
贮藏温度对方格星虫与革囊星虫质构、内源酶与水分变化的影响
2020-08-17 08:34:54白云霞庞一扬江虹锐刘小玲
食品工业科技 2020年15期
卢 鑫,白云霞,庞一扬,江虹锐,刘小玲
(广西大学轻工与食品工程学院,广西南宁 530000)
方格星虫(Sipunculuslnnaeus,1766)又名沙肠子、沙虫等,为方格星虫科(Sipunculidae rafinesque,1814)[1];革囊星虫(Phascolosomaleuckart,1828)又名海泥虫、泥丁等,为革囊星虫科(Phascolosomatidae stephen et edmonds,1972)[2]。两种星虫均无节,呈蠕虫状,属海产体腔动物,在我国沿海从南到北均有分布,其中以广西海区资源较丰富[1]。星虫味道鲜美、营养丰富,药用价值较高,故有“海洋虫草”的美誉,是一种高值海珍品,具有重要经济价值[3]。目前研究学者针对星虫的研究主要集中在物种分类、基因研究、人工养殖、营养分析、功能活性[4-6]、加工风味[7-8]与组织结构[9-11]等方面,关于其不同温度贮藏期间的质构品质变化未见报道。
星虫具有独特的质构特征,表现为质地优良,口感脆嫩,深受广大消费者的青睐,而贮藏过程对星虫的质构品质会产生显著影响。水分、内源蛋白酶活、组织结构变化是影响水产品品质优劣的重要因素[12-13]。Zhang等人研究发现鲢鱼质构与肌原纤维极显著相关[14]。而内源蛋白酶及水分的变化将直接影响纤维结构[15-16]。
本实验主要比较了不同贮藏温度对方格星虫与革囊星虫质构、纤维结构、内源蛋白酶活及水分特性变化的影响,综合反映其质构品质变化规律,为后期更好实现星虫保活保鲜提供理论参考。
1 材料与方法
1.1 材料与仪器
方格星虫(SN)、革囊星虫(PE) 广西南宁安吉海鲜市场采购;Tris-HCl 北京索莱宝科技有限公司;酪蛋白 北京索莱宝科技有限公司;三氯乙酸(TCA) 成都市科隆化学品有限公司;以上试剂均为分析纯。
TA.XT plus型物性测定仪 英国Stable Micro systems公司;F16502型扫描电镜 荷兰PHENOM公司;Himac CR21N型高速冷冻离心机 日本日立公司;Lab Master-aw型水分活度仪 瑞士Novasina公司;SQP型电子天平 德国Sartorius公司;MR20-025V-1型核磁共振成像分析仪 上海纽迈电子科技有限公司。
