
几何形态测量法结合Micro CT扫描对两水系花斑裸鲤的形态分析
2020-08-07 01:23:58李晓慧汤永涛
水生生物学报 2020年4期
李晓慧 汤永涛 田 菲 赵 凯
(1.中国科学院西北高原生物研究所高原生物适应与进化重点实验室,青海省动物生态基因组学重点实验室,高原鱼类进化与功能基因组学实验室,西宁 810008;2.中国科学院大学,北京 100049;3.河南师范大学,新乡 453007)
花斑裸鲤(Gymnocypris eckloniHerzenstein),隶属鲤形目(Cypriniformes),鲤科(Cyprinidae),裂腹鱼亚科(Schizothoracinae),主要分布在我国柴达木水系格尔木河和黄河水系上游干支流[1]。花斑裸鲤物种最早是由俄国鱼类学家Herzenstein于1891年提出,他运用尽可能多的标本研究花斑裸鲤的形态性状,指出花斑裸鲤存在两个地理种群,主要分布于黄河水系上游及柴达木水系格尔木河,并将格尔木河的花斑裸鲤定为模式标本[2]。在随后的近一百年里,我国鱼类学家相继对花斑裸鲤的形态和分类进行过大量研究。武云飞等[1]对两水系花斑裸鲤的形态进行大量统计分析,结果显示除扎陵湖与鄂陵湖种群的鳃耙数偏高,呈现湖泊种所具有的地区性差异外,其余特征,包括口裂、背鳍距尾鳍基部距离、下颌形态等在两种群间均没有差异。此外,陈毅峰和曹文宣[3]经过大量形态学研究也得到了类似结果,认为两水系花斑裸鲤形态相似,是同一物种的不同地理种群。同样,赵铁桥[4]在对格尔木河、黄河水系上游与河西水系不同地理种群花斑裸鲤的形态学研究中提出,花斑裸鲤具有中间形态,其形态上的差异难以构成种级分化标准。以上学者基于形态学研究均认为黄河与格尔木河的花斑裸鲤是同一物种的不同地理种群。
然而,自从20世纪80年代随着分子生物学的发展,分子水平上对两水系裸鲤系统关系的研究却呈现出与形态学研究完全不一致的结果。赵凯等[5]利用线粒体标记Cytb基因对黄河水系、柴达木水系格尔木河花斑裸鲤的系统发育关系进行了研究,发现两水系花斑裸鲤经过地理隔离已经发生了独立进化。随后,Zhao等[6]在增加了祁连裸鲤的样品后,发现花斑裸鲤的两个地理种群没有形成单系群,而显示为相互独立起源的并系,再次肯定了上述结果的有效性。这一研究结果对花斑裸鲤的分类学及其两个地理种群的分类地位提出了不同的看法。
