
毛竹扩张对亚热带常绿阔叶林主要树种叶结构型性状的影响
2020-08-18 07:24钟雅琪钟全林李宝银徐朝斌程栋梁乐新贵郑文婷
生态学报 2020年14期
钟雅琪,钟全林,,*,李宝银,余 华,4,徐朝斌,程栋梁,,乐新贵,郑文婷,6
1 福建师范大学地理科学学院, 福州 350007
2 湿润亚热带山地生态国家重点实验室培育基地, 福州 350007
3 福建师范大学福建省植物生理生态重点实验室, 福州 350007
4 闽江学院海洋学院, 福州 350108
5 江西阳际峰国家自然保护区管理局, 贵溪 335400
6 福州大学环境与资源学院, 福州 350116
植物功能性状(plant function traits,PFT)是指影响植物个体生存、发育和繁殖所表现出的生理、形态和生活习性上的特性[1-2]。它是植物长期演化过程中对不同生活环境适应的结果[3],反映了植物对生存环境中各种资源的利用情况与适应性[4-6],是连接植物与环境的桥梁[7]。叶片是植物进行光合作用,合成光合产物以及获取养分的重要场所,它与环境接触面积最大、是植物对环境变化响应明显的敏感性器官[5]。在植物各器官功能性状中,叶功能性状(leaf functional traits,LFT)与植物的生存策略及资源利用能力等联系密切[8],能够直接反映植物适应环境所形成的生存对策[9- 11]。叶功能性状可以划分为两大类型,即叶结构型性状和叶功能型性状。其中前者是指植物叶片的生物化学结构特征,它在一定环境中保持相对稳定不变,并且能反映植物为获得最大程度的碳收入而采取的适应性策略[12];后者则是反映叶片生长代谢的指标,它会随着时间和空间的变化而改变,稳定性相对较差,难以深入解释植物在长期进化过程中的适应策略[13]。因此,本文仅从叶结构型性状方面开展亚热带常绿阔叶林主要树种对毛竹扩张的响应与适应机制研究,具有重要的生态学意义。
毛竹(Phyllostachysedulis)属于禾本科(Poaceae)竹亚科(Bambusoideae)刚竹属(Phyllostachys),是一种特殊的高大克隆乔木状散生植物。它依靠其鞭根进行无性繁殖,向临近的林分内生长并逐渐扩张、蔓延,具有强大的繁衍生殖和空间拓展能力[14]。亚热带常绿阔叶林是我国重要的地带性植被,具有丰富的生物多样性资源和巨大的生态效益[15],在调节气候、涵养水源和保持水土等方面发挥着重要作用[16]。由于毛竹在向临近常绿阔叶林扩张过程中,与阔叶树种的生态位存在着交叉或重叠[16],其必然会与阔叶树种争夺光、水、热、土壤养分以及地上空间等资源,可能导致林木叶片功能性状发生变化,进而影响林木的正常生长与生存。