
人工林内外气象要素差异分析
2020-08-12 13:34翟禄新晏忠凤程楠迟晋浙梁晟荣胡婷郭颜钰
防护林科技 2020年6期
翟禄新,晏忠凤,程楠,迟晋浙,梁晟荣,胡婷,郭颜钰
(广西师范大学环境与资源学院,广西师范大学可持续发展创新研究院,岩溶生态与环境变化研究广西高校重点实验室,广西 桂林 541006)
森林与水的关系是生态水文学研究的核心问题[1]。森林生态水文研究已经成为水循环与生物圈相互作用的一个重要领域。森林冠层、枯落物及林下土壤都参与水循环的过程,是水循环中必不可少的部分。因为森林有遮挡、吸收等作用,使得林内的热量、水分等都得到重新分配,与林外相比产生了新的时空分布格局[1]。气象要素成为林内外生态水文效应的重要指标。
目前,国内外对广西桂北地区典型树种的生态水文效应研究还比较少。为了研究林内外的生态水文效应,在广西桂北地区典型树种的林外林内建立自动气象站,收集林内外气象数据,同时在林内布置土壤探头,并尝试采用“原位观测”对枯落物持水能力进行定量观测。在获取一定气象数据的基础上,对收集到的数据进行处理分析,以发现林内外生态水文规律,研究结果可为涵养水资源、防止水土流失、区域水资源管理、认识和理解生态水文效应、探索森林如何影响和改善小气候提供科学依据。
1 研究区概况
研究区位于广西壮族自治区桂林市,桂林市地处广西东北部(25°15′23.3″ N,110°19′25.5″ E,平均高程150 m)。该地区属于中亚热带湿润季风气候,夏季雨量充沛,冬季温和少雨,年平均降水量1 900 mm左右,年平均气温19 ℃;地带性土壤为红壤,有显著的脱硅富铝化成土特征,pH4.0~6.0,酸性较强;该地区树种类型丰富,天然林破坏较为严重。为此,我们选取了在该地区具有代表性的树种进行研究,林区内树种不唯一。试验区位于广西师范大学雁山校区田径场东北侧,主要树种包括榕树(FicusmicrocarpaLinn. f.)、刺槐(RobiniapseudoacaciaL.)、高山榕(FicusaltissimaBl.)、黄葛(Ficusvirensvar.sublanceolata)、枫香树(LiquidambarformosanaHance)、银杏(GinkgobilobaL.)、桂花树(OsmanthusfragransLour.)等。
2 研究方法
2.1 林内外水文要素测定
根据《地面观测气象规范》,在选取样方的林地内外架设自动气象站。对2019年10月—2020年4月的林内外温度、湿度、太阳辐射、风向、风速、降水等气象要素实行1次·h-1的定时观测。
2.2 计算分析方法
通过自动气象站定时观测收集林内外温度、湿度、风速、降水等气象要素,利用人工定时称质量观测枯落物质量。采用SPSS统计软件对林内外气象要素进行一元线性回归和相关分析,并利用平均截流量分析林内冠层截留效应。利用matlab软件借助彭曼蒙特斯公式计算林内外潜在蒸发量。
3 结果分析
3.1 冠层截留效应
植被冠层是森林水文生态效应的第一个活动层,是植被水源涵养及森林水文系统中降水再分配的首要层级。因此,植被冠层截留降水过程是水循环过程中的重要环节[3]。植被冠层截留雨量包括降雨终止时被截留在树体表面的雨水和在降雨过程中通过蒸发作用从树体表面返回大气的雨量[4]。降水强度、降水量和森林郁闭度是影响植被冠层截流量的重要指标,其中降水量占主导地位。冠层截留的基本规律是:随着森林郁闭度的增加,冠层截留量显著增大;随着林外降水量增加,冠层截留量也会显著增大,但是当林外降雨量增加到一定值后,冠层截留量增加趋势减小,直至保持不变;一般降雨量相同的情况下,随着降雨强度增大,冠层截留量减小。选取10次降水分析他们和林冠层截留之间的关系,如表1。
由表1可见,在选取的10次样本容量中,12月2日、12月25日、12月30日、1月2日等4次降水雨量较少,降水量不足1 mm,由于受森林郁闭度的影响,降水大部分或全部被冠层截留,透冠雨量基本为0,截留达到100%;11月13日、12月1日、12月20日和12月26日等4次降水雨量相对较多,降水量达到5 mm左右,在不同森林郁闭度影响下,冠层截留量为48.89%~64.91%,截留效应较强;12月21日降水量较多,冠层截留为5.84%,占比在10%以下,这表明随着降雨量的增加截留量呈现下降趋势。表明在不同森林郁闭度的影响下,冠层截留量一般随着降雨量增大而增加,但增加到一定程度之后,随着降雨量增加,冠层截留量呈减少趋势。
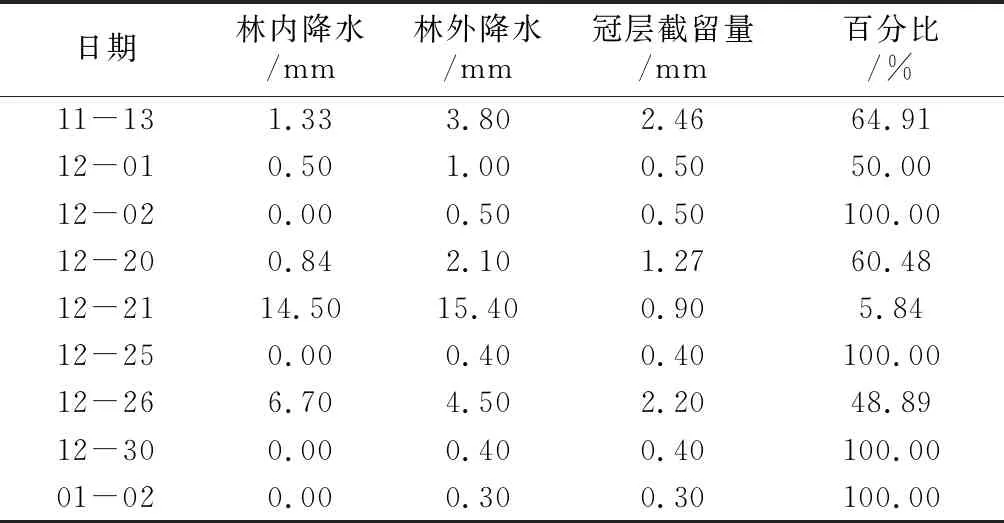
表1 林内外降水与冠层截留量记录表