1.2 实验方法
1.2.1 星虫样品的贮藏实验 将市场上采购的鲜活方格星虫(体长13±1 cm,体宽1.0±0.2 cm,体质量15±2 g)、革囊星虫(体长15±5 cm,体宽1.0±0.2 cm,体质量10±2 g)置于人工气候培养箱中贮藏,光照强度控制在20%,湿度控制在60%;通过预实验观察两种星虫分别在4 ℃贮藏48 h,30 ℃贮藏22 h后开始出现死亡现象;因此选择分别于4 ℃下贮藏0、4、8、16、24、36、48 h,30 ℃下贮藏0、2、4、6、10、14、22 h后,随机抽取3~5条,除去内脏等不可食部分,对其质构特征、组织结构、内源蛋白酶活、水分分布、水分含量、水分活度进行测定。
1.2.2 质构特征测定 采用BSK剪切探头测定两种星虫剪切力、咀嚼性,剪切测试程序的主要参数为:选用HDP/BS探头,测试前速度:1 mm/s测试速度:5 mm/s测试后速度:10 mm/s剪切距离:50 mm触发力:5.0 g;;采用P/2N穿刺探头测定两种星虫硬度、韧性,穿刺测试程序的主要参数为:选用P/2N(针形)探头,测试前速度:1 mm/s;测试速度:1 mm/s;测试后速度:1 mm/s;穿刺距离:10 mm;触发力:5.0 g。
1.2.3 组织结构扫描电子显微镜(SEM)观察 参考肖桂华[12]的SEM样品处理及观察方法,将在4和30 ℃贮藏不同时间的两种星虫除去内脏等不可食部分清洗并沥干表面水分后置于-80 ℃预冻12 h后,冷冻干燥48 h(冷阱温度-50 ℃,真空度10 Pa隔热板温度30 ℃),对冻干后的样品切成1.0 cm×0.5 cm方形组织块,对其横切面采用离子溅射喷金,SEM放大1000倍对两种星虫肌肉组织环肌层(CML)进行观察。
1.2.4 内源蛋白酶活的测定 将不同时间贮藏后的整条星虫匀浆后称取5.00 g,在4 ℃的条件下,按料液比1∶4 (m:v),加入20 mL pH8.0的Tris-HCl,4 ℃下浸泡6 h,4 ℃下1000 r/min离心10 min,取上清液即为粗酶液。取两支试管为1和2,各加入4.00 mL的Tris-HCl缓冲液和1.60 mL的1%酪蛋白后,往1加入4.00 mL的10% TCA,振荡摇匀1 min,两支试管放入35 ℃水浴保温5 min,往两支试管各加入0.80 mL的粗酶液,摇匀振荡,1 min,35 ℃水浴反应15 min,2中加入2.40 mL的10% TCA,摇匀振荡1 min,将两支试管中样品10000 r/min离心15 min,取上清液,以1为空白组,2为反应组,于275 nm波长下测定吸光值[17]。