目前,有关毛竹扩张对亚热带常绿阔叶林影响的研究多集中于毛竹扩张对森林的物种组成[17]、群落结构[18]、生物多样性[16]、N循环[19]等群落学与生态效应影响方面,而有关毛竹扩张对常绿阔叶林林木叶功能性状影响方面的研究较少。本文以存在于常绿阔叶林和竹阔混交林中的四个相对优势树种,即常绿阔叶树种甜槠(Castanopsiseyrei)和青冈(Cyclobalanopsisglauca)、落叶阔叶树种青榨槭(Acerdavidii)和枳椇(Hoveniaacerba)为研究对象,试图探究:(1)毛竹扩张是否会改变乔木层林木叶结构型性状特征?(2)毛竹扩张对不同生活型林木叶结构型性状的影响是否具有一致性?(3)毛竹扩张是否改变乔木层4种林木间叶结构型性状的差异特征?(4)毛竹扩张是否会影响乔木层林木叶结构型性状指标间的关系特征?研究结果可为探明我国亚热带常绿阔叶林主要森林植物叶功能性状对毛竹扩张的响应机制,进一步开展毛竹扩张对常绿阔叶林的林分质量与生态功能的影响机制研究等提供理论依据,并为适度控制毛竹扩张、开展常绿阔叶林生态系统健康研究等提供实践指导。
1 材料与方法
1.1 研究区概况
江西阳际峰自然保护区地处江西省东北部,贵溪市境内(27°51′07″—28°00′24″N, 117°13′42″—117°24′22″E),属武夷山脉中段西北坡(图1),保护区面积10946 hm2,森林覆盖率高达99.7%,境内保存有6000余hm2原生性较强的常绿阔叶林,1878种高等植物,分别属8个植被型72个群系,240科830属,是我国亚热带常绿阔叶林的典型代表。保护区属中亚热带湿润季风山地气候,年平均气温11.4—18.5℃,年平均降水量1870—2191.3 mm,无霜期200—256 d,区内气象灾害少,光、温、水等气候资源时空分布较均匀,有利于动植物生长及繁衍,为科学研究提供了理想的研究平台。
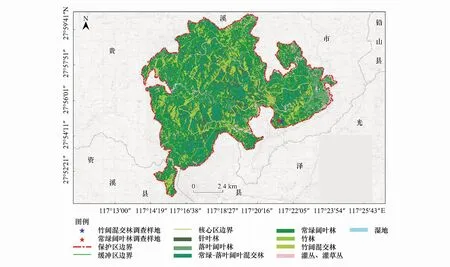
图1 研究区位置图
1.2 样地设置与样品采集
在江西省阳际峰国家级自然保护区内,沿毛竹水平扩张方向,分别设置6个常绿阔叶林样地与6个竹阔混交林样地,共12个样地。样地面积为15 m×15 m。所选取的样地海拔在700—1200 m,竹阔混交林样地受毛竹扩张影响达20余年。于生长季(2017年6月)对样地内毛竹及胸径5.0 cm以上的林木进行每木调查,并分树种计算其平均胸径、平均高及重要值。选取在常绿阔叶林与竹阔混交林乔木层中都有分布且具有一定代表性的两常绿阔叶树种(甜槠与青冈)与两落叶阔叶树种(青榨槭与枳椇)共4树种,对其叶结构型性状进行分析。