
壳聚糖复合硅酸钠对冬枣采后苯丙烷代谢及病程相关蛋白的影响
2020-08-17 08:34张静茹张乐乐常璐璐张少颖
食品工业科技 2020年15期
张静茹,张乐乐,常璐璐,张少颖
(山西师范大学食品科学学院,山西临汾 041004)
冬枣为鼠李科(Rhamnaceae)枣属(Zizyphus),别名冰糖枣、苹果枣、雁来红、冻枣[1]。因其果皮薄,果肉脆甜,果核小,可食率高且富含丰富的维生素、多种矿物质、多酚、花色苷以及抗癌物质环磷酸腺苷(cAMP)和环磷酸鸟苷(cGMP),成为被大众所喜爱的一种鲜食枣品。然而其采后呼吸强度大,对空气中的CO2敏感,极易失水、褐变、软化、酒化和腐烂,在常温条件下贮藏3 d左右就会失水、软化、霉烂[2]。在冬枣收获、储存、加工和运输的过程中都容易出现由链格孢菌(A.alternata)引起的黑斑病,导致冬枣腐烂。这不仅造成了冬枣的品质下降,更造成了经济上的损失。有研究表明A.alternata还可在寄主细胞内产生真菌毒素,对人体健康造成潜在危害[3]。近年来主要采用化学杀菌剂来控制真菌引起的病害。但是,化学杀菌剂的使用会造成环境污染、菌株抗药性增强、致癌作用等问题,寻找无污染、无公害的新的防治植物采后病害的方法成为当务之急。
壳聚糖(CTS)是甲壳素脱乙酰基产生的一种多糖类物质。因其具有无毒无味、生物可降解性、选择透过性和抑菌性等优点,已被广泛应用于果蔬贮藏[4]。CTS可抑制或者杀灭水果上的多种病原菌,例如链格孢菌、灰霉菌、枯草芽孢杆菌[5]等。有研究表明,CTS能够增强马铃薯[6]的苯丙烷类代谢酶的活性以及提高梨[7]防御相关酶的活性。谢春晖[8]发现用不同浓度CTS处理冬枣,与其他处理冬枣相比,1% CTS涂膜处理效果最佳,冬枣腐烂率最低,硬度、可滴定酸下降最缓慢。
硅作为地球表面第二大元素,在植物抗病性方面有着重要的作用。硅酸钠作为硅的供体,可抑制杏果实[9]和冬枣[10]黑斑病等。Na2SiO3可以通过苯丙烷途径激活活性氧代谢从而提高果实的抗逆性,保持果蔬品质[11]。Na2SiO3与其他抗菌试剂的复合处理也能增强果实的抗病性,有研究报道Na2SiO3与脱氢乙酸钠联合使用对在贮藏期间柑橘果实酸腐病菌(Geotrichumcitri-aurantii)引起的酸腐病有显著的抑制作用[12]。CTS和Na2SiO3处理对冬枣采后抗病性的影响已有研究,但二者结合使用对于果蔬的抗病性影响研究较少。通过结合使用可以发挥各自的特性,提高保鲜的总体效果。因此,本实验旨在研究CTS和Na2SiO3复合处理对冬枣果实苯丙烷代谢及相关防御酶的影响,以期丰富CTS和Na2SiO3复合处理在果蔬采后抗性诱导作用方面的研究,同时为果蔬采后病害防治提供一种新方法。
1 材料与方法
1.1 材料与仪器
供试白熟期冬枣采自山西省临汾市尧乡冬枣种植基地,手工采摘,装箱后迅速运回实验室,挑选大小均一、无病虫害机械损伤的冬枣作为试验用果;链格孢菌(A.alternata) 在PDA上培养备用,北纳生物有限公司;壳聚糖(相对分子质量:15~20万之间;脱乙酰度:87%~90%) 山东奥康科技公司;硅酸钠、硼砂、双氧水、氢氧化钠 分析纯,天津市风船化学试剂科技有限公司;聚乙烯吡咯烷酮 Sigma;L-苯丙氨酸 天津博迪化工股份有限公司;愈创木酚 武汉东康源科技有限公司;p-香豆酸 上海邦景事业有限公司;β-巯基乙醇、福林酚 山东西亚化学工业有限公司。
