
谷子TCP转录因子家族成员鉴定与生物信息学分析
2020-08-11 07:39白永宏赵赞延魏雄博王亚洁陈国梁
南方农业学报 2020年6期
白永宏 赵赞延 魏雄博 王亚洁 陈国梁


摘要:【目的】鑒定谷子TCP转录因子家族成员,并进行生物信息学及组织表达特性分析,为探究谷子TCP转录因子在植物生长发育、激素信号途径和生物胁迫中的生物学功能提供理论依据。【方法】通过HMMER构建隐马尔可夫模型,在谷子蛋白数据库中搜索含有TCP特征性结构域的TCP蛋白,对其理化性质、保守基序和系统发育进化进行分析,并对其基因结构、启动子顺式作用元件及组织表达特性进行分析。【结果】从谷子蛋白数据库中共鉴定出23个TCP转录因子(编号SiTCP1~SiTCP23),氨基酸数量为95~451个,分子量为9743.9~46502.6 Da,理论等电点(pI)为4.2682~11.8710,其编码基因在9条染色体上呈不均匀方式分布。23个谷子TCP蛋白被分为3个亚族,其中GroupⅠ和Group Ⅲ中各有10个TCP蛋白,GroupⅡ有3个TCP蛋白。谷子TCP蛋白保守基序的组成存在一定差异,但亲缘关系较近的蛋白具有相同的保守基序,部分谷子TCP蛋白含有特殊的基序;除SiTCP5、SiTCP20、SiTCP18和SiTCP22外,其余TCP蛋白均含有保守Motif 1和Motif 2。除SiTCP1、SiTCP6和SiTCP14基因仅包含1个内含子外,其余TCP基因均无内含子,且亲缘关系较近的基因具有相似结构。谷子TCP基因的启动子含有光应答元件、激素响应元件、应激响应元件及分生组织表达应答元件等。谷子TCP基因在不同组织中具有特异性,且大部分TCP基因在穗状花序和茎中高效表达。【结论】同亚族的谷子TCP转录因子家族成员具有相似的保守基序和基因结构,推测其在激素信号途径、应答非生物胁迫(干旱和低温)及生长发育中发挥重要作用。
关键词: 谷子(Setaria italic L.);TCP;转录因子;生物信息学;组织表达;系统发育进化
Abstract:【Objective】Gene family members of TCP transcription factorin foxtail millet were identified,their bioinformatics and expression in different tissues were analyzed. This would provide a theoretical basis for studying the function of foxtail millet TCP transcription factor in development,hormone signaling pathway and biotic stress. 【Method】By constructing a hidden Markov model by HMMER,the TCP proteins contained TCP domain were scanned in the foxtail millet protein database,its physicochemical property,conserved motifs and phylogenetic evolution were analyzed,and the gene structure,promoter region cis-acting elements and tissue expression specific of encoding gene were analyzed. 【Result】Twenty-three TCP transcription factors(number: SiTCP1-SiTCP23) were identified from foxtail millet protein database,the number of amino acids was 95-451,and the molecular weight was 9743.9-46502.6 Da. The theoretical isoelectric point(pI) was 4.2682-11.8710. The encoding genes were distributed unevenly on nine chromosomes. The 23 foxtail millet TCP transcription factors were divided into three subgroups,GroupⅠ and Group Ⅲ contained each 10 TCP proteins, and GroupⅡcontained 3 proteins. There were some differences in the composition of the conserved motifs infoxtail millet TCP proteins,but proteins with close genetic relationship had the same conserved motifs,some of which contained special motifs. Except SiTCP5,SiTCP20,SiTCP18,and SiTCP22,the remaining TCP proteins contained conservative Motif 1 and Motif 2. With the exception of SiTCP1,SiTCP6 and SiTCP14,which contain only one intron,all the other TCP genes did not contain introns,and the closely related genes had similar structures. The foxtail millet TCP gene promoter region contained light response elements, hormone response elements,stress response elements, meristem expression response elements. The TCP genes of foxtail millet was specific in different tissues and most TCP genes were highly expressed in spica and stem. 【Conclusion】The members of foxtail millet TCP transcription factor in the same subgroup have similar conserved motifs and gene structures,which are speculated to play important roles in hormone signaling pathways, response to abiotic stress (drought and low temperature), and growth and development.