4树种重要值分别为甜槠14.68%、青冈10.99%、青榨槭12.18%、枳椇3.16%,为阳际峰自然保护区常绿阔叶林中2生活型植物(常绿阔叶与落叶阔叶)的相对优势种。对所选取的4树种分别样地、分树种选择胸径、株高接近其平均胸径和平均高的健康木3株进行叶片采样。选取林冠中层完全展开的外层成熟健康叶片进行采样,所采叶片的叶龄均为当年生叶,分东、南、西、北四个方向选取1小枝,每个枝条选3—5片叶。将所采集叶片立即置于塑封袋中,编号带回实验室。样地基本情况见表1与表2。

表1 江西阳际峰国家级自然保护区不同林分样地概况
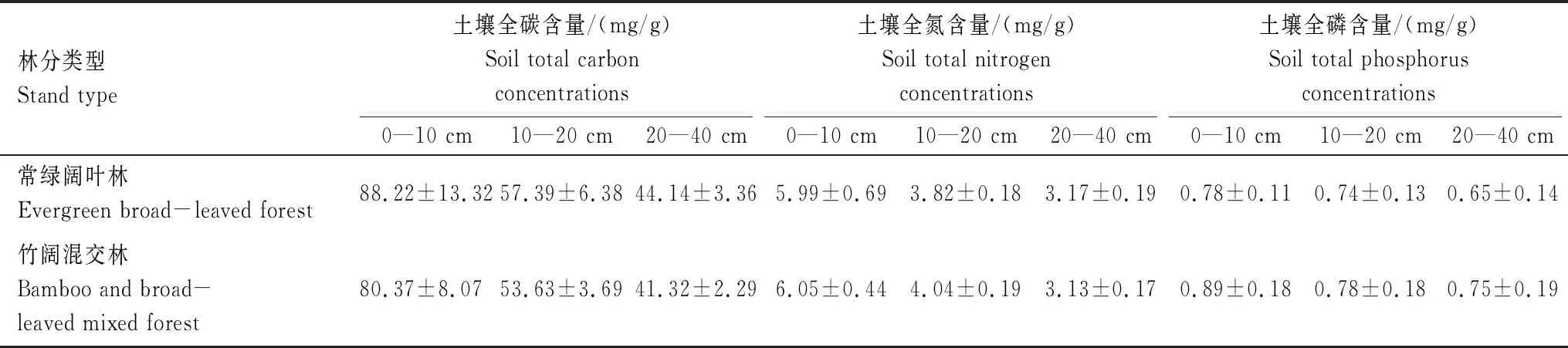
表2 江西阳际峰国家级自然保护区不同林分样地土壤养分含量
1.3 叶功能性状指标选取与测定
选取叶长(leaf length, LL)、叶宽(leaf width, LW)、叶面积(leaf area, LA)、比叶面积(specific leaf area, SLA)、叶厚(leaf thickness, LT)、叶鲜重(leaf fresh weight,LFW)、叶干重(leaf dry weight, LDW)、叶干物质含量(leaf dry matter content, LDMC)、叶组织密度(leaf tissue density, LTD)、叶相对含水量(leaf relative water content, LRWC)、叶形态指数(leaf morphology index, LMI)等11个叶结构型性状进行分析。叶结构型性状测定步骤如下:
利用EPSON V19扫描仪扫描叶片,用Image J软件计算LL、LW、LA;用游标卡尺(精度0.01 mm)测定叶片沿着主脉同一侧方向上、中、下3个位置上的厚度(避开主叶脉),取平均值即为该叶片LT;采用电子天平(精度0.01 g)称取LFW后,将叶片样品浸泡于清水中,于5℃的黑暗环境中浸12 h,取出后迅速用吸水纸去除叶片表面的水分,称其饱和鲜重;之后将其放在75℃下恒温箱内烘干48—72 h后称重,称LDW。计算其LRWC、LDMC、SLA、LMI及LTD等叶性状指标。计算公式如下:
SLA(cm2/g)=叶面积(cm2)/叶干重(g)
LDMC(g/kg)=叶干重(g)/叶饱和鲜重(g)
LRWC(%)=[叶鲜重(g)-叶干重(g)]/[叶饱和鲜重(g)-叶干重(g)]×100%
LTD(g/cm3)=叶干重(g)/[叶面积(cm2)·叶厚(mm)]
LMI(cm/cm)=叶长(cm)/叶宽(cm)
1.4 数据处理