长期以来,对花斑裸鲤的形态学研究多采用传统形态测量方法,这一方法容易受主观因素的影响,也容易忽视一些肉眼难以观察到的性状[7,8],同时传统的形态分类标准,如口裂、鳃耙等多为环境适应性特征,易受环境影响而发生改变,不能客观反映物种的形态特征。随着科学技术的发展,现代几何形态测量法(Modern geometric morphometrics) 和Micro CT扫描技术(Micro computed tomography,微计算机断层扫描技术) 被广泛应用于物种形态鉴定。几何形态测量法是于20世纪90年代Bookstein[9]和Rohlf[10]以数学为基础所创,将统计学方法应用于形态分析,通过标志点(Landmark) 与滑动半标志点(Semi-landmark) 相互补充,对整体轮廓进行描绘[11],并最终通过多元统计分析对物种形态进行相似性比较和差异分析[12—14],进而定量判定形态特征的异同。经过近30年的发展,几何形态测量法已渗透到多个领域中[15—17],且在裂腹鱼类的形态鉴定中也已应用成熟[18—20]。Micro CT扫描是基于X射线对活体小动物进行断层扫描的一项技术,是利用不同组织对X射线吸收和透过率不同的原理实现的[21]。Micro CT扫描技术因其超高的分辨率更容易发现骨骼结构上的差异,现也广泛应用于小鼠骨密度研究、硬骨鱼类脊椎研究等多种小动物的骨骼形态研究方面[22—24]。
因此,为清楚阐释两水系裸鲤是否因为传统测量方法的局限性,使其分子与形态学研究不一致这一问题,本研究运用新型的几何形态测量法并结合Micro CT扫描技术,对两水系花斑裸鲤的头部形态及骨骼特征进行了定量分析,这对正确评价两水系花斑裸鲤真实的系统发育关系具有重要意义。
1 材料与方法
1.1 实验材料
本研究所用花斑裸鲤两个地理种群的样品分别采集于青藏高原东北部的黄河水系上游河段和柴达木水系格尔木河(图1),共采集116个样本(表1),同时作为本次实验对照组的青海湖裸鲤(Gymnocypris przewalskii przewalskii) 样品均采集于青海湖水系(图1),共19个样本(表1)。采集时间分别为2010年、2012年和2014年。采集时使用流刺网、地笼捕捞,并保存于95%酒精中,部分保存于福尔马林溶液中,现均存于中国科学院西北高原生物研究所高原鱼类进化与功能基因组学实验室和中国科学院西北高原生物研究所标本馆内。

图1 采样地点示意图Fig.1 Map of the head-water area of the Yellow River,Golmud River,and Qinghai Lake showing the sampling sites
1.2 图片信息获取
选取头部侧面轮廓和下颌轮廓对黄河与格尔木河花斑裸鲤(N=116) 进行形态比较。此外,补充了19个青海湖裸鲤样本作为外类群进行对照。具体方法如下。
所有样品均使用锡恩帝HX700数码相机拍摄,每个样品采集两张照片,分别为鱼头部左侧和腹部图片,拍摄时用镊子或大头针固定鱼的形态使其保持自然状态[19]。为减少系统误差,使用翻拍架固定相机位置,布置蓝色背景布,放置刻度尺,统一样品摆放位置,以确保样品与采集面保持平行,与相机高度距离相等。为减少拍摄时的人为误差与偶然误差,所有拍摄均由一人完成。