表2 冠层截留量与降水量一元回归标准化系数
由表2可见,其中R表示拟合优度(相关系数),用R衡量林外降水和冠层截留量的拟合程度,R=0.175,表示只考虑降水量的情况下,冠层截流量和林外降水总量的相关性不明显。表中调整后的R2=0.066,表示林外降水仅仅可以解释冠层截留量6.6%的变化。因此拟合效果较差(图1)。
3.2 林内外气象要素对比
林内外温湿度出现差异的主要原因是由于森林冠层对太阳辐射的吸收和地面长波辐射作用导致的,茂密的森林冠层也阻碍林内外空气的交换,降低了林内风速,使得林内林木及其他植物蒸腾和林地蒸发等产生的水汽在林内保持较多,导致林内的空气湿度高于林外[5]。
3.2.1 温度 温度与林木生长紧密相关,它是维持植物生命活动的重要气象因子。通过对比分析林内外温度差异,尝试说明林内外温度存在差异的主要原因。

表3 林内温度与林外温度一元回归标准化系数一览表
由表3可见,相关系数R=0.966,说明林内温度和林外温度相关性很强,并且呈现正相关,林外温度越高,林内温度也越高。R2=0.933,表明林外温度可以解释林内温度93.3%的变化,拟合效果较好(图2)。
图3显示(选取11月6日至1月7日数据),冬季林内温度变化剧烈,林外温度变化波动较小,且由于受到森林的影响,冬季林内温度始终高于林外温度。另外由于太阳辐射能大部分被植被冠层吸收或反射、散射出去,且林内温度本身就高,风速较小。由于林内空气温度变化主要依靠林内外空气对流运动实现,因此林内温度变化略微滞后于林外温度变化[6]。
3.2.2 湿度 林分对林内空气湿度的调节作用是非常明显的,但是不同季节表现出的调节作用是有很大差别的。