利用Canoco 5软件中的主成分分析法分析并筛选出对各树种具有重要影响的主要叶结构型性状指标;据此,运用SPSS20.0软件中的独立样本T检验法分析毛竹扩张对4乔木层林木总体及不同生活型树种主要叶结构型性状影响,并采用双因素方差分析法分析毛竹扩张对各树种主要叶结构型性状的影响。利用模型lgY=lgα+βlgM(式中Y、M分别代表两主要叶结构型性状指标,α为截距,β为斜率)模拟竹阔混交林与常绿阔叶林内林木主要叶结构型性状间关系,分析毛竹扩张对其主要叶结构型性状关系的影响。利用R3.5.2数据分析软件中的标准化主轴回归分析法(SMA)计算所选模型的截距、斜率及斜率的置信区间,并对斜率进行异质性检验,在斜率同质时计算其共同斜率[20]。
2 结果与分析
2.1 林木主要叶结构型性状指标的筛选
为了更好地揭示各树种叶结构型性状的内在规律,对所测定的LL、LW、LA、SLA、LT、LFW、LDW、LDMC、LTD、LRWC以及LMI等11个叶结构型性状进行主成分分析(PCA)。结果显示:第1主成分(第1排序轴)解释了51.52%的变异,第2主成分(第2排序轴)解释了21.31%的变异,累计解释量为72.83%。由此可见,前两个主成分能较好地反映对树种影响较大的叶结构型性状,且其影响大小主要由第一主成分决定。其中与第一主成分相关最密切的叶结构型性状指标主要为LL、LW、LA、SLA、LTD、LMI及LDMC,且LL、LW、LA和SLA均与第一主成分轴呈正相关,LTD、LMI和LDMC则均与第一主成分轴呈负相关,其相关性大小依次为:LW>LA> LL> LDMC >LTD >SLA >LMI(图2)。因此,本研究确定选取LL、LW、LA、SLA、LTD、LMI和LDMC 共7个指标作为主要叶结构型性状,用于分析毛竹扩张对林木叶结构型性状的影响。
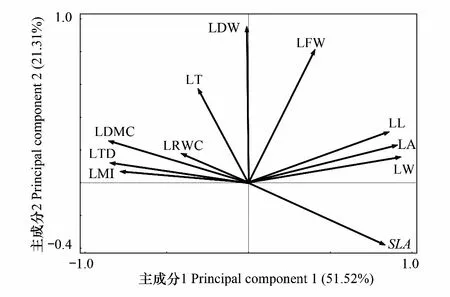
图2 毛竹扩张下4种主要植物叶结构型性状主成分分析图
2.2 毛竹扩张对乔木层林木总体叶结构型性状的影响
以阔叶林中的甜槠、青冈、青榨槭、枳椇4主要树种的各叶结构型性状均值进行分析,结果发现,4林木的LL、LW、LA、LTD、LMI、LDMC及SLA均值在常绿阔叶林与竹阔混交林之间虽存在差异,但差异不明显(P>0.05)(图3),说明毛竹扩张对常绿阔叶林乔木层林木总体叶结构型性状的影响不明显。
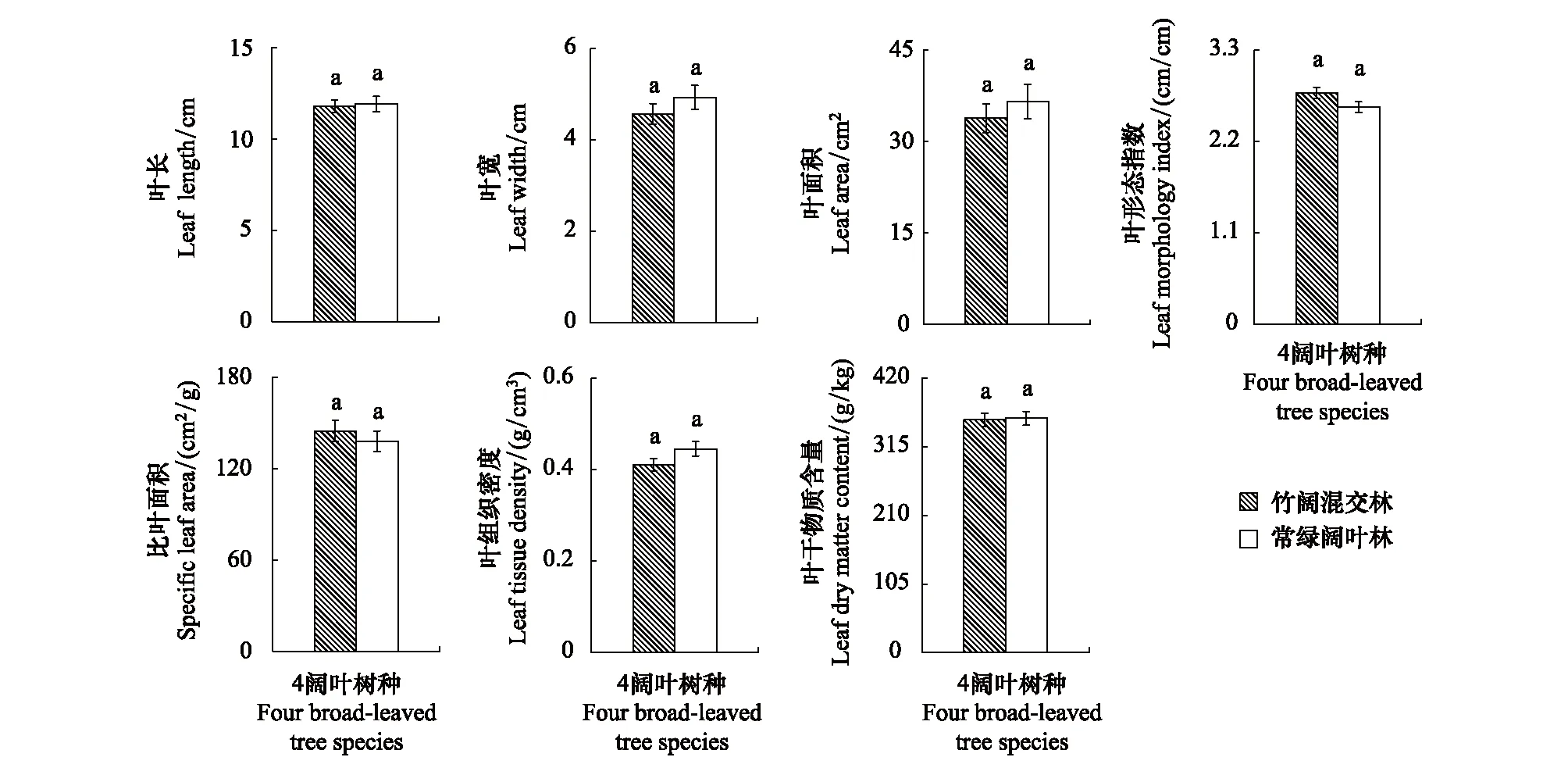
图3 乔木层林木同一叶结构型性状指标在不同林分的表现特征
2.3 毛竹扩张对不同生活型阔叶树种叶结构型性状的影响