Key words:Setaria italic L.; TCP; transcription factor; bioinformatic; tissue expression; phylogenetic evolution
0 引言
【研究意义】谷子(Setaria italic L.)原产于我国,俗称小米,是禾本科狗尾草属粮食作物,具有耐旱、耐贫瘠的特性(李顺国等,2018;李任建等,2020),其谷粒营养价值极高,富含维生素和其他微量元素,既有食用价值,又具有药用价值(王海岗等,2016)。Teosinte branched1/Cycloidea/Proliferating cell factor(TCP)转录因子在植物生长发育方面具有重要的作用,但其进化史尚不清楚(Liu et al.,2019),因此对谷子TCP转录因子进行生物信息学分析,可为探究TCP转录因子的系统发育及生物学功能提供科学依据,同时对培育优良谷子种质具有重要意义。【前人研究进展】TCP转录因子是一类植物特有的蛋白,具有由60个氨基酸组成的高度保守TCP结构域。研究起初,最早发现的TCP基因是玉米tb1(Teosinte branched1)基因(Doebley et al.,1997)、金草鱼cyc(Cycloidea)基因(Luo et al.,1999)和水稻pcfs(Proliferating cell factors)基因(Kosugi and Ohashi,1997;Cubas et al.,1999)。后续研究发现,TCP转录因子参与植物的多种生长发育过程,包括叶片发育(Palatnik et al.,2003;Efroni et al.,2008)、花的发育(Busch and Zachgo,2007;Nag et al.,2009)、种子萌发(Tatematsu et al.,2008)及激素信号途径(Schommer et al.,2008;Sugio et al.,2011)。在水稻中,OsTB1蛋白是影响植株侧枝发育的负调节转录因子(Takeda et al.,2003)。在拟南芥中,AtTCP2、AtTCP3、AtTCP4、AtTCP10和AtTCP24基因参与调控植物叶片形态发生(Palatnnik et al.,2003);AtTCP14基因在种子萌发前期的表达水平较高,正调节胚生长,说明其在种子萌发过程中发挥重要作用(Tatematsu et al.,2008);拟南芥植株过表达OsTCP19基因会导致其生长素响应基因3(IAA3)、脱落酸不敏感基因3(ABI3)和脱落酸不敏感基因4(ABI4)表达上调,但脂氧合酶基因(LOX2)表达下调,引起侧根形成减少等发育异常(Mukhopadhyay et al.,2015)。此外,TCP转录因子功能与激素途径密切相关。在拟南芥中,AtBRC1/TCP18与生长素通路发生相互作用以调控植物分支发育,而AtTCP8在控制开花方面具有重要作用(Wang et al.,2019)。在柳枝稷中,TCP转录因子PCF5是一种具有反式激活活性并控制叶片发育的核定位蛋白(Xie et al.,2017)。在玉米中,共鉴定出46个TCP基因,其中ZmTCP42基因在抗干旱胁迫中发挥正调控作用(Ding et al.,2019)。目前谷子全基因组已测序完成,为研究谷子功能基因提供强有力的数据支撑(Zhang et al.,2012;袁玺垒等,2018)。还有研究发现,突变小麦TaTCP9基因能使其穗粒长度增加(Bennetzen et al.,2012;Zhao et al.,2018);玉米中的大部分TCP基因均在茎和穗中表达(Chai et al.,2017)。【本研究切入点】目前,谷子TCP转录因子家族成员尚未进行系统鉴定,且其成员个数、系统进化及其编码基因结构等相关研究也鲜见报道。【拟解决的关键问题】通过HMMER构建隐马尔可夫模型,在谷子蛋白数据库中搜索含有TCP特征性结构域的TCP蛋白,对其理化性质、保守基序和系统发育进化进行分析,并对其编码基因进行基因结构、启动子顺式作用元件及组织表达特性分析,为探究谷子TCP转录因子在植物生长发育、激素信号途径和生物胁迫中的生物学功能提供理论依据。