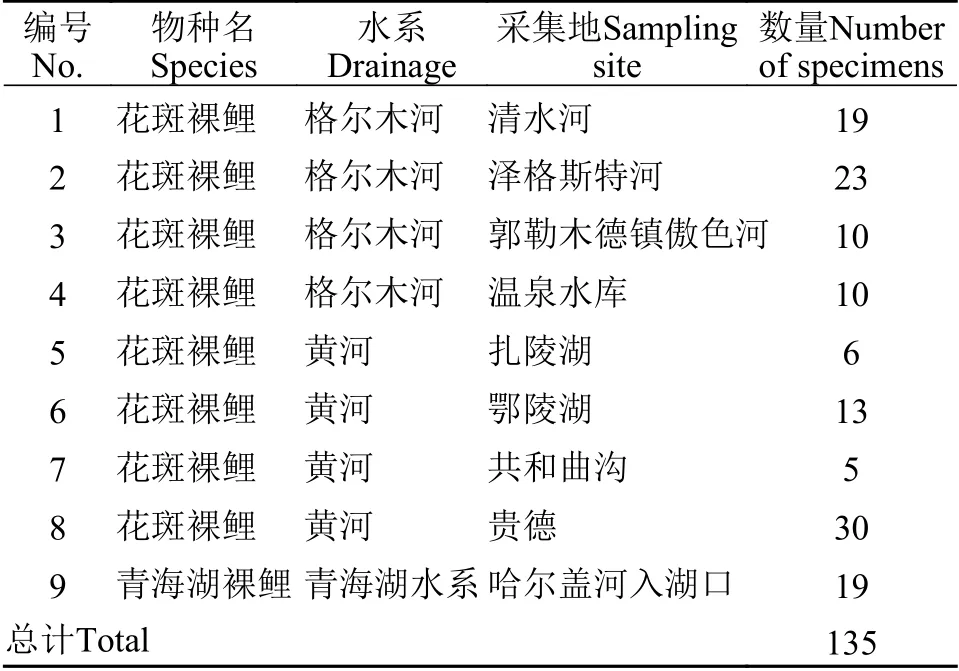
表 1 样品的采集水系、地点及数量Tab.1 Populations,numbers,river systems and sampling locations used in this study
1.3 标志点与半标志点选取
标志点需选取既有同源性又能准确反映物种形态信息的位点,根据类似的研究[19,20],并结合花斑裸鲤实际形态特点,本次实验选取了花斑裸鲤头部侧面5个标志点: ①头骨背缘末端,②吻上唇顶点,③吻下唇顶点,④口裂末端,⑤腹中线上腮盖条膜后缘。为使其能更加精确地描绘头部轮廓,结合文献采用滑动技术的方法选取了17个半标志点。半标志点的确定是在IMP软件包MakeFan8 (canisius.edu/~sheets/IMP%208.htm) 软件中实现的,首先确定同源性位点,然后以两者之间的连线做11条距离相等的垂线,其垂线与头部轮廓的交点即为半标志点(图2a)。花斑裸鲤下颌轮廓选取了2个标志点:①下颌骨上端,②下颌骨下端。下颌骨轮廓半标志点的确定同样在MakeFan8中实现,以确定的两个标志点作连线为直径画半圆,再以一组射线将其分成11份,则射线与下颌轮廓的交点即为半标志点(图2b)。随后利用TpsDig 2.04软件记录标志点与半标志点。
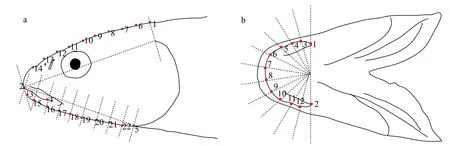
图2 花斑裸鲤标志点、半标志点的选取Fig.2 Landmarks and semi-landmarks of G. eckloni
1.4 图像数字化处理与统计分析
采用TPS系列软件对图片进行数字化处理[25]。利用TpsUtil (tpsutil.updatestar.com) 修改样本的ID,把本研究所涉及到的照片数据“矢量化”,最终生成扩展名为“.NTS”的二维坐标矩阵文件,用tpsDig 2.17 (life.bio.sunysb.edu/morph) 统一所有照片的比例尺以及按上述标志点对样本头部侧面和下颌进行标定。随后将生成的“.NTS”文件导入MorphoJ[26](flywings.org.uk/MorphoJ_page.htm) 软件中,进行New Procrustes fit消除非形变因素对数据分析的影响,即对样本进行普氏印叠(Generalized Procrustes Analysis,GPA)。GPA实际上是对数据标准化的过程,即通过对每个样品的标志点进行平移、缩放、旋转的过程来消除非形状因素产生的误差,使样本的几何形态得到精准的客观复原[9—13]。