通过对两常绿阔叶树种与两落叶阔叶树种的主要叶结构型性状分析,结果显示,毛竹扩张对常绿阔叶林乔木层不同生活型阔叶树种叶结构型性状的影响存在差异(图4),常绿阔叶林内落叶阔叶树种叶结构型性状对毛竹扩张影响的敏感性相对高于常绿阔叶树种。毛竹扩张对常绿阔叶树种的LTD及落叶阔叶树种的LW与LMI两叶结构型性状指标具有显著影响(P<0.05)。竹阔混交林中常绿阔叶树种的LTD显著小于其在常绿阔叶林的LTD(P<0.05),而其LL、LW、LA、SLA、LDMC以及LMI等叶结构型性状指标在竹阔混交林与常绿阔叶林两林分内则差异不显著(P>0.05)。竹阔混交林中落叶阔叶树种的LMI显著高于常绿阔叶林(P<0.05),但其LW则显著小于常绿阔叶林(P<0.05),而其LL、LA、SLA、LTD以及LDMC等叶结构型性状指标在两林分内差异不显著(P>0.05)。

图4 不同生活型树种叶结构型性状指标的比较
2.4 毛竹扩张对不同树种叶结构型性状的影响
不同树种对毛竹扩张影响的响应程度不同(图5)。竹阔混交林中的甜槠LL、LW和LA显著大于常绿阔叶林(P<0.05),但其LMI、SLA、LTD和LDMC在两林分间则差异不显著(P>0.05);毛竹扩张会使竹阔混交林中青冈的LMI显著增加,明显高于常绿阔叶林的LMI(P<0.05),但其LTD、LDMC则显著小于常绿阔叶林(P<0.05),而其LL、LW、LA和SLA在两林分间则差异不显著(P>0.05);竹阔混交林中的枳椇LMI显著高于常绿阔叶林(P<0.05),但其LW与LTD则显著小于常绿阔叶林(P<0.05),而其LL、LDMC、LA和SLA在两林分间则差异不显著(P>0.05);竹阔混交林中的青榨槭LW显著小于其在常绿阔叶林的LW(P<0.05),但其在两林分内的LL、LA、SLA、LMI、LDMC和LTD则差异不显著(P>0.05)。由此可见,随着毛竹向常绿阔叶林的扩张,4乔木层林木的部分叶结构型性状会发生显著的改变。
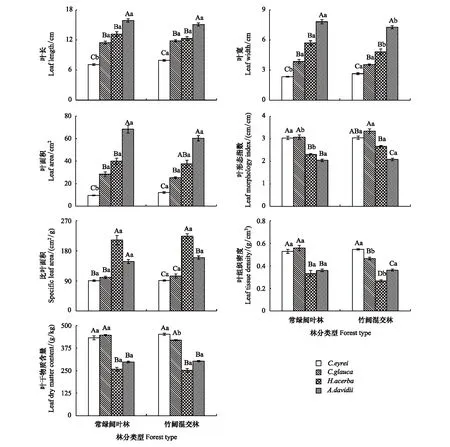
图5 不同树种叶结构型性状指标的比较
毛竹扩张还会改变不同树种间部分叶结构型性状的差异特征(图5)。毛竹扩张会改变乔木层4种林木间LTD与LDMC两性状指标的大小次序。4林木LTD大小次序在常绿阔叶林中其表现为:青冈>甜槠>青榨槭>枳椇,其中,甜槠和青冈的LTD显著大于枳椇及青榨槭(P<0.05),且甜槠和青冈两常绿阔叶树种间及枳椇和青榨槭两落叶阔叶树种间的LTD差异均表现不明显(P>0.05);但在竹阔混交林中其LTD大小次序则表现为:甜槠>青冈>青榨槭>枳椇,且各树种间LTD均存在显著差异(P<0.05)。