1 材料与方法
1. 1 试验材料
从NCBI数据库(https://www.ncbi.nlm.nih.gov)中搜索获取谷子全基因组数据。从TAIR(http://www. arabiopsis.org/tools/bulk/sequences/index.jsp)中下载拟南芥TCP蛋白的氨基酸序列,从NCBI數据库中下载水稻TCP蛋白的氨基酸序列。
1. 2 谷子TCP转录因子家族成员鉴定
在Pfam(http://pfam.xfam.org/)中获得TCP蛋白特征性结构域(Pfam编号PF03634),通过HMMER构建隐马尔可夫模型,在谷子蛋白数据库中搜索含有TCP特征性结构域的蛋白,然后通过NCBI中的CDD (https://www.ncbi.nlm.nih.gov/cdd)检测是否含有TCP特征性结构域,剔除不含特征性结构域的蛋白,最终得到谷子TCP转录因子家族成员。利用ExPASy ProtParam对谷子TCP转录因子家族成员的理化性质进行预测。
1. 3 谷子TCP基因染色体定位及基因结构分析
在NCBI数据库(https://www.ncbi.nlm.nih.gov)中获得谷子TCP基因在染色体上的位置,然后使用MapInspect绘制基因定位图。利用GSDS(http://gsds1.cbi.pku.edu.cn/)进行基因结构分析。
1. 4 谷子TCP蛋白保守基序分析
使用MEME(http://meme-suite.org/tools/meme)对谷子TCP蛋白模体进行保守基序分析,保守基序长度设为6~100个氨基酸,保守基序个数设为10个。
1. 5 谷子TCP蛋白系统发育进化分析
选取模式植物拟南芥和水稻的TCP蛋白与谷子TCP蛋白进行氨基酸序列比对,并利用MEGA 7.0邻接法构建系统发育进化树。
1. 6 谷子TCP基因组织表达特性分析
谷子TCP转录因子基因在穗状花絮、根、茎和叶中的表达数据(FPKM)来源于SIFGD数据库(http://www.stucralbiology.cau.cn/SIFGD),通过MeV绘制谷子TCP转录因子基因的组织表达热图(李任建等,2020)。
1. 7 谷子TCP基因启动子顺式作用元件分析
从NCBI数据库下载谷子基因组数据和GFF文件,使用TBtools提取谷子TCP基因家族的启动子序列,利用PlantCare(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)在线网站分析启动子区域的顺式作用元件,并利用TBtools进行可视化。
2 结果与分析
2. 1 谷子TCP转录因子家族成员鉴定与染色体定位分析结果