然后运用MorphoJ对普氏坐标数据进行主成分分析(Principal components analysis,PCA),用以分析不同水系花斑裸鲤几何形态上的差异;之后将二维坐标数据导入到PAST3.0[27]中进行辨别分析(Discriminant analysis,DLA),根据其不同地理种群二维坐标的特征值判断两水系花斑裸鲤的种群归属问题;进行典型变量分析(Canonical variable analysis,CVA),讨论花斑裸鲤两个地理种群间的差异,并计算马氏距离(Mahalanobis distance) 和普氏距离(Procrustes distance),来定量分析两种群间的形态差异量。
1.5 Micro CT扫描
本次实验使用QuantumGX Micro CT扫描系统,分别选取黄河水系花斑裸鲤21条,格尔木河花斑裸鲤22条进行骨骼扫描,分析两水系花斑裸鲤在骨骼方面是否存在未被发现的细微特征。参数设置: 电压70 kV,电流114 μA(该电流为此电压下的最大电流),视场86 mm,X-射线渗透Cu 0.1 mm,扫描模式采用High Resolution 4min,其余保持默认参数。
2 结果
2.1 两水系花斑裸鲤头部形状的主成分分析
几何形态学分析显示,花斑裸鲤的两个地理种群在主成分分析上并没有形成两个形态组,而是聚在一起 (图3)。头部轮廓的前两个PCA共解释了85.15%的变异量,其中PC1的贡献率为70.42%,PC2的贡献率为14.73%。在PCA二维分布散点图中,黄河与柴达木水系格尔木河的花斑裸鲤聚在一起,有很大程度的重叠,并没有明显的形态差异。沿PC1轴的正方向,两水系花斑裸鲤最明显的变化表现在1、2、3号特征点上,即口型位置发生了变化,逐渐表现为口亚下位的特点,符合花斑裸鲤的口型特征。
下颌轮廓的前两个PCA共解释了87.9%的变异量,其中PC1的贡献率为80.98%,PC2的贡献率为6.92%。在下颌轮廓的PCA散点图中,两水系花斑裸鲤与青海湖裸鲤都有一定程度的重叠,彼此之间均没有明显差异。下颌形态的网格图同样显示两水系花斑裸鲤在下颌形态上没有差异,均表现为吻钝圆,下颌弧形。
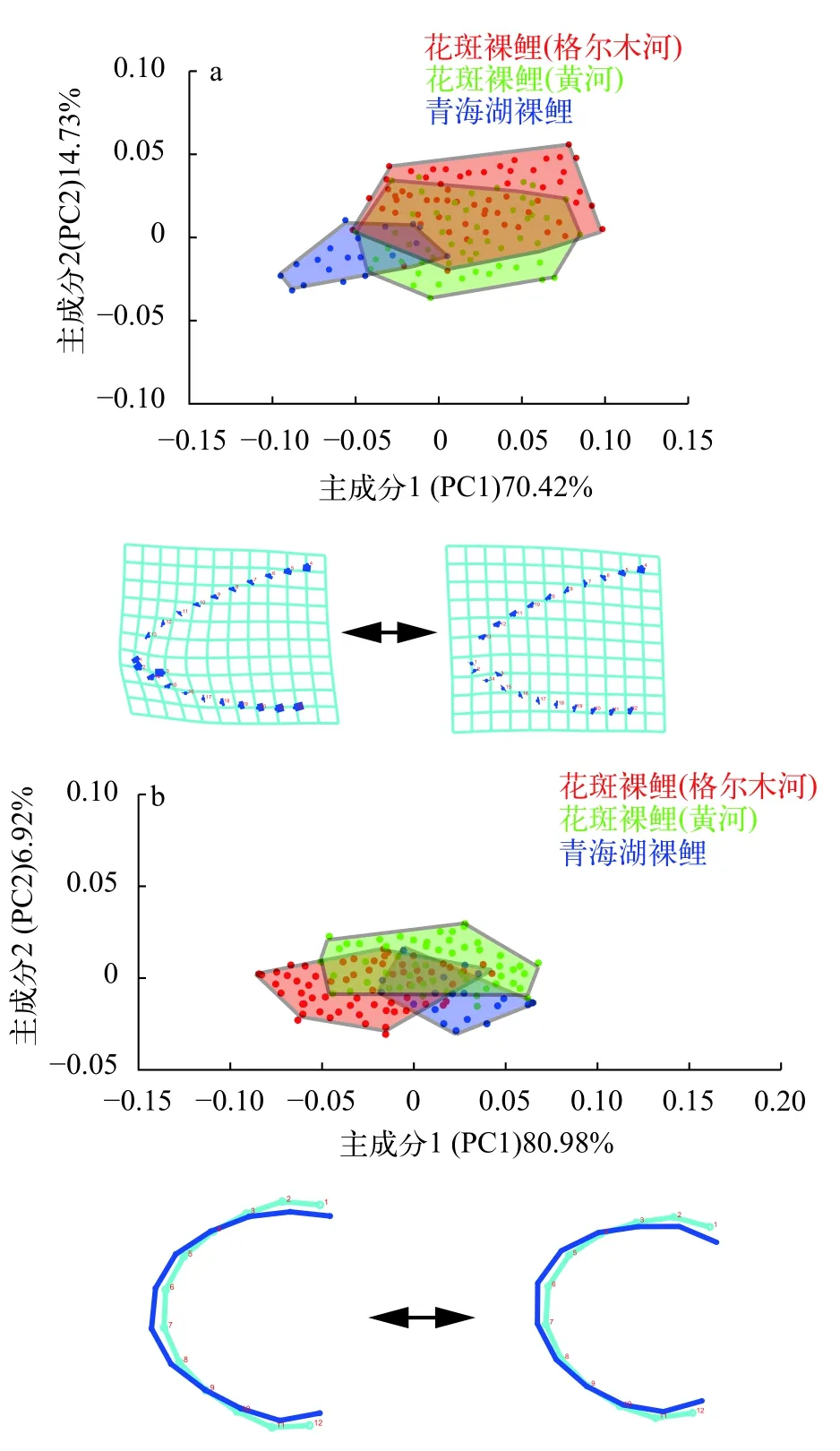
图3 黄河与格尔木河水系花斑裸鲤头部侧面轮廓(a) 与下颌轮廓(b) 的主成分分析Fig.3 Principal component analysis of the head profile (a) and mandibular profile (b) of the G. eckloni in Yellow River and Golmud River
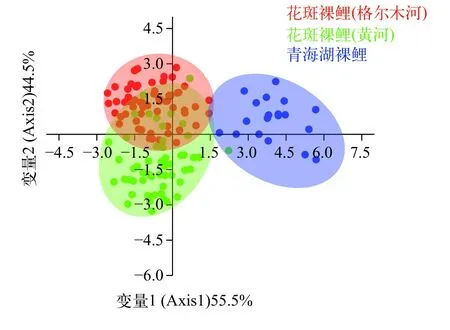