乔木层4林木间LDMC在常绿阔叶林中的大小次序为:青冈>甜槠>青榨槭>枳椇,但在竹阔混交林中其大小次序则表现为:甜槠>青冈>青榨槭>枳椇;两森林类型均表现为:青冈和甜槠两常绿阔叶树种的LDMC显著大于枳椇和青榨槭两落叶阔叶树种(P<0.05),但两常绿阔叶树种间及两落叶阔叶树种间的LDMC差异均不明显(P>0.05)。4林木间的LMI与SLA虽在常绿阔叶林与竹阔混交林中的大小次序未发生改变,但各树种间差异的显著性则在两森林类型中表现不同。4林木间的LL、LW与LA在常绿阔叶林与竹阔混交林中的大小次序及其差异显著性均表现为一致。上述结果说明,毛竹扩张会改变乔木层4林木间LMI、LTD、SLA及LDMC的差异特征,但不会改变4林木间的LL、LW与LA差异特征。
2.5 毛竹扩张对乔木层林木叶结构型性状间关系影响分析
毛竹扩张会影响常绿阔叶林乔木层林木部分叶结构型性状间关系(图6)。LL与LW、LW与LA在竹阔混交林与常绿阔叶林中虽然均分别拥有共同斜率,但竹阔混交林中LL与LW、LW与LA的截距均显著小于常绿阔叶林,这说明毛竹扩张会改变常绿阔叶林内乔木层林木的LL与LW、LW与LA之间的数量关系。但毛竹扩张对LA与LL、SLA与LA、SLA与LTD、SLA与LDMC间的关系影响较小,它们在竹阔混交林与常绿阔叶林中拥有共同斜率(即异速生长指数)及共同截距(即异速生长常数)(图6),说明毛竹扩张并不会改变常绿阔叶林内乔木层林木LA与LL、SLA与LTD、SLA与LA、SLA与LDMC性状间关系。
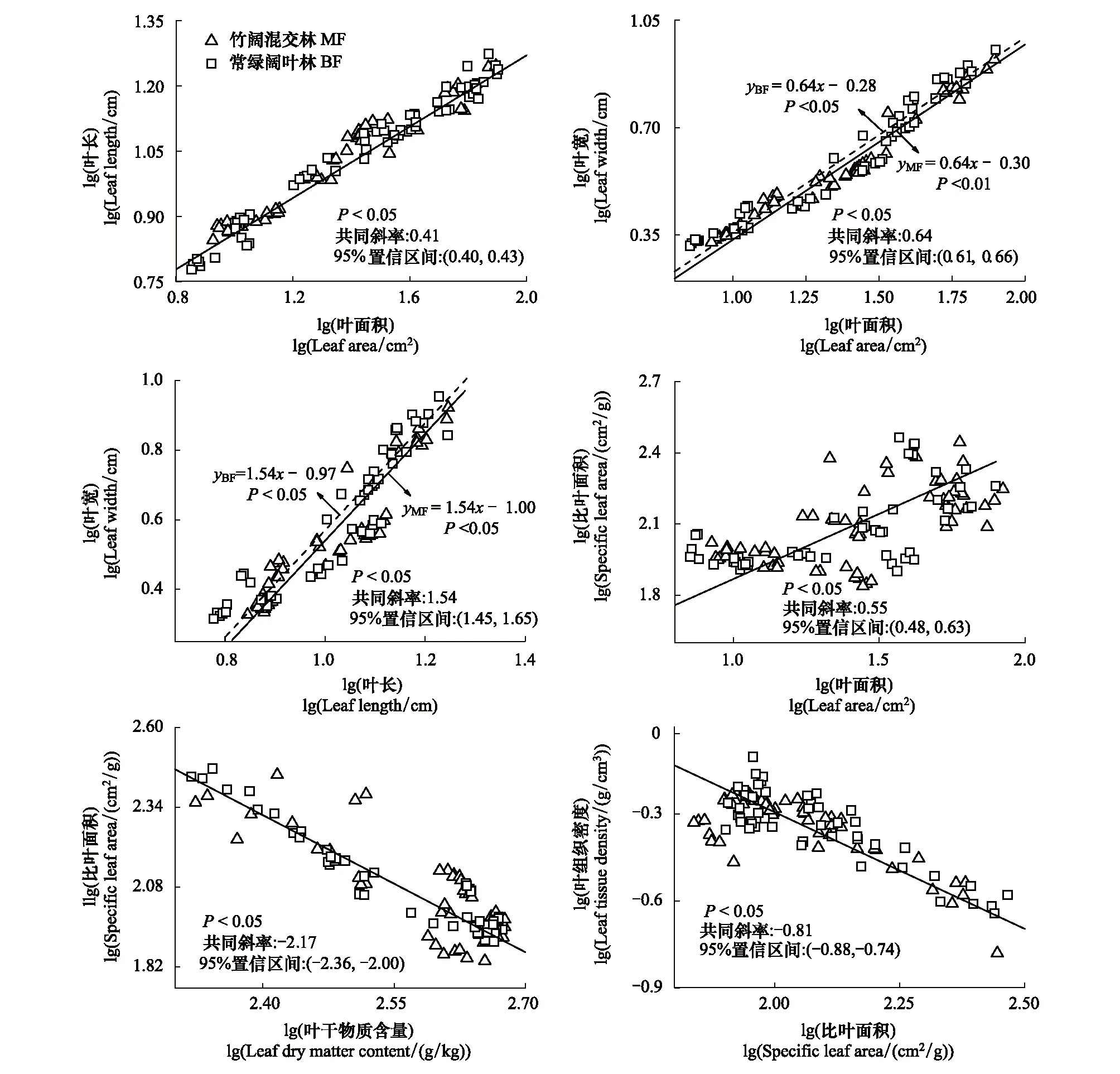
图6 不同林分内林木LL、LW、LA、LTD、LT、SLA之间的线性关系
3 讨论
3.1 毛竹扩张对乔木层林木总体叶结构型性状特征的影响
植物叶性状的分异主要受植物类群、植物体高度和生活型影响,植物类群对植物叶性状的影响要大于环境因子(如温度、水分和经纬度等),以科类群影响为最大[21- 22]。本研究结果显示,4乔木层树种总体叶结构型性状特征在竹阔混交林与常绿阔叶林虽存在差异,但差异不显著,分析原因,主要是由于本研究所选择的树种是两森林类型内共有种,其叶性状主要受植物种影响,毛竹扩张后,虽然会对森林内植物的光照、水分等环境因子造成影响,但环境对叶性状的影响要小于植物种影响,从而导致毛竹扩张对常绿阔叶林乔木层林木总体叶结构型性状影响不明显。
3.2 毛竹扩张对不同生活型阔叶树种叶结构型性状的影响