从谷子蛋白数据库中共鉴定到23个TCP转录因子(编号SiTCP1~SiTCP23),其理化性质及编码基因染色体定位信息如表1所示。23个谷子TCP蛋白的氨基酸数量为95~451个,平均氨基酸数量为286个,分子量为9743.9~46502.6 Da;理论等电点(pI)为4.2682~11.8710,平均为8.3800,其中,有5个TCP蛋白的理论等电点小于7.0000(由酸性氨基酸组成),其余18个TCP蛋白的理论等电点均大于7.0000(由碱性氨基酸组成)。由表1和图1可知,23个谷子TCP基因在染色体上呈不均匀方式分布,Chr1~Chr9上分别有4、4、3、1、4、1、3、0和3个TCP基因,Chr4和Chr6上仅有1个TCP基因,占基因总数的4.8%。两个或多个旁系同源基因在染色体上的距离不超过100 kb,且同源性在80%以上被称为串联复制,在基因家族扩张和进化过程中发挥重要作用(Huang et al.,2012)。SiTCP9基因与SiTCP10基因相邻近,且同源性为90.48%,推测二者发生串联重复事件。
2. 2 谷子TCP转录因子的系统发育进化分析结果
为明确谷子TCP转录因子家族的进化情况,基于拟南芥、水稻和谷子的TCP蛋白氨基酸序列构建系统发育进化树,结果如图2所示。谷子的23个TCP蛋白被分为3个亚族:GroupⅠ、GroupⅡ和Group Ⅲ。其中,GroupⅠ和Group Ⅲ中各有10个TCP蛋白,分别占谷子TCP蛋白总数的43.48%;SiTCP7、SiTCP17和SiTCP22被划分为GroupⅡ,占谷子TCP蛋白总数的13.04%。SiTCP2、SiTCP3、SiTCP4、SiTCP8、SiTCP12、SiTCP16、SiTCP18、SiTCP19、SiTCP20和SiTCP23聚类在GroupⅠ;SiTCP7、SiTCP17和SiTCP22聚类在GroupⅡ,其中,SiTCP22和OsTB1转录因子在系统发育进化树上的亲缘关系最近,由于OsTB1是影响水稻侧枝的负调节转录因子(Takeda et al.,2003),故推测SiTCP22在谷子侧枝发育过程中具有同样的调节作用;SiTCP1、SiTCP5、SiTCP6、SiTCP9、SiTCP10、SiTCP11、SiTCP13、SiTCP14、SiTCP15和SiTCP21聚类在GroupⅢ。由于AtTCP2、AtTCP3、AtTCP4、AtTCP10和AtTCP24基因能参与调控植物叶片形态发生(Palatnnik et al.,2003),SiTCP15、SiTCP14和SiTCP9基因与AtTCP3和AtTCP24基因聚类在一起,故推测SiTCP9、SiTCP15和SiTCP14基因参与调控谷子叶片形态发生。
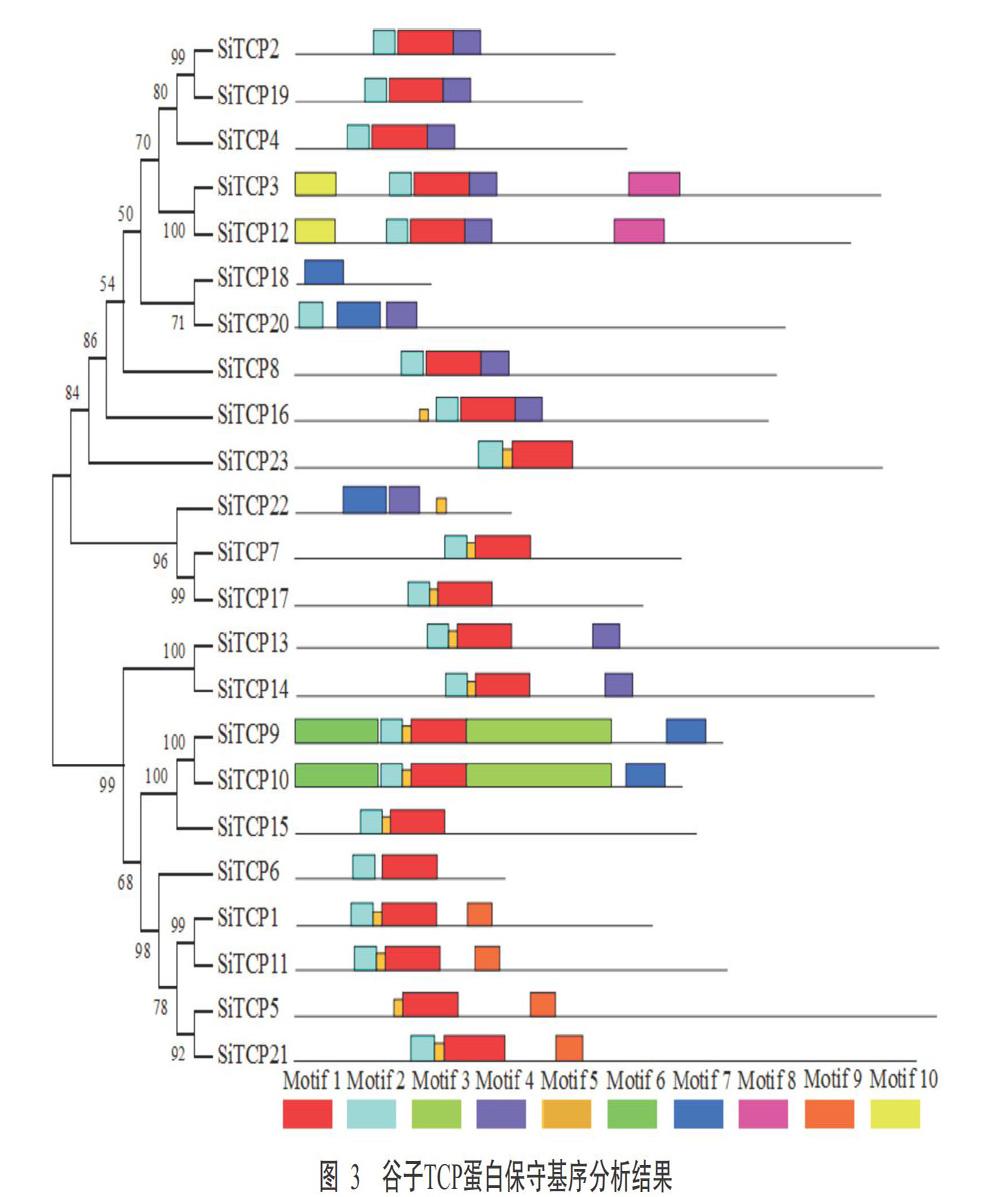
2. 3 谷子TCP蛋白保守基序分析结果
为分析谷子TCP蛋白结构多样性,对其保守基序进行分析,结果如图3所示。23个谷子TCP蛋白中,除SiTCP5不含Motif 2,SiTCP20不含Motif 1,SiTCP18和SiTCP22不含Motif 1和Motif 2外,其余TCP蛋白均含有Motif 1和Motif 2。谷子TCP蛋白保守基序的组成存在一定的差异,但亲缘关系较近的蛋白具有相同的保守基序,如SiTCP3与SiTCP12、SiTCP9與SiTCP10、SiTCP5与SiTCP21、SiTCP13与SiTCP14、SiTCP7与SiTCP17、SiTCP1与SiTCP11,以及SiTCP2、SiTCP4和SiTCP19三者所包含的基序相同。此外,部分谷子TCP蛋白还含有特殊的基序,如Group Ⅲ的SiTCP1、SiTCP5、SiTCP11和SiTCP21均含有Motif 9;GroupⅡ的SiTCP7和SiTCP17均含有Motif 5;GroupⅠ的SiTCP3和SiTCP12均含有Motif 3和Motif 8)。可见,同一亚族的TCP蛋白序列具有相似的保守基序。
2. 4 谷子TCP基因结构分析结果