单位酶活力(U)定义[18]为:在275 nm波长下,单位质量样品在单位时间内水解酪蛋白底物而增加0.001吸光度值所需的酶量为1个酶活力单位。
酶活力(U/g)=ΔOD/[15 min×0.001×(0.8 mL÷20 mL)×5 g]
式中:ΔOD为实验组与空白组吸光值变化量;15 min为反应时间;0.8 mL为所取粗酶液体积,20 mL为总粗酶液体积,5 g为样品质量。
1.2.5 水分特性的测定
1.2.5.1 水分分布的测定 使用核磁共振成像分析仪,参考巩涛硕等[19]和宋朝鹏等[20]的方法,将星虫样品切成长宽为2 cm×1 cm肉块,擦干样品表面水分,称量重量在0.80±0.02 g,保鲜膜包裹后置于低场核磁共振(LF-NMR)分析仪中。CPMG序列参数:回波个数NECH=15000,回波时间TE=0.6 ms,累加采样次数NS=4,每个样品重复三次。
1.2.5.2 水分含量的测定 参照《GB 5009.3-2016 食品安全国家标准 食品中水分的测定》,采用直接干燥法测定方格星虫与革囊星虫在不同贮藏阶段的水分含量[21]。
1.2.5.3 水分活度的测定 使用水分活度测定仪测定(温度设置为25 ℃)。
1.3 数据处理
每组试验三个平行,用Excel 2016对数据进行处理,采用SPSS 20.0 Duncan法进行显著性差异分析(P<0.05表示差异显著),使用Origin 9.1进行作图。
2 结果与分析
2.1 不同温度贮藏过程中星虫质构特性的变化
因样品厚度仅1~2 mm,不适用压缩模式进行质构测定,故选用穿刺与剪切模式测定星虫质构特征:硬度、韧性、剪切力、咀嚼性。两种星虫在4及30 ℃贮藏过程中质构的变化分别如图1(A)~(D)、(E)~(G)所示。SN在4 ℃下贮藏0~48 h,硬度、韧性、剪切力、咀嚼性呈下降趋势,在16~24 h显著下降(P<0.05),故4 ℃贮藏时间应控制在0~16 h,贮藏16 h其硬度、咀嚼性分别下降19.08%、33.94%。在30 ℃贮藏0~22 h,除韧性上升其他呈下降趋势,且硬度、剪切力、咀嚼性在10~14 h变化显著(P<0.05),因此30 ℃贮藏应控制在10 h内,其硬度、咀嚼性分别下降41.61%、41.39%。