在森林生态系统中,不同生活型植物的表型可塑性不同。有研究显示,常绿阔叶植物与落叶阔叶植物在SLA、叶片寿命、矿质元素含量等叶性状方面存在差异[23],而常绿阔叶树种在竞争激烈的环境中往往比落叶阔叶树种更占优势[24]。受毛竹扩张影响,常绿阔叶林中的常绿阔叶树种仅有LTD发生显著变化,毛竹扩张会导致常绿阔叶树种LTD显著降低;而落叶阔叶树种则表现为LW与LMI两叶结构型性状指标变化明显,毛竹扩张会导致落叶阔叶树种的LW显著减小、LMI则显著升高,反映了落叶阔叶树种对毛竹扩张的敏感程度相对高于常绿阔叶树种。有研究表明, LTD与植物的碳储备能力有关,LTD较高体现了叶片以慢速周转方式生长,具有较多的碳储备[25]。毛竹扩张会使常绿阔叶林中的常绿阔叶树种LTD显著减小,反映了常绿阔叶树种为适应毛竹扩张而采取开拓性生长策略,即植物主要投资于生长速率和资源获取能力[26],减少碳储备而将更多养分投入到高生长以保持自身的竞争优势。受毛竹扩张影响,阔叶林中的落叶阔叶树种则通过减小LW,增大LMI,调整叶片形态,减少对保护组织的投资,调节叶片对散射光与漫射光的吸收,增大其耐阴性,以提高其对毛竹扩张造成光资源相对不足的环境适应能力[27]。
3.3 毛竹扩张对4乔木层林木叶结构型性状的影响
植物叶片对环境变化敏感且可塑性大[28],可以反映植物对环境的适应能力[8]。不同树种间同一叶结构型性状指标对毛竹扩张影响的响应程度存在差异,同一树种各叶结构型性状指标对毛竹扩张影响的响应程度也不同。甜槠是本研究区域常绿阔叶林中的优势种群,属于壳斗科(Fagaceae),锥属(Castanopsis),具有较高的耐荫性与资源争夺能力[29],毛竹扩张会使甜槠LL、LW以及LA显著增大,即表现为叶片由小到大的变化趋势,反映了甜槠对毛竹扩张后光环境减弱的适应性,增大其光合作用面积,补偿较低光量子通量密度造成的光合速率下降,从而保持其资源竞争优势[30],以促进其生存与生长。青冈为壳斗科青冈属(Cyclobalanopsis),耐干旱瘠薄,是适应能力强的广适种,面对毛竹扩张,其LTD和LDMC显著减小,表明青冈为适应毛竹扩张所引起的养分及水热资源争夺激烈的环境,减少养分的储备以及对结构组织构建的投资,从而将更多的养分用于周转及生长;而其LMI显著增大,叶片从椭圆形向细长型变化,体现了叶片形态对光胁迫的适应。甜槠与青冈虽同属于壳斗科植物,但受毛竹扩张影响,甜槠LL、LW及LA发生显著改变,而青冈的LL、LW及LA则变化不明显,但其LTD、LDMC、LMI则发生了显著改变,这表明即使是同科植物面对毛竹扩张其适应策略也不相同。枳椇为鼠李科(Rhamnales)枳椇属(Hovenia)植物,生长快、易存活、对土壤气候要求不高,毛竹扩张会导致其LW与LTD显著减小、LMI显著增大,说明该树种受毛竹扩张影响,会通过改变叶形态,减小碳存储投资,提高养分运转利用效率以获得竞争优势,从而适应毛竹扩张对其生长的影响。