图4 花斑裸鲤头部侧面轮廓的判别分析Fig.4 The scatter diagram based on the LDA for the cephalic side contour of the G. eckloni
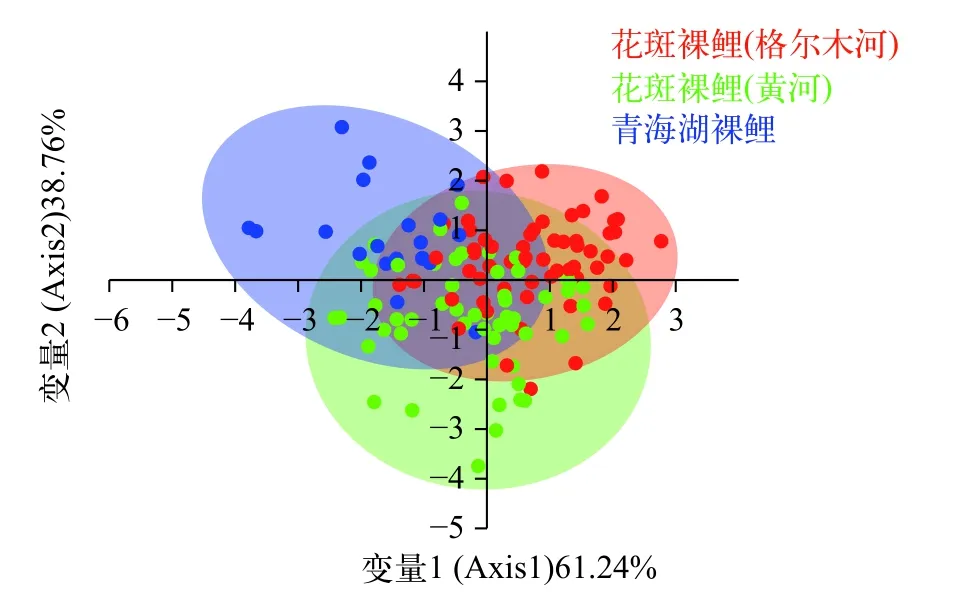
图5 花斑裸鲤下颌骨轮廓的判别分析Fig.5 The mandible morphological variations of the G. eckloni based on LDA
2.2 两水系花斑裸鲤头部形态的判别分析
除通过PCA分析计算研究样本几何形态上的差异外,本研究采用判别分析探讨两水系花斑裸鲤在形态上是否属于同一类别(图4和图5),结果显示: 在侧面轮廓的判别分析中,Axis1和Axis2分别占总体变异的55.5%和44.5%,判别分析的结果显示黄河水系与柴达木水系格尔木河的花斑裸鲤在侧面轮廓有很大程度的重叠,属于同一类别,与青海湖裸鲤存在明显差异(图4);图5为下颌骨形态的判别分析结果,其中Axis1和Axis2分别代表总变异的61.24%和38.76%,花斑裸鲤2个地理种群与青海湖裸鲤之间均有不同程度的重叠,三者在下颌形态轮廓特征上均无明显差异。判别分析结果与PCA结果基本一致。
2.3 两水系花斑裸鲤头部形态的典型变量分析
为量化花斑裸鲤两个地理种群间的差异,本文对两水系花斑裸鲤分别进行了头部形态的典型变量分析(图6),并计算马氏距离与普氏距离(表2—表5)。头部侧面轮廓的典型变量分析中CV1和CV2分别代表总体变异的59.42%和40.58%,在CV1轴上两水系花斑裸鲤没有差异,沿CV1轴正方向表现出口亚下位的特点,与PCA分析一致。在CV2轴上两水系花斑裸鲤有部分重叠,其差异主要表现在头部长宽比上,沿正值方向主要表现为头部形状略圆一些,正值处的特征在黄河水系花斑裸鲤种群中更明显一些。
下颌形态的典型变量分析中CV1代表总体变异量的75.74%,CV2代表总体变异量的24.26%。CV1展示的差异形态主要表现在下颌骨的形状上,CV1正值处格尔木水系花斑裸鲤的特征比较明显。两水系花斑裸鲤在CV2轴上存在重叠,没有明显差异。
此外通过计算花斑裸鲤两个地理种群与青海湖裸鲤头部形态的普氏距离与马氏距离,得到了与上述相似的结果,即三者之间没有明显的形态变异。特别是在头部侧面轮廓计算中相较于青海湖裸鲤,花斑裸鲤的两个地理种群间的变异较小,无明显形态特征。
2.4 Micro CT扫描结果