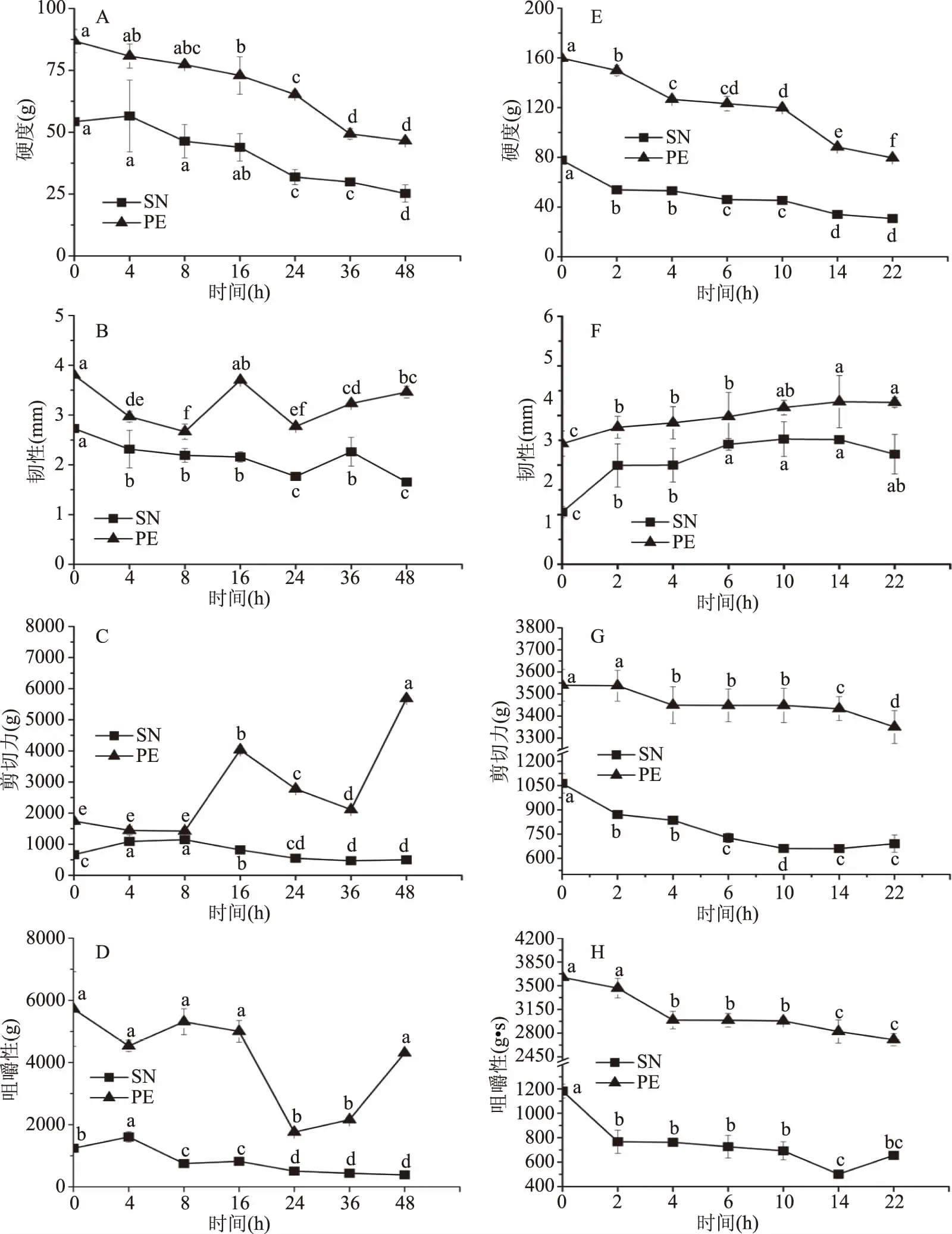
图1 方格星虫与革囊星虫在4、30 ℃藏期间的质构变化Fig.1 Changes of SN and PE’ texture during 4 and 30 ℃ storage注:同折线上不同小写字母代表差异显著(P<0.05),图3~图6同;图(A)~(D)为4 ℃贮藏,(E)~(H)为30 ℃贮藏。
PE在4 ℃贮藏0~48 h,其硬度与韧性变化趋势与SN一致,剪切力与咀嚼性先下降后上升,在0~16 h内硬度和咀嚼性分别下降16.06%、12.62%;在30 ℃贮藏0~22 h,其质构变化趋势同样与SN一致,在10~14 h质构变化显著(P<0.05),其硬度、咀嚼性分别下降25.02%、17.68%。
上述结果表明,星虫贮藏过程中质构除韧性外均下降,4 ℃低温贮藏相比30 ℃贮藏能够保持星虫质构时间更长。冯丁丁[22]研究海参在4、20 ℃条件下质构变化,结果显示在20 ℃条件下1/4 d硬度与咀嚼性明显下降,在4 ℃下15~25 d硬度和咀嚼性急剧下降,因原料及样品处理差异导致贮藏时间不一,但表明4 ℃低温贮藏能有效减缓质构的变化,与本文研究结果一致。
2.2 不同温度贮藏过程中星虫微观结构的变化
质构与肌原纤维蛋白极显著相关[14],星虫组织结构以肌肉组织为主,肌肉组织主要由肌原纤维构成,因此对贮藏过程中星虫肌原纤维结构变化进行观察。SN在4及30 ℃贮藏过程中不同时间点肌肉组织变化如图2(A)、(B)所示,在4 ℃贮藏0~16 h,环肌层(CML)肌原纤维由排列整齐的直线状,逐渐出现弯曲现象且纤维间空隙增大;在贮藏16 h后如图中标记处可以看出,由于肌肉组织皱缩严重,纤维排列更加密集,与0 h处肌肉纤维差异明显,故4 ℃贮藏SN应控制在16 h内,其肌肉纤维变化相对较小。在30 ℃贮藏,CML肌原纤维随贮藏时间增长,逐渐皱缩变细,纤维间空隙变大,在前10 h变化不明显,14、22 h变化较大,纤维皱缩严重,间隙明显增大,故30 ℃贮藏SN应控制在10 h内,其肌肉纤维变化相对较小。