759S紫外可见分光光度计 上海棱光技术有限公司;IMS-40全自动雪花制冰机 常熟市雪科电器有限公司;CP214电子天平 常州奥豪斯仪器有限公司;H1850R台式高速冷冻离心机 湖南湘仪实验室仪器开发有限公司;MDF-U53V超低温冰箱 日本三洋集团;SpectraMax M2/M2e酶标仪 上海普迪生物科技有限公司;DZKW-D-4恒温水浴锅 上海科恒实业发展有限公司。
1.2 实验方法
1.2.1 孢子悬浮液的配制 参照He等的方法[13],将斜面链格孢菌接种于PDA平板上,在28 ℃下恒温培养7~10 d。孢子成熟后,用含有0.05% 吐温20的生理盐水将其从平板上刮下,用双层纱布过滤掉多余菌丝,在显微镜下观察计数,将其最终孢子悬浮液的浓度调成1×105个/mL。
1.2.2 损伤接种及取样 将冬枣平均分成5组,分别用蒸馏水、1% CTS、1% CTS+10 mmol/L Na2SiO3、1% CTS+40 mmol/L Na2SiO3、1% CTS+160 mmol/L Na2SiO3浸泡5 min,自然晾干。每组100个冬枣,重复三次。用打孔器在冬枣的赤道处打出一个深2 mm,直径为3 mm的小孔,放置1 h。将5 μL孢子悬浮液打入小孔内,将接种后的果实放入大保鲜盒内,用保鲜膜封口。室温(22 ℃),相对湿度(RH)90%下贮藏,每隔4 d取样,进行病斑直径的测定。
取冬枣损伤处皮下0.2~10 mm范围内的果肉,液氮冷冻后磨粉,保存在-80 ℃的超低温冰箱,用于后期指标的测定。
1.2.4 苯丙烷类代谢相关酶活性的测定 苯丙氨酸解氨酶(PAL)活性的测定参照曹建康等[14]的方法,适当修改。取4 g枣粉,加入4 mL提取液(含有40 g/L PVPP、2 mmol/L EDTA和5 mmol/Lβ-巯基乙醇)。4 ℃、12000×g离心30 min,收集上清液,即为提取酶液。PAL反应体系为3 mL 50 mmol/L pH8.8的硼酸缓冲液、0.5 mL 20 mmol/L L-苯丙氨酸溶液、0.5 mL酶液。煮沸 6 min作为对照,37 ℃下保温1 h,加入0.1 mol 6 mol/L HCl终止反应,在290 nm处测定其吸光度值。以每小时每克冬枣实(鲜重)酶促反应体系吸光度值增加0.01为1个PAL活性单位(U),单位是U·h-1·g-1FW。
肉桂酸4-羧化酶(C4H)的测定方法参照范存斐等[15]的方法,适当修改。取1 g枣粉,加入3 mL提取液(50 mmol/L pH8.9 Tris-HCl,15 mmol/Lβ-巯基乙醇、4 mmol/L MgCl2、5 mmol/L VC、5 μmol/L亮肽素、5 mmol/L PMSF、0.15% PVP、10%甘油),4 ℃、12000×g离心25 min,取上清液为提取酶液。反应体系包括0.8 mL酶液,2.2 mL缓冲液(50 mmol/L pH8.9 Tris-HCl、2 μmol/L反式肉桂酸、2 μmol/L NADPNa、5 μmol/L G-6-PNa2),加入0.1 mL 6 mol/L HCl终止反应。在340 nm处测定其吸光度值。OD值每分钟变化0.01作为一个酶的活性单位,单位是U·min-1·g-1FW。