本研究分别对两水系花斑裸鲤的骨骼进行Micro CT扫描,并补充13条青海湖裸鲤的CT扫描结果作为对照,结果显示在背鳍支鳍骨插入椎骨间的位置上有所不同。背鳍支鳍骨插入椎骨的相对位置如表6所示: 在黄河水系花斑裸鲤中,背鳍第一支鳍骨位于11—12椎骨间,第二支鳍骨位于13—14椎骨间(图7a)的样本占到总研究样本的61.9%;在格尔木河水系花斑裸鲤的研究中,背鳍第一支鳍骨位于9—10椎骨间,第二支鳍骨位于11—12椎骨间(图7b)的样本占到总研究样本的59.1%;青海湖裸鲤在该特征上存在显著区别于两水系花斑裸鲤的形态特征,即第一支鳍骨与第二支鳍骨插入椎骨无间隔的性状(图7c),且占到研究样本的61.5%。此外三者均存在背鳍第一支鳍骨位于10—11椎骨间,第二支鳍骨位于12—13椎骨间的样品,但用卡方检验进行统计分析,结果显示**P<0.001,说明三者在背鳍支鳍骨插入椎骨间的相对位置上存在极显著差异。
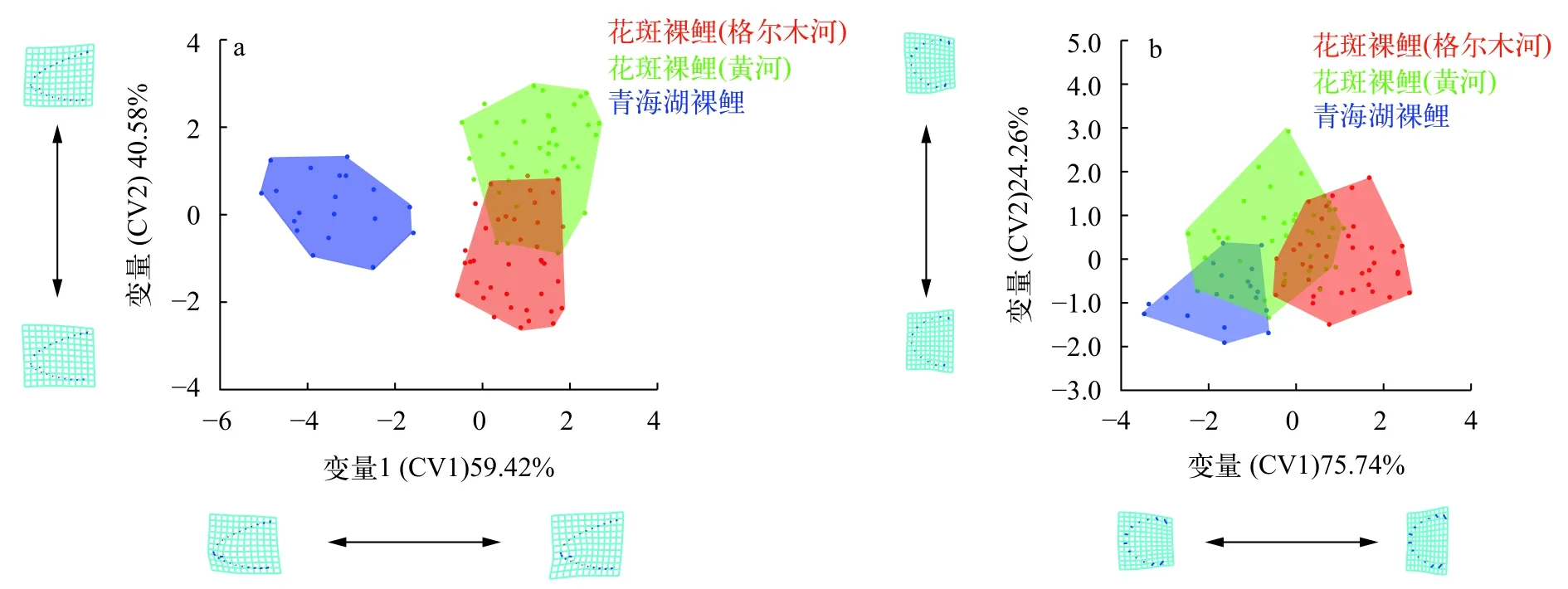
图6 黄河与格尔木河水系花斑裸鲤头部侧面轮廓(a) 与下颌轮廓(b) 的典型变量分析Fig.6 Canonical variable analysis of the head profile (a) and mandibular profile (b) of the G. eckloni in Yellow River and Golmud River
3 讨论
3.1 不同水系花斑裸鲤的形态分析
武云飞等[1]、曹文宣和陈毅峰[3]根据形态特征对黄河与柴达木水系格尔木河花斑裸鲤的形态学特征进行了研究,他们基于20多项形态分析得出了一致结论,即两水系花斑裸鲤的形态学特征没有差异,应属同一物种的不同地理种群。此外,赵铁桥在对134尾格尔木河、黄河上游水系及河西水系不同地理种群花斑裸鲤的形态学研究中发现,其下颌角质内缘、第一鳃弓鳃耙数等七项鉴别特征中均存在变异性大、性状不稳定等特点,难以作为种级鉴别特征,其形态上的细微差异不能作为物种分类标准[4]。本文通过采用更为先进的形态测量方法,对两水系花斑裸鲤头部形态及骨骼特征进行精确的定量分析,得到了比之前的研究更为丰富的结果。对花斑裸鲤两个地理种群头部轮廓定量分析的结果显示与上述前人的研究结果一致,即对头部轮廓特征点做PCA分析、判别分析和典型变量分析(图3—图6)均没有发现二者在头部轮廓形态上存在差异,马氏距离与普氏距离也同样显示两水系花斑裸鲤头部轮廓的变异量很小。然而值得关注的是,基于骨骼3D模型的比较,发现花斑裸鲤两个地理种群在“背鳍第一支鳍骨插入椎骨位置”这一性状上有着显著性差异(图7),该性状在虾虎鱼属中是一个重要的鉴别特征[28]。本研究是首次发现该性状在裂腹鱼类种群间存在显著性差异,但是否可以作为裂腹鱼类的一个鉴别特征还需今后广泛取样,在整个裂腹鱼亚科上进行实验验证。