PE在不同温度下贮藏其肌肉组织变化如图2(C)、(D)所示,在4 ℃贮藏0~16 h,PE纤维变粗,逐渐出现杂乱,16 h后开始出现皱缩断裂,24 h处变化明显;在30 ℃贮藏肌肉纤维随贮藏时间增长,纤维宽度变细,纤维排布由疏松变紧密,纤维间空隙变小,贮藏14 h纤维与0 h相比出现明显变化。故PE在4 ℃及30 ℃贮藏应分别控制在16、14 h内。
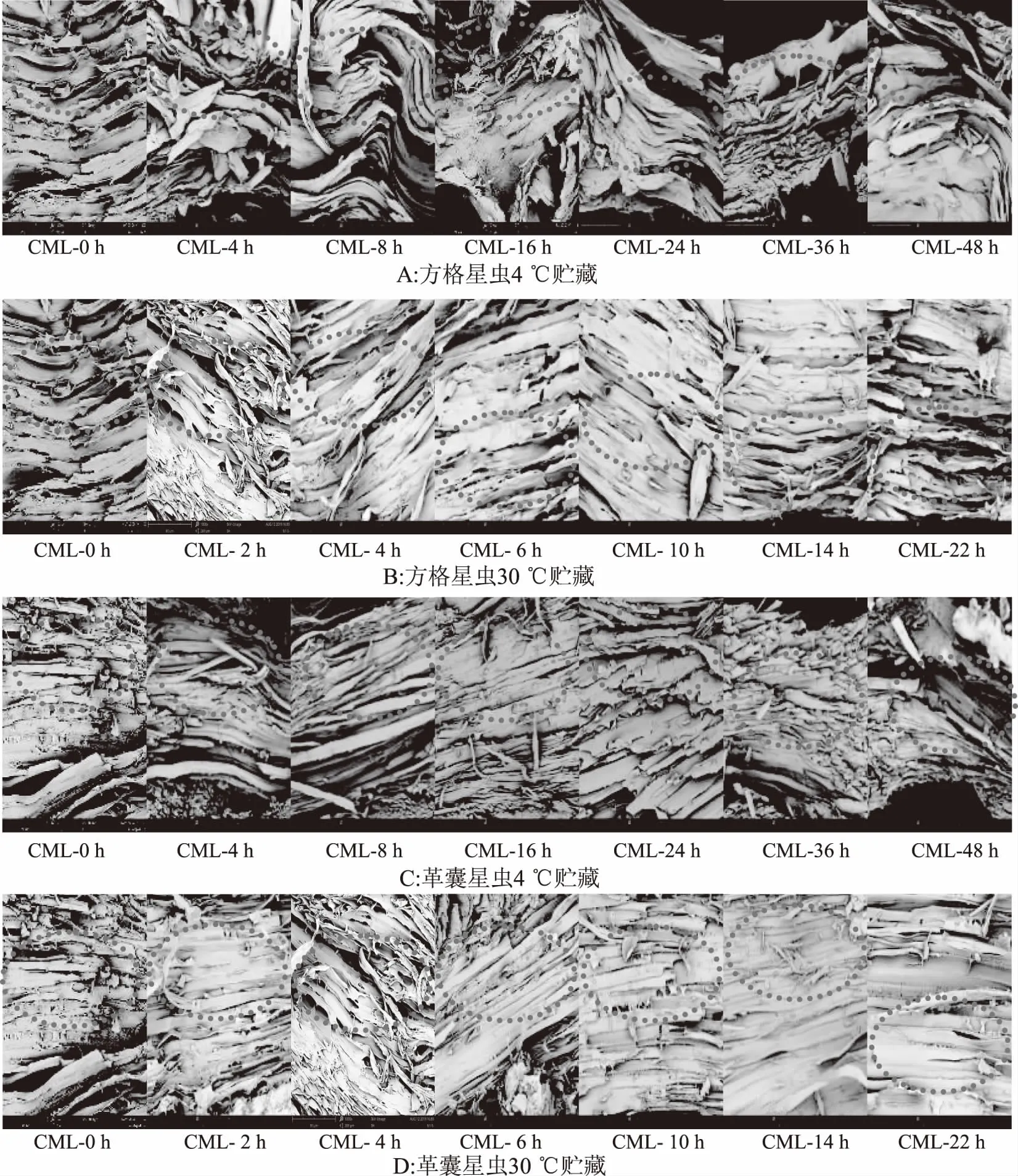
图2 星虫在4、30 ℃贮藏期间的肌肉组织变化Fig.2 Change of SN and PE’s muscle tissue during 4 and 30 ℃ storage
上述结果表明,星虫在贮藏过程中纤维结构会发生明显变化,且4 ℃比30 ℃贮藏纤维变化程度更缓慢。鱼肉组织中与星虫组织同样存在大量肌原纤维,沈妮[23]研究带鱼在0与4 ℃下贮藏组织结构的变化,发现随贮藏时间延长,肌纤维间隙增大,紊乱松散,甚至出现断裂,且温度越低越能够有效抑制带鱼肉蛋白的变性降解,越有利于维持带鱼肉肌纤维的完整性。其研究结果中肌原纤维变化规律与本研究相似,且4 ℃相比于30 ℃温度更低,其中4 ℃低温贮藏星虫纤维更完整,变化程度更小,两者结果相一致。
2.3 不同温度贮藏过程中星虫内源蛋白酶活的变化
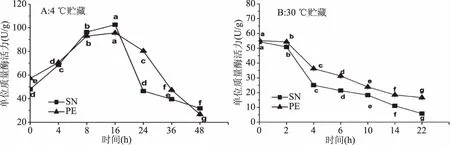
图3 星虫在4、30 ℃贮藏过程中星虫内源蛋白酶活性的变化Fig.3 Change of PE and SN’ endogenous protease activity during 4 and 30 ℃ storage
内源蛋白酶直接影响蛋白变化,蛋白变化对纤维组织结构影响较大,进而引起质构劣变[23],因此对贮藏过程中星虫内源蛋白酶活变化进行观测。星虫在4与30 ℃贮藏过程中单位质量内源蛋白酶活的变化如图3(A)、(B)所示。SN于4 ℃贮藏在0~16 h单位质量内源蛋白酶活由48.0 U/g上升至102.6 U/g,16 h后酶活显著下降(P<0.05),最终在48 h降至31.8 U/g;SN在30 ℃贮藏随时间增长,酶活呈下降趋势,0、10、22 h SN内源蛋白酶活分别为54.4、18.3、5.8 U/g,10 h后酶活显著下降(P<0.05)。