4-香豆酰-辅酶A连接酶(4CL)的测定方法参照张杼润等[16]方法,适当修改。取2 g枣粉,加入6 mL预冷的0.2 mol/L的Tris-HCl(含有25%甘油和0.1 mol/L DTT,pH为8.0)缓冲液,4 ℃、12000×g离心20 min,取上清液为提取酶液。1 mL酶液中依次加入1.5 mL反应液(含有0.9 mL 15 μmol/L MgSO4、0.3 mL 5 μmol/L P-香豆酸、0.3 mL 5 μmol/L CoA)、0.3 mL 50 μmol/L ATP,40 ℃水浴10 min后加入0.1 mol 6 mol/L HCl终止反应,在333 nm处测定其吸光度值。OD值每分钟变化0.1 作为一个酶的活性单位(U),单位是U·min-1·g-1FW。
POD活性的测定参照曹建康等[14]的方法。取3 g枣粉,加入3 mL提取缓冲液(含1 mmol/L PEG、4% PVPP、1% TritonX-100)混匀后于4 ℃、12000×g离心30 min,取上清液为提取酶液。0.5 mL酶液中加入3 mL 25 mmol/L愈创木酚溶液,0.2 mL 0.5 mmol/L H2O2混匀,470 nm处测定吸光度值的变化。以每分钟吸光度变化1所需的酶量为一个POD单位,单位是U·min-1·mg-1protein。
1.2.5 总酚、类黄酮和木质素的测定 总酚含量的测定参照Xiao等[17]方法并修改。配制含有0、0.02、0.04、0.06、0.08、0.10、0.12、0.14、0.16、0.18 mg/mL的标准没食子酸溶液。取0.2 mL各浓度标准液,加入2 mL Folin-Ciocalteau试剂(稀释10倍)混匀后室温放置5 min,然后加入2 mL 10% Na2CO3溶液,室温避光放置90 min,在765 nm测定其吸光度值。以吸光度对浓度进行线性回归分析,求得回归方程和决定系数分别为A=4.3216C+0.0694,R2=0.997。取0.5 g枣粉,加入4 mL 80%的乙醇,混匀后于4 ℃、8000×g离心20 min。取0.2 mL上清液,依次加入Folin-Ciocalteau、Na2CO3溶液,测量方法与制作标曲方法相同。样品中总酚的含量用没食子酸当量(mg·g-1)来表示。
类黄酮含量的测定参照Jia等[18]方法并修改。配制含有0、0.4、0.8、1.2、1.6、2.0 mg/mL的芦丁标准溶液。各取0.1 mL,加入0.3 mL 5% NaNO2摇匀后静置5 min,加入0.3 mL 10% Al(NO3)3放置6 min。最后加入2 mL 1 mol/L NaOH溶液,室温下放置15 min,在510 nm处测定其吸光度值。以吸光度对浓度进行线性回归分析,求得回归方程和决定系数分别为A=0.0732C-0.0067,R2=0.9967。样液的制备与测量总酚方法相同。取0.1 mL上清液,依次加入NaNO2、Al(NO3)3、NaOH溶液,测量方法与制作标曲方法相同。样品中的黄酮含量用芦丁的当量(mg·g-1)来表示。