表 2 花斑裸鲤不同种群头部侧面轮廓的马氏距离比较Tab.2 Comparison of Mahalanobis distance of head profile of the G. eckloni in Yellow River and Golmud River

表 3 花斑裸鲤不同种群头部侧面轮廓的普氏距离比较Tab.3 Comparison of Procrustes distance of head profile of the G. eckloni in Yellow River and Golmud River
3.2 不同水系花斑裸鲤形态演化的生态学机制
物种形态的演化深受生态压力的影响,特别是地质地貌的改变对物种的形态分化往往起着重要作用,鱼类口型下颌等环境适应性特征更是与其所处的流水环境和食物组成息息相关。高原河流生态位少,食物供给相对有限,在高原鱼类中发生形态趋同的现象极为普遍,如汤永涛等[20]在对高度特化等级裂腹鱼类基于简化基因组水平的研究中就发现裸鲤属与裸裂尻鱼属存在的大量形态趋同导致了分类学混乱。然而骨骼是构成脊椎动物的基本结构,在长期的形态演化中受环境影响较小,不仅起到支撑身体形态的作用,也是物种分类的重要依据[29]。早在20世纪90年代,孔晓瑜等[30]根据鱼类的骨骼特征,头骨、椎骨及附肢骨骼的差异将中国鳜亚科分为了三个属。在虾虎鱼的分类鉴定中,Birdsong等[31]将“背鳍支鳍骨插入椎骨间的相对位置”作为一个重要的分类依据,对200个属500种虾虎鱼进行了分类鉴定。

表 4 花斑裸鲤两个地理种群下颌轮廓的马氏距离比较Tab.4 Mahalanobis distance of mandibular contour between two geographic populations of G. eckloni

表 5 花斑裸鲤两个地理种群下颌轮廓的普氏距离比较Tab.5 Procrustes distance comparison of mandibular contour between two geographic populations of G. eckloni

表 6 不同水系样品背鳍支鳍骨与椎骨相对位置统计表Tab.6 The relative position of dorsal fin bone and vertebra in different drainage of sample
就本文而言,花斑裸鲤的两个地理种群(黄河和格尔木河)基于前人在形态学上的研究应属同一物种,但对两者分子水平上的研究显示他们是由不同祖先进化而来的并系。本研究所用的几何形态测量法,与传统测量方法相比,更能精确定量物种的形态特征,但采用几何形态测量法,分析两水系花斑裸鲤的环境适应性特征,头部形态及下颌骨轮廓,仍未发现两者存在显著差异,几何形态测量法支持了传统测量结果。