青榨槭为槭树科(Aceraceae)槭属(Acer)乔木,形态可塑性高,适应性强,既可在沟谷、林缘等光源充足的开阔地生长,又可在光源竞争激烈的林下生长[31],毛竹扩张仅会造成其LW显著减小,说明该树种的叶结构型性状受毛竹扩张影响较小。
3.4 毛竹扩张对乔木层林木叶结构型性状间关系的影响
植物为了适应环境的变化,通过权衡资源分配和调节生理过程,在形态上表现出特殊的植物功能性状的变化[32-33],进而形成一系列性状组合[34-35],具有一定的可塑性。毛竹扩张会对乔木层林木部分叶结构型性状间关系产生显著影响,使乔木层林木LL与LW、LW与LA间的截距显著减小,表明乔木层林木面对毛竹扩张,在相同的LL投入下,竹阔混交林中乔木层林木LW的投入相对较小;而在相同的LA投入下,竹阔混交林中乔木层林木LW的投入也相对较小。这主要是由于毛竹扩张加剧植物对光、水、热等资源的竞争,而水胁迫会影响叶片的形态。有研究显示,LW减小有利于植物叶片的直接散热,而不通过蒸腾降温[36],防止水分流失,提高乔木层林木对毛竹扩张造成的林地土壤水分降低压力的适应能力。
毛竹扩张可能还会对常绿阔叶林内乔木层其他林木的叶功能型性状产生影响,并可能会对茎干与细根功能性状产生影响,这些研究有助于探清常绿阔叶林主要树种对毛竹扩张的响应与适应机制。
4 结论
毛竹扩张对常绿阔叶林乔木层4树种总体叶结构型性状无明显影响,但其对不同生活型阔叶树种叶结构型性状的影响存在差异,落叶阔叶树种叶结构型性状对毛竹扩张影响的敏感性相对高于常绿阔叶树种。毛竹扩张会造成乔木层树种的部分叶结构型性状发生显著改变。受毛竹扩张影响,同一叶结构型性状变化在不同树种间存在差异;同一树种不同叶结构型性状对毛竹扩张的响应程度也存在差异。毛竹扩张会导致常绿阔叶林乔木层树种部分叶结构型性状间关系发生改变,它会改变林木LL与LW、LW与LA之间的数量关系。
猜你喜欢
建筑与预算(2022年6期)2022-07-08
农村实用技术(2020年1期)2020-12-16
意林·少年版(2020年13期)2020-08-02
汉语世界(2020年3期)2020-06-19
东坡赤壁诗词(2020年2期)2020-06-04
绿色中国(2019年18期)2020-01-04
现代园艺(2018年14期)2018-01-18
农家科技下旬刊(2017年8期)2017-11-13
科技创新导报(2016年21期)2016-12-17
学与玩(2009年2期)2009-03-09