PE在4 ℃贮藏在0~16 h酶活由57.1 U/g升至95.7 U/g,16 h后酶活显著下降(P<0.05),最终在48 h下降至26.8 U/g。星虫在4 ℃贮藏单位质量内源蛋白酶呈先上升后下降的趋势,与王博[24]研究中国蛤蜊、菲律宾蛤仔、三疣梭子蟹、锯缘青蟹、虾夷扇贝在0~4 ℃贮藏内源蛋白酶活整体呈先上升后下降的趋势一致。星虫在4 ℃贮藏16 h及30 ℃贮藏0~22 h随时间增长酶活下降,可能由于细胞逐渐凋亡瓦解导致酶活降低[25]。
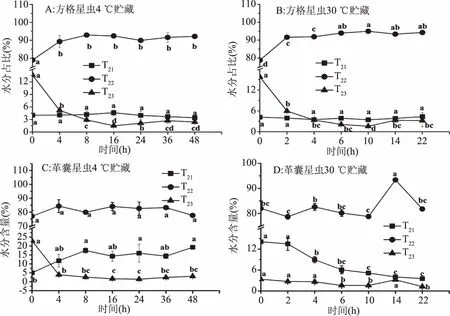
图4 星虫在4、30 ℃贮藏过程中不同水分占比的变化Fig.4 Changes of different water percent of SN and PE during 4 and 30 ℃ storage
2.4 不同温度贮藏过程中星虫水分特性的变化
2.4.1 水分分布的变化 水分的分布变化直接影响纤维结构,进而对质构产生影响[15-16]。星虫在4、30 ℃贮藏过程中不同水分变化如图4所示,根据弛豫时间可以将水分分为:T21(1~10 ms)为结合水,T22(10~100) ms为不易流动水,T23(100~1000) ms为自由水[26],结果显示星虫中T22占比最高。SN在4 ℃贮藏过程中,T21水分占比在0~16 h变化不显著(P>0.05),T22水分占比呈上升趋势,在0~16 h占比上升13.42%,16 h与0 h相比显著上升(P<0.05),T23水分占比呈下降趋势,在16 h下降达12.11%,与0 h相比显著下降(P<0.05);SN在30 ℃贮藏过程中,T21水分占比在0~10 h变化不显著(P>0.05),因为它具有0.1~1 ms的短弛豫时间,并且这些分子与大分子和蛋白质紧密相关,因此波动不大[27]。T22水分占比呈上升趋势,在10 h占比最高,且上升16.19%,与0 h相比显著上升(P<0.05),T23水分占比呈下降趋势,在10 h占比最低,下降达14.18%,与0 h相比显著下降(P<0.05)。
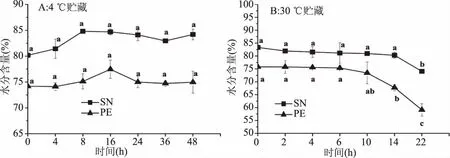
图5 星虫在4、30 ℃贮藏过程中水分含量的变化Fig.5 Change of water content of SN and PE during 4 and 30 ℃ storage