木质素含量的测定参照Morrison[19]方法并作修改。取1 g枣粉加入4 mL预冷的95%乙醇混匀,于4 ℃、12000×g离心10 min。沉淀物用95%乙醇冲洗3次,再用乙醇∶正己烷=1∶2 (v/v)冲洗3次。收集沉淀干燥,干燥后加入1 mL 25%溴乙酰,70 ℃恒温水浴30 min。加入1 mL 2 mol/L NaOH溶液中止反应。再加入2 mL冰醋酸和0.1 mL 7.5 mol/L盐酸羟胺。再次离心,取上清液0.25 mL用醋酸定容至5 mL。在280 nm处测定其吸光度值。木质素含量用OD280·g-1FW表示。
在明确了著作权所特有的性质后,我们就可得知,未经许可演绎人取得著作权后,法律并未赋予其以法律规定以外的方式利用其作品的权利。其是否有权对其作品实施控制,仍须得到原作品著作权人的许可。这是因为,哪里有独创性表达,哪里就有保护,未经许可演绎作品中包含有原作品著作权人的独创性表达,因此原作品著作权人可依法对未经许可演绎作品进行控制,以禁止或许可未经许可演绎人乃至于第三人利用未经许可演绎作品。而赋予未经许可演绎人著作权,仅赋予了其许可或禁止他人以特定方式利用其作品的权利。他人欲利用未经许可演绎作品,需获得双重许可,除了得到未经许可演绎人的许可之外,还须得到原作品著作权人的许可。
1.2.6 病程相关酶活性的测定 CHT活性的测定参照曹建康等[14]的方法。取3 g枣粉,加入5 mL预冷丙酮,混匀后于4 ℃、12000×g离心30 min。反应体系中依次加入0.5 mL 50 mmol/L乙酸-乙酸钠缓冲液(pH=5.2)、0.5 mL 10 g/L 胶状几丁质悬浮液,反应管和对照管各加入 0.5 mL酶液,对照管煮沸5 min。反应管37 ℃水浴1 h后加入0.1 mL 30 g/L的脱盐蜗牛酶混匀继续水浴1 h,然后加入0.2 mL 0.6 mol/L四硼酸钾溶液,煮沸3 min后迅速冷却,于585 nm处测定吸光度值。酶活性单位是U·mg-1protein。
PPO活性的测定参照曹建康等[14]的方法。粗酶液与POD活性测定的提取方法相同。反应体系中依次加入2 mL 50 mmol/L乙酸-乙酸钠缓冲液(pH=5.5)、1 mL 50 mmol/L邻苯二酚溶液,最后加入0.1 mL酶液。于420 nm处测定吸光度值的变化,以每分钟吸光度变化增加1为一个活性单位,单位是U·min-1·mg-1protein。
GLU活性的测定参照曹建康等[14]的方法。粗酶液与CHT活性测定的提取方法相同。反应体系加入100 μL 4 g/L的昆布多糖、100 μL酶液,煮沸5 min的酶液作为对照。反应管37 ℃水浴40 min后,依次加入1.8 mL蒸馏水、1.5 mL DNS试剂,沸水加热5 min,用蒸馏水稀释至20 mL,混匀。于540 nm处测定吸光度值。酶活性单位是U·mg-1protein。
1.3 数据处理
以上指标均取3个平行样,重复测定3次。使用Excel 2010计算平均值并绘制图表,SPSS statistics 20进行Duncan’s多重比较进行差异分析。图中不同字母表示差异显著(P<0.05)。使用Adobe Photoshop CC 2018作图。
2 结果与分析
2.1 CTS+Na2SiO3处理对果实病斑直径的影响