为了明确谷子TCP基因结构多样性,通过GSDS绘制23个TCP基因内含子与外显子结构图(图4),结果显示,除SiTCP1、SiTCP6和SiTCP14基因仅包含1个内含子外,其余TCP基因均无内含子,且亲缘关系较近的基因具有相似结构,如SiTCP3与SiTCP12、SiTCP9与SiTCP10、SiTCP5与SiTCP21均具有相似的基因结构,SiTCP2、SiTCP4与SiTCP19三者间及SiTCP7、SiTCP17与SiTCP22三者间均具有相似的基因结构。可见,同一亚族的TCP基因具有相似的基因结构。
2. 5 谷子TCP基因启动子顺式作用元件分析结果
为探索谷子转录因子基因家族的功能和调控模式,使用PlantCARE数据库对每个TCP基因的启动子(2000 bp区域)进行顺式作用元件分析,结果(图5)发现多种类型的光响应顺式元件,包括G-box、GT1-motif、TATC-box和I-box等。此外,还发现激素响应顺式元件、应激响应元件及分生组织表达响应元件,其中,激素响应的顺式元件包括MeJA(茉莉酸响应元件)、ABA(脱落酸响应元件)、SA(水杨酸响应元件)、Auxin(生长素响应元件)和GARE(赤霉素响应元件),应激响应元件包括Drought(干旱响应元件)和LTR(低温响应元件)。综上所述,谷子TCP基因家族在激素信号途径、非生物胁迫(干旱和低温)响应及生长发育等方面具有重要作用。
2. 6 谷子TCP基因的组织特异性表达分析结果
利用MeV分析谷子TCP基因在不同组织中的表达情况,结果如图6所示。23个谷子TCP基因中,有泛表达基因,如SiTCP12基因在穗状花序、根、茎和叶中均高效表达,也有特异表达基因,如SiTCP5和SiTCP21基因仅在穗状花序中高效表达。此外,SiTCP9、SiTCP11、SiTCP12、SiTCP14和SiTCP15基因均在叶中表达较高,其中SiTCP9基因在根和穗状花絮中几乎不表达,在茎中表达量极低,SiTCP11基因在茎中的表达量也极低,SiTCP14基因在根中不表达,SiTCP15基因在茎中不表达。SiTCP10基因仅在叶中有微弱的表达,在穗状花絮、根和茎中均不表达。SiTCP1、SiTCP7、SiTCP8、SiTCP17和SiTCP22基因在根、茎、叶和穗状花序中均不表达。总体而言,大部分谷子TCP基因可在穗状花絮中表达。
3 讨论
谷子是我国北方主要的杂粮粮食作物,其耐旱、耐贫瘠能力比其他作物强,且营养价值极高,富含维生素和其他微量元素,除了食用价值高外,药用价值也较高(王海岗等,2016),已逐渐成为科研热点。植物的生长发育和抗逆机制较复杂,涉及许多转录因子的调控,其中,TCP转录因子参与植物的多种生长发育过程。目前,已从模式植物水稻和拟南芥中分别鉴定到22和23个TCP转录因子(Yao et al.,2007)。本研究从谷子中共鉴定出23个TCP转录因子,说明单子叶植物和双子叶植物中TCP转录因子数量无明显差异。此外,本研究发现,23个TCP蛋白保守基序的组成及其编码基因结构存在一定差异,但亲缘关系较近的转录因子具有相同的保守基序及相似的编码基因结构,且组织表达特性也较相似,故推测其具有相似的调控功能。SiTCP22与OsTB1聚类在一起,由于OsTB1是影响水稻侧枝的主要负调节蛋白(Takeda et al.,2003),故推测SiTCP22在调节侧枝发生方面具有相同的功能;AtTCP3、AtTCP24、SiTCP9、SiTCP15、SiTCP14与OsTCP1聚类在Group Ⅲ,亲缘关系较近,拟南芥TCP转录因子基因AtTCP3和AtTCP24参与调控植物叶片形态发生(Palatnnik et al.,2003),故推测SiTCP9、SiTCP15和SiTCP14基因参与调控植物叶片形态发生,具有相同的生物学功能。在拟南芥中,AtTCP14基因在种子萌发前期具有较高的表达水平,正调节胚生长(Tatematsu et al.,2008),AtTCP14与SiTCP17聚类在Group II,故推测SiTCP17在种子萌发过程中发挥相同的作用。SiTCP3与SiTCP20同聚在Group I,亲缘关系较近,二者具有相似的表达模式,均在茎中特异高表达。
