现状及可持续利用对策[J].西南农业学报,2010,23(1):266-269.
[14] 杨燕,田鸿,张小平,等.基于ITS和ISSR的羊肚菌种质资源遗传多样性分析[J].西南农业学报,2018,31(10):2004-2009.
[15] 吴兴亮.贵州大型真菌资源[M].贵阳:贵州科学出版社, 1989:20-21.
[16] 汪建文,洪江,邓春英.中国贵州药用真菌资源Ⅰ子囊菌门[J].贵州科学,2018,36(5):18-23.
[17] 袁明生,孙佩琼.中国大型真菌彩色图谱[M].成都:四川科学技术出版社,2013:47-49.
[18] DU X H, ZHAO Q, YANG Z L, et al. How well do ITS rDNA sequences differentiate species of true morels (Morchella) [J]. Mycologia, 2012,104(6):1351-1368.
[19] GREENE D F, MACDONALD S E, CUMMING S, et al. Seedbed variation from the interior through the edge of a large wildfire in alberta [J].Canadian journal of forest research, 2005,35(7),1640-1647.
[20] 杜习慧.黑色羊肚菌支系的物种资源,生殖模式和遗传多样性研究进展[J].菌物研究,2019,17(4):240-251.
猜你喜欢
参花(上)(2022年6期)2022-06-16
大众科学(2021年10期)2021-12-23
学苑创造·A版(2021年9期)2021-09-16
贵阳文史(2017年5期)2018-02-02
全国新书目(2014年7期)2014-09-19
贵阳文史(2009年5期)2009-12-21
当代贵州(2009年18期)2009-11-26
雕塑(1996年1期)1996-07-12