贮藏期间冬枣病斑直径逐渐增大,且处理组的病斑直径显著低于对照组(P<0.05)。前12 d各处理组之间并差异不明显,贮藏后期整体来说CTS+Na2SiO3处理效果优于CTS单独处理(图1和图2)。第20 d CTS处理组的病斑直径比复合40、160 mmol/L Na2SiO3处理组的高出36.29%、21.11%。CTS+40 mmol/L Na2SiO3处理组对病斑直径的抑制效果最好,因此,选择CTS+40 mmol/L Na2SiO3处理组进行后续指标的分析。

图1 CTS+Na2SiO3处理对冬枣挑战接种A. alternata病斑直径的影响Fig.1 Effects of CTS+Na2SiO3 treatment on colony diameterin winter jujube inoculated with A. alternata注:不同小写字母表示不同处理组间差异显著(P<0.05)。
2.2 CTS+Na2SiO3处理对果实苯丙烷代谢相关酶活性的影响

图2 CTS+Na2SiO3处理对冬枣挑战接种A. alternata贮藏20 d后各处理的病斑扩展情况Fig.2 Effects of CTS+Na2SiO3 treatment on theexpansion of lesions in winter jujube inoculatedwith A. alternata after storage for 20 d注:对照(a);1% CTS处理(b);1% CTS+10 mmol/LNa2SiO3处理(c);1% CTS+40 mmol/L Na2SiO3处理(d);1% CTS+40 mmol/L Na2SiO3处理(e)。
PAL、C4H、4CL是苯丙烷代谢途径中的三个关键酶,POD处于苯丙烷代谢途径末端[20]。随着贮藏时间的延长,果实中PAL、C4H的活性呈现出先上升后下降的趋势,且在第12 d出现活性高峰(图3A、B)。从第8 d开始处理组与对照组出现显著差异(P<0.05)。在第20 d CTS+Na2SiO3处理组PAL、C4H比对照组分别高出37.16%、120.48%。如图3C所示,4CL的活性呈上升趋势且复合处理组的活性高于对照组,贮藏4 d之后差异显著(P<0.05)。尤其是在第8 d,CTS+Na2SiO3处理组是对照组4CL活性的1.73倍。贮藏期间枣果实中POD的变化趋势与C4H相似,但其活性峰值出现在第8 d(图3D),比C4H活性高峰提前4 d出现。
2.3 CTS+Na2SiO3处理对果实总酚、类黄酮和木质素的影响
总酚、类黄酮、木质素是苯丙烷代谢途径的重要产物。总酚和类黄酮是苯丙烷代谢的最终产物,与植物抗氧化活性[21]及果实抗病性密切相关。如图4A、4B所示,在整个贮藏期间,CTS+Na2SiO3处理组与对照组的总酚与类黄酮含量均呈先下降再上升最后下降趋势,在第12 d出现峰值。CTS+Na2SiO3处理在贮藏中后期(8~20 d)显著提高了枣果实的总酚、类黄酮含量(P<0.05)。贮藏结束时,CTS+Na2SiO3处理组果实的总酚、类黄酮含量分别比对照组高17.34%、6.18%。
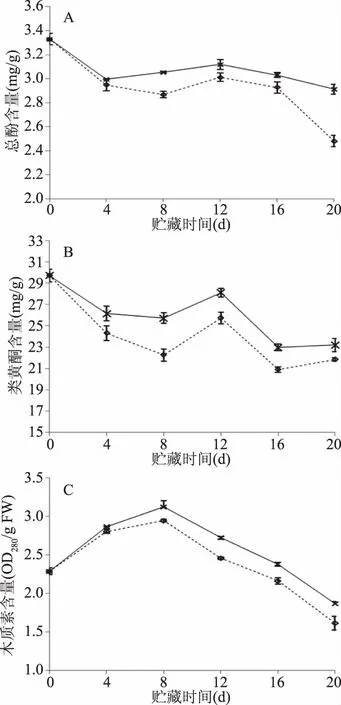