本研究还发现,23个谷子TCP基因具有明显的组织表达特异性,但大部分TCP基因在穗状花序中表达。Chai等(2017)研究发现,玉米中的大部分TCP基因在茎和穗中表达。Zhao等(2018)研究发现,小麦中大部分TCP基因在穗苗和未成熟种子发育过程中均有表达。可见,TCP基因在进化过程中具有保守性。SiTCP9、SiTCP11、SiTCP12、SiTCP14和SiTCP15基因在叶中特异高效表达,推测这5个基因在叶片发育过程中具有重要作用,在今后的研究中可通过RT-PCR检测标记基因表达水平、光合速率、蒸腾速率和叶片相对水含量等来验证其是否参与叶片形态发生。SiTCP5、SiTCP13、SiTCP12、SiTCP20和SiTCP19基因在穗状花序中特异高表达,可能是穗状花序发育的潜在调控基因,今后的工作重心应通过RT-PCR、GUS染色等实验手段去验证其在穗状花序发育过程中可能的作用。
4 结论
同亚族的谷子TCP转录因子家族成員具有相似的保守基序和基因结构,推测其在激素信号途径、应答非生物胁迫(干旱和低温)及生长发育中发挥重要作用。
参考文献:
李任建,申哲源,李旭凯,韩渊怀,张宝俊. 2020. 谷子NBS-LRR类基因家族全基因组鉴定及表达分析[J]. 河南农业科学,49(2):34-43. [Li R J,Shen Z Y,Li X K,Han Y H,Zhang B J. 2020. Genome-wide identification and expression analysis of NBS-LRR gene family in Setaria italica[J]. Journal of Henan Agricultural Sciences,49(2):34-43.]
李顺国,刘斐,刘猛,刁现民. 2018. 新时期中国谷子产业发展技术需求与展望[J]. 农学学报,8(6):96-100. [Li S G,Liu F,Liu M, Diao X M. 2018. Technology need and prospect of foxtail millet industry development in China in the new era[J]. Journal of Agriculture,8(6):96-100.]
Liu M M,Wang M M ,Yang J,Wen J,Guo P C,Wu Y W,Ke Y Z,Li P F,Li J N,Du H. 2019. Evolutionary and comparative expression analyses of TCP transcription factor gene family in land plants[J]. Interational Journal of Molecular Sciences,20(14):3591. doi:10.3390/ijms2014 3591.
Luo D,Carpenter R,Copsey L,Vincent C,Clark J,Coen E. 1999. Control of organ asymmetry in flowers of Antirrhinum[J]. Cell,99(4):367-376.