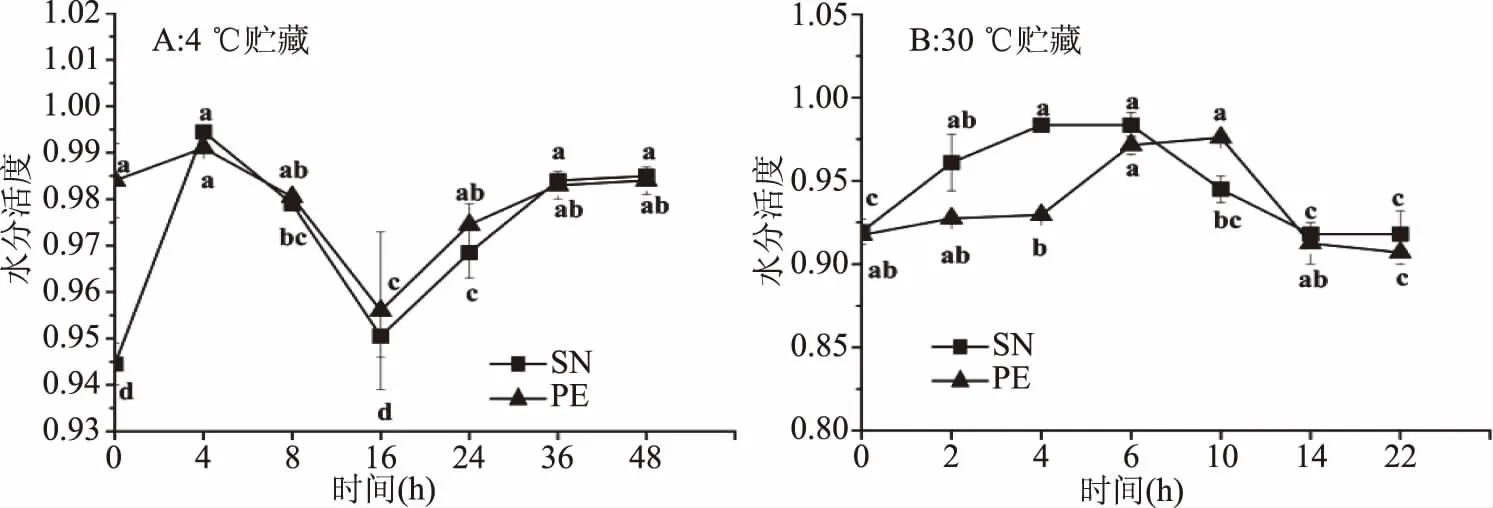
图6 星虫在4、30 ℃贮藏过程中水分活度的变化Fig.6 Change of water activity of SN and PE during 4 and 30 ℃ storage
PE贮藏过程中水分分布变化与SN基本一致。不同温度贮藏星虫T21结合水占比变化不显著(P>0.05),在4 ℃贮藏T22不易流动水占比上升率高于30 ℃,T23自由水占比下降率低于30 ℃,可能由于贮藏温度越高导致自由水流失量大。崔宏博等[28]研究南美白对虾在贮藏过程中水分分布的变化时发现,T22不易流动水占比下降,T23自由水占比上升,与星虫水分变化相反,可能由于两种样品处理方式及样品内部结构差异导致。
2.4.2 水分含量的变化 两种星虫虫体水分含量超80%[29],在贮藏过程中对其含量变化的观测具有必要性。在4 ℃贮藏,随时间增长星虫水分含量变化如图5(A)所示,SN水分含量高于PE,变化不显著(P>0.05),整体呈先上升后下降的趋势,SN在贮藏8 h水分含量最大,达84.82%;PE在贮藏16 h水分含量达到最大值77.46%。
在30 ℃贮藏下,两种星虫水分含量的变化如图5(B)所示,整体呈下降趋势,在0~10 h下降不显著(P>0.05);SN前10 h含水量由83.34%下降至80.96%,22 h最终下降至74.05%,PE在相同时间点水分含量分别为75.81%、73.44%、59.10%。星虫30 ℃贮藏10 h后水分含量显著下降(P<0.05),与贮藏过程中水分蒸发损失有关[28]。30 ℃贮藏水分呈下降趋势且变化比4 ℃贮藏更明显,这是由于低温贮藏能抑制某些微生物的生长活动,温度升高贮藏时间延长微生物生长繁殖速率加快,对蛋白质等营养物质的分解加快,星虫结合水的能力下降,导致水分含量下降[30]。
2.4.3 水分活度的变化 水分活度反映了水分子的存在状态以及自由水与结合水的比例[31]。星虫在4 ℃贮藏过程中不同时间点水分活度由图6(A)可得,星虫水分活度呈先下降后上升趋势,两种星虫水分活度变化趋势基本一致,在16 h处水分活度最低分别为0.95、0.96,在36 h后趋于稳定。说明在4 ℃贮藏16 h前星虫中水分的结合程度呈上升趋势,16 h以后随着水分流失,水分的结合程度降低。