图4 CTS+Na2SiO3处理对冬枣挑战接种A. alternata苯丙烷代谢产物的影响Fig.4 Effect of CTS+Na2SiO3 treatment on metabolites ofphenylpropan in winter jujube inoculated with A. alternata注:总酚含量(A)、类黄酮含量(B)和木质素含量(C)。
木质素是植物细胞壁成分之一,可以通过与细胞壁的结合使植物组织木质化,保护细胞免受病原菌的侵害。如图4C所示,各处理枣果实中的木质素含量呈先上升后下降的趋势,在第8 d达到峰值且第12 d CTS+Na2SiO3复合处理木质素含量比对照组高11.02%。CTS+Na2SiO3复合处理冬枣的木质素含量在贮藏8~20 d内均显著高于对照组(P<0.05)。
2.4 CTS+Na2SiO3处理对CHT、PPO和GLU活性的影响
CHT是一种存在于植物中以几丁质为底物的一种水解酶,GLU的水解底物是某些真菌和一些病原菌细胞壁的主要成分,这两种酶对于植物防卫反应具有协同作用。各处理组在贮藏过程中的CHT、GLU活性均先升高后降低,对照外CHT的活性高峰出现在第8 d,GLU的活性高峰推迟4 d出现(图5A和图5B),其酶活性分别是对照组的2.5倍和1.47倍。复合处理组CHT、GLU活性从第8 d开始显著高于对照组(P<0.05)。
PPO可以将植物中的酚类物质转化为醌和单宁从而抑制病原菌的扩散。贮藏期间PPO的活性呈上升趋势,且在第12~16 d之间对照组活性显著高于复合处理组(P<0.05,图5C)。
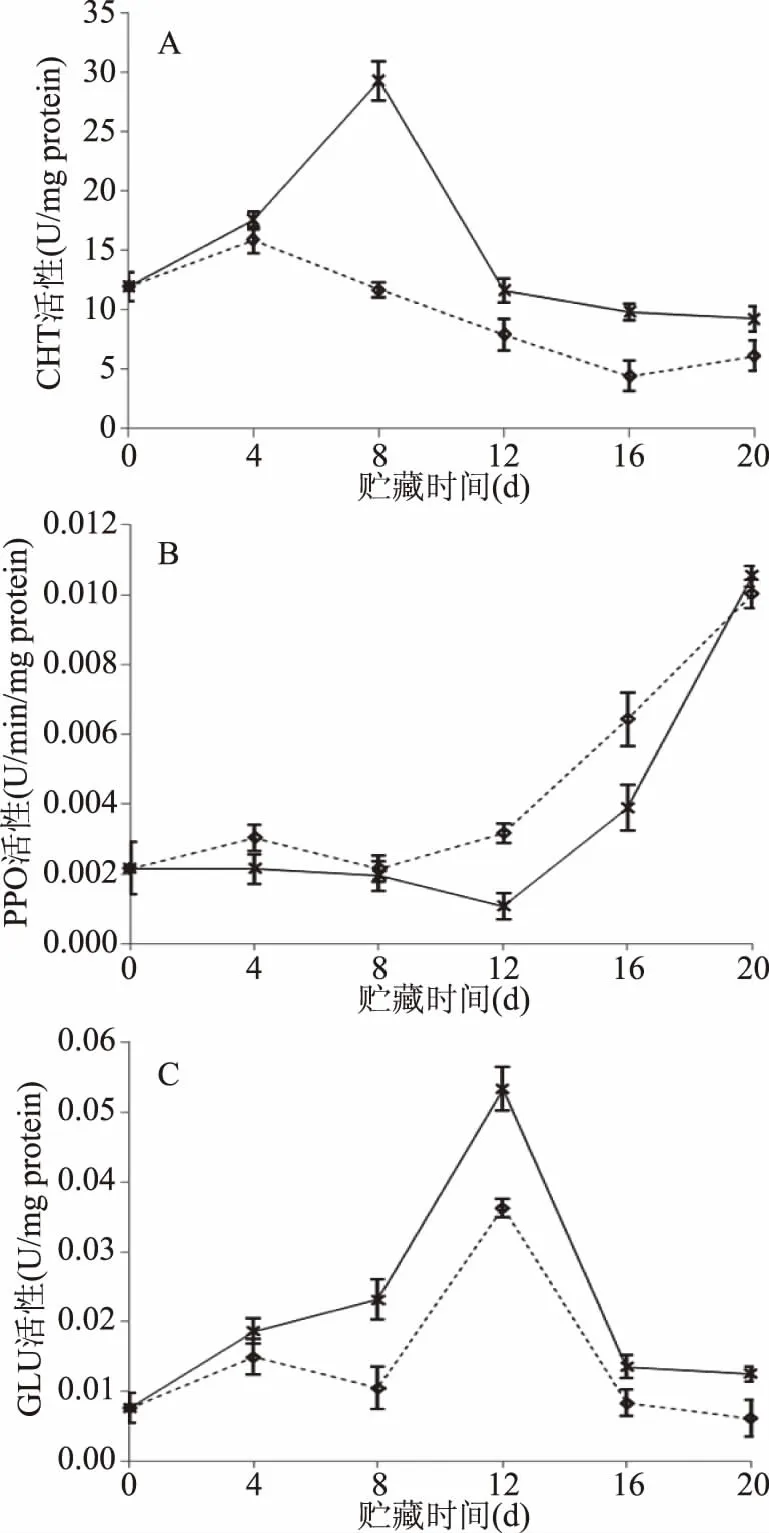
图5 CTS+Na2SiO3处理对冬枣挑战接种A. alternata病程相关酶活性的影响Fig.5 Effect of CTS+Na2SiO3 treatment on theactivities of enzymes related to the pathogenesisin winter jujube inoculated with A. alternata注:CHT活性(A)、PPO活性(B)和GLU活性(C)。
3 讨论

苯丙烷途径是植物特有的次生代谢途径,植物受到逆境胁迫时,其代谢产物酚类、类黄酮可以作为抗病毒剂,保护植物免受生物或非生物胁迫,并且赋予果蔬颜色及味道[25]。PAL是苯丙烷衍生物途径中的第一个关键限速酶,可以催化苯丙氨酸转化为反式肉桂酸。C4H将反式肉桂酸转化为P-香豆酸。4CL是苯丙烷途径中最后一个关键酶,可以催化肉桂酸为相应的辅酶A酯,控制苯丙烷代谢主途径向分支途径转折,其下游次级代谢产物为花青素、水杨酸、木质素等其他参与抗性反应的物质[26]。CTS可以诱导植物产生萜类、异黄酮、生物碱等植保素,进而诱导抗性蛋白活性提高并且木质素含量增加。CTS处理苹果后,其PAL 和 POD 活性增加,总酚、类黄酮和木质素的含量也有所上升[27]。研究表明Na2SiO3处理可以提高SOD、PAL活性,诱导杏果实总酚、木质素增加,提高杏果实的活性氧与苯丙烷代谢[11]。魏娟等[28]研究表明Na2SiO3处理提高了甜瓜果实PAL、CHS基因mRNA的表达量,说明Na2SiO3处理可在基因转录水平上促进苯丙烷代谢途径的关键酶的表达,从而提高果实抗病性。