Mukhopadhyay P,Tyagi A K,Tyagi A K. 2015. OsTCP19 influences developmental and abiotic stress signaling by modulating ABI4-mediated pathways[J]. Scientific Reports. doi:10.1038/srep09998.
Nag A,King S,Jack T. 2009. miR319a targeting of TCP4 is critical for petal growth and development in Arabidopsis[J]. Proceedings of the National Academy of Sciences of the United States of America,106(52):22534-22539.
Palatnik J F,Allen E,Wu X,Schommer C,Schwab R,Carrington J C,Weigel D. 2003. Control of leaf morphogenesis by microRNAs[J]. Nature,425(6955):257-263.
Schommer C,Palatnik J F,Aggarwal P,Chetelat A,Cubas P,Farmer E E,Nath U,Weigel D. 2008. Control of jasmonate biosynthesis and senescence by miR319 targets[J]. PLoS Biology,6(9):e230.
Sugio A,Kingdom H N,Maclean A M,Grieve V M,Hogenhout S A. 2011. Phytoplasma protein effector SPA11 enhances insect vector reproduction by manipulating plant development and defense hormone biosynthesis[J]. Proceedings of the National Academy of Sciences of the United States of America,108(48):E1254-E1263.
Takeda T,Suwa Y,Suzuki M,Kitano H,Ueguchi-Tanaka M,Ashikari M,Matsuoka M,Ueguchi C. 2003. The OsTB1 gene negatively regulates lateral branching in rice[J]. The Plant Journal,33(3):513-520.
Tatematsu K,Nakabayashi K,Kamiya Y,Nambara E. 2008. Transcription factor AtTCP14 regulates embryonic growth potential during seed germination in Arabidopsis thaliana[J]. The Plant Journal,53(1):42-52.
Wang X Y,Xu X T,Mo X W,Zhong L Y,Zhang J C,Mo B X,Kuai B K. 2019. Overexpression of TCP8 delays Arabidopsis flowering through a FLOWERING LOCUS C-dependent pathway[J]. BMC Plant Biology,19(1):534. doi:10.1186/s12870-019-2157-4.
Xie Q,Liu X,Zhang Y,Tang J,Yin D D,Fan B,Zhu L H,Han L B,Song G L,Li D Y. 2017. Identification and characterization of microRNA319a and its putative target gene,PvPCF5,in the bioenergy grass switchgrass(Panicum virgatum)[J]. Forntiers in Plant Science,8:396. doi: 10.3389/fpls.2017.00396.
Yao X,Ma H,Wang J,Zhang D. 2007. Genome-wide compara-tive analysis and expression pattern of TCP gene families in Arabidopsis thaliana and Oryza sativa[J]. Journal of Integrative Plant Biology,49:885-897.
Zhang G Y,Liu X,Quan Z W,Cheng S F,Xu X,Pan S K,Xie M,Zeng P,Yue Z,Wang W L,Tao Y,Bian C,Han C L,Xia Q J,Peng X H,Cao R,Yang X H,Zhan D L,Hu J C,Zhang Y X,Li H N,Li H,Li N,Wang J Y,Wang C C,Wang R Y,Guo T,Cai Y J,Liu C Z,Xiang H T,Shi Q X,Huang P,Chen Q C,Li Y R,Wang J,Zhao Z H,Wang J. 2012. Genome sequence of foxtail foxtail millet(Setaria italica) provides insights into grass evolution and biofuel potential[J]. Nature Biotechnology,30(6):549-554.
Zhao J M,Zhai Z W,Li Y N,Geng S F,Sun G Y,Guan J T,Jia M L,Wang F,Sun G L,Feng N,Kong X C,Chen L,Mao L,Li A L. 2018. Genome-wide identification and expression profiling of the TCP family genes in spike and grain development of wheat(Triticum aestivum L.)[J]. Forntiers in Plant Science,9:1282. doi:10.3389/fpls. 2018.01282.
(責任编辑 陈 燕)
猜你喜欢
中文信息(2017年2期)2017-04-13
江苏农业科学(2016年11期)2017-03-21
中学生数理化·中考版(2016年7期)2016-12-07
智能计算机与应用(2016年4期)2016-09-26
中学物理·高中(2016年8期)2016-08-08
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
恋爱婚姻家庭·养生版(2011年8期)2011-05-14
天津农业科学(2009年1期)2009-12-28
现代计算机(2009年9期)2009-12-02