星虫在30 ℃贮藏过程中不同时间点水分活度由图6(B)可得,两种星虫水分活度整体呈先上升后下降的趋势,两种星虫水分活度上升下降幅度较一致,SN水分活度在30 ℃贮藏6 h后开始下降,PE在10 h后出现下降。说明在30 ℃贮藏,星虫水分的结合程度先下降后上升,由于不易流动水向自由水移动,且星虫水分损失严重导致。
SN与PE水分活度大小相差较小,4 ℃贮藏与30 ℃贮藏过程两种水分活度变化相反,可能由于在4 ℃贮藏中星虫水分流失较小,水分结合度高,则水分活度先下降,随着贮藏时间越长,16 h后水分流失,结合程度下降,则水分活度上升;在30 ℃贮藏中星虫水分流失较大,水分结合度低,则水分活度先上升,随着贮藏时间越长,SN达到6 h,PE达到10 h后水分基本不明显流动,结合程度上升,则水分活度下降。
3 结论
星虫在4 ℃下贮藏0~16 h相对30 ℃贮藏0~10 h,其质构特征下降程度较小,且纤维组织变化相对不明显,内源蛋白酶活及水分特征变化相对缓慢;4 ℃贮藏16~24 h与30 ℃贮藏10~14 h各指标结果发生明显变化。综上所述低温4 ℃贮藏星虫能有效延缓其质构变化,表现为纤维组织、内源酶活和水分特征相应变化程度。因此在对方格星虫革囊星虫运输贮藏过程中,4 ℃贮藏应控制在0~16 h,30 ℃贮藏应控制在0~10 h的时间范围,从而降低贮藏对星虫品质影响,此研究对星虫贮藏过程中食用价值的保护具有重要意义。
猜你喜欢
材料与冶金学报(2017年3期)2017-09-21 01:54:49
上海农业学报(2016年2期)2016-10-27 00:49:48
核科学与工程(2016年3期)2016-01-03 07:22:11
华东理工大学学报(自然科学版)(2015年4期)2015-12-01 04:00:23
动物营养学报(2015年10期)2015-12-01 02:26:21
云南中医学院学报(2014年3期)2014-07-31 18:58:03
化学与生物工程(2014年1期)2014-03-27 09:26:57
中国有色金属学报(2011年10期)2011-11-24 08:37:08
故事作文·高年级(2009年11期)2009-12-11 10:24:24
故事作文·高年级(2009年9期)2009-12-11 10:24:18