本研究表明,CTS+Na2SiO3处理冬枣,激活了PAL、C4H、4CL及POD等苯丙烷代谢相关酶的活性,且增加了总酚、类黄酮、木质素等苯丙烷代谢产物的含量。总酚、类黄酮、木质素的增加可能与PAL活性的增加有关,第12 d时PAL、总酚、类黄酮同时达到峰值,但木质素含量在第8 d达到峰值,说明CTS+Na2SiO3处理可能通过其他途径诱导木质素的形成。4CL也可以通过催化香豆酸合成酚类物质前体,其活性高低与其他苯丙烷代谢酶的平衡可调节酚类物质的合成有关[29]。POD参与了木质素的合成,并且氧化酚类物质形成对病原菌毒害较大的醌类物质[27]。木质素含量的提高可以增强果实细胞壁抵御病原菌的能力。这表明CTS+Na2SiO3可通过调节植物的苯丙烷代谢酶的活性及产物积累,增强冬枣的抗病性,维持其采后品质。
病程相关蛋白是指植物在病理或病理相关的环境下产生的一类可以直接攻击病原菌的蛋白,受病原菌或者其他外部胁迫诱导表达,与植物的抗病性及其适应环境胁迫有重要作用。CHT与GLU的底物分别是真菌细胞壁中的几丁质与葡聚糖,可水解真菌细胞壁,从而具有抗真菌作用。Meng等[30]发现CTS处理损伤接种后的梨果实,会显著增强其CHT、GLU、POD的活性,并抑制病原菌的生长。本研究发现,CTS+Na2SiO3处理冬枣提高其CHT、GLU活性,但抑制其PPO活性。这与CTS处理F.sulphureum挑战接种马铃薯块茎研究结果相反[31]。可能是CTS+Na2SiO3复合处理改变了果蔬的内部环境,也可能是因为实验材料和病原菌的不同引起了差异。同时,实验结果也暗示,CTS+Na2SiO3复合处理并不能通过提高PPO活性来抑制A.alternata引起的冬枣腐烂。CTS+Na2SiO3处理增加了果实抗病性,不仅表现在苯丙烷代谢的增强,也表现在抑制病原菌和采后病害的发生。CHT、GLU、PAL、POD活性增加,可以抵御病原菌的扩张。CTS+Na2SiO3处理对A.alternata抑制作用及其如何在果实内部发挥信号分子作用还需进一步研究。
4 结论
1% CTS+40 mmol/L Na2SiO3能有效地控制冬枣采后病害的发生,贮藏效果最佳。CTS+Na2SiO3复合处理采后冬枣,可减少病斑直径的扩展;增加苯丙烷途径相关酶PAL、C4H、4CL、POD活性,并且促进了苯丙烷途径产物总酚、类黄酮、木质素的形成;提高了抗病相关蛋白CHT、GLU活性,但不能诱导PPO活性的升高。实验结果表明,CTS+Na2SiO3复合处理能调节冬枣的苯丙烷代谢,诱导病程相关蛋白活性升高,从而增强冬枣的抗病性。
猜你喜欢
家庭医药(2021年18期)2021-12-03
保健与生活(2021年13期)2021-07-14
能源工程(2021年1期)2021-04-13
农产品市场周刊(2020年23期)2020-01-25
今日农业(2019年12期)2019-08-13
电子制作(2019年10期)2019-06-17
恋爱婚姻家庭(2019年30期)2019-03-23
石油石化绿色低碳(2019年6期)2019-01-14
天然产物研究与开发(2018年11期)2018-11-30
灾害医学与救援(电子版)(2018年1期)2018-06-05