表4 林内湿度与林外湿度一元回归标准化系数一览表
由表4可见,相关系数R=0.825,说明林内湿度和林外湿度相关性很强。并且呈现比较明显的正相关关系,林外湿度越大,林内湿度也越大,主要是因为林内林木和灌丛分布密集,其蒸腾作用显著,从而使得林内湿度较大。R2=0.680,表明林外湿度能解释林内湿度68%的变化,拟合效果良好(图4)。
由图5(选取11月6日至1月7日数据)可以看出,林外湿度波动较大,可能是受到降水的影响,林外湿度不稳定,而林内湿度主要受到植物蒸腾的影响,波动比较小。
3.2.3 风速 风速影响林内外的空气交换。在茂密林内风速很大程度上受到森林郁闭度的影响,会使得林内风速远小于林外风速,从而影响林内外的潜在蒸散发量。

表5 林内风速与林外风速一元回归标准化系数一览表
由表5可见,相关系数R=0.829,说明林内风速与林外风速具有良好的相关性,并且呈现出相对明显的正相关关系,但是由于林内风速变化受到森林郁闭度和冠层叶面积指数等因素的影响较大,所以拟合图分布效果一般,总体来说,林外风越大,林内风速也越高。R2=0.686,表明林外风速能解释林内风速68.6%的变化,拟合效果一般(图6)。
图7显示(选取11月6日至1月7日数据),在选取的研究区内冬季林内风速远小于林外风速。
3.3 林内外蒸发能力分析
潜在蒸散发是指具备充分土壤水分条件下垫面的全部蒸发量[7]。严格意义上来说,蒸散发表示水分从陆面转化为水蒸气进入大气的所有过程总和,包括水面蒸发、陆面蒸发和植物蒸发[8]。
目前来看,潜在蒸散发估算方法主要分为3种类型[7]:(1)基于辐射的方法,包括Priestley-Taylor、Makkink等;(2)基于温度的方法,包括Thomthwaite等;(3)基于辐射、温度和空气动力学项的Penman-Monteith方法。Penman-Monteith方法计算精度高,常常被作为潜在蒸散发(ET0)的估算标准。本文所应用的参数比较简单,仅获取了温度和总辐射的数据,利用matlab软件借助彭曼蒙特斯(Penman-Monteith)公示计算林内外潜在蒸发量。
图8显示(选取11月7至1月6日数据),利用彭曼蒙特斯公式计算出林内外的潜在蒸散发分布差异明显。潜在蒸散发与温度、太阳辐射、海拔高度、风速等很多因素相关。在冬季,温度较低,太阳辐射较弱,加上林冠层的影响,林内的直接蒸发很小,大多数是由植物蒸腾作用引起的水分蒸发,相比于林内,林外的大多数蒸发量是由水面和陆面蒸发带来的,因此潜在蒸发量较大。

表6 林内潜在蒸发与林外潜在蒸发一元回归标准化系数
由表6可见,相关系数R=0.765,说明林内潜在蒸发与林外潜在蒸发具有较强的相关性,并且呈现出明显的正相关关系,林外潜在蒸发较强时林内潜在蒸发也会增加。R2=0.577,说明林外潜在蒸发能解释林内潜在蒸发57.7%的变化,二者拟合效果良好(图9)。
4 结论
4.1 在不同森林郁闭度的影响下,冠层截留量随着降雨量增大而增加,增加到一定程度之后,随着降雨量增加,冠层截留量减少。
4.2 林内温度和林外温度相关性很强,并且呈现正相关。冬季林内温度变化剧烈,林外温度变化波动较小,林内温度变化略微滞后于林外温度变化,且由于受到森林郁闭度的影响,冬季林内温度始终高于林外温度。
4.3 林内湿度和林外湿度相关性很强,并且呈现比较明显的正相关关系。林内湿度较大,但林内湿度波动相比林外较小。
4.4 林内风速与林外风速具有良好的相关性,并且呈现出相对明显的正相关关系。在大量分布植被的林地内风速很大程度上受到森林郁闭度的影响,使得林内风速远小于林外风速。
4.5 在冬季,温度较低,太阳辐射较弱,加上林冠层的影响,林内的直接蒸发很小,大多数是由植物蒸腾作用引起的水分蒸发,相比于林内,林外的大多数蒸发量是由水面和陆面蒸发带来的,因此潜在蒸发量较大。
猜你喜欢
农业工程学报(2022年12期)2022-09-09
作物杂志(2022年3期)2022-07-06
作物学报(2022年5期)2022-03-16
新疆农业科学(2021年11期)2021-12-23
绿色科技(2021年1期)2021-02-22
热带作物学报(2020年6期)2020-08-04
福建林业(2020年5期)2020-03-18
天津农业科学(2015年12期)2015-12-03
农民致富之友(2014年7期)2014-10-21