但通过Micro CT扫描技术对花斑裸鲤两地理种群的骨骼结构进行分析,却观察到在“背鳍第一支鳍骨插入椎骨位置”这一更具分类学意义的特征上具有显著性差异。本文认为这很可能是由于两水系相似的生态环境造成的适应性形态趋同。青藏高原东部的黄河上游与柴达木水系的格尔木河地理相邻,同时发源于昆仑山脉[32]。作为花斑裸鲤模式生物产地的格尔木河,河水补给主要来源于高山融雪,食物组成相对有限,硅藻类是其水体生态系统的重要组成部分[33],底栖动物仅有寡毛类和钩虾。黄河上游同处高海拔,河水多来源于高山冰雪融水,且食物组成中同为硅藻类占据优势[1,34],兼有少量摇蚊幼虫等。花斑裸鲤的两个地理种群均为杂食性鱼类,以硅藻、摇蚊幼虫为主要食物,两个地理种群食性结构相似,在长期自然选择中很可能导致与食性相关的形态特征,如口裂、下颌形状等发生了趋同进化。此外,另一值得关注的结果是花斑裸鲤两个地理种群与青海湖裸鲤的“背鳍支鳍骨相对于椎骨位置”这一性状,在三者的种群内部均出现了分化,且均存在“背鳍第一支鳍骨插入第10—11椎骨间,第二支鳍骨插入12—13椎骨间”性状,这一结果又对两水系花斑裸鲤形态上的异同做出了另一种解释,可能暗示了三者在“背鳍支鳍骨插入椎骨相对位置”这一性状上尚未完全分化。青海湖裸鲤与花斑裸鲤属近缘种,且遗传分化时间短[5],很可能在青海湖裸鲤快速成种后,其骨骼特征分化速度较慢,还处于分化阶段。而花斑裸鲤一直以来被认为是广布种,具有广泛的形态多态性和不同水系的遗传分化[4,6],因而在青海湖裸鲤快速成种后,两水系花斑裸鲤在头部保留了原来的特征,支鳍骨相对于椎骨的位置却呈现一定的多态性。
总之,鱼类的表型性状深受环境压力的影响,在长期自然选择条件下,一些环境适应性特征往往会发生改变,而骨骼系统,如椎骨数、鳍条数等则受环境压力相对很小。因而采用几何形态测量法与Micro CT扫描技术对鱼类进行分类鉴定具有十分重要的意义,它比传统测量方法更能精确分析形态特征。近年来,现代几何形态测量法已在裂腹鱼上得到应用,但Micro CT扫描技术却是在花斑裸鲤为代表的裂腹鱼上的首次尝试,并发现了“背鳍支鳍骨相对于椎骨位置”这一性状在裂腹鱼亚科不同种群间存在显著差异的特性。综上所述,本研究丰富了花斑裸鲤不同地理种群间的形态学数据,并在形态学角度为两水系花斑裸鲤的分类地位及种群生态学提供了理论支持。但若要清楚阐释两水系花斑裸鲤的进化关系,真正厘清两者形态异同的生态学机制还需结合更广泛的分子数据,挖掘更多有意义的核基因或简化基因组数据进行深入研究,同时“背鳍支鳍骨相对于椎骨位置”这一性状是首次在裂腹鱼类上发现,Micro CT也是首次应用在裂腹鱼类的骨骼结构研究中,今后可作为一种模式在裂腹鱼类上进行广泛的实验验证,建立裂腹鱼类特征数据库,判断该性状是否可作为裂腹鱼类的鉴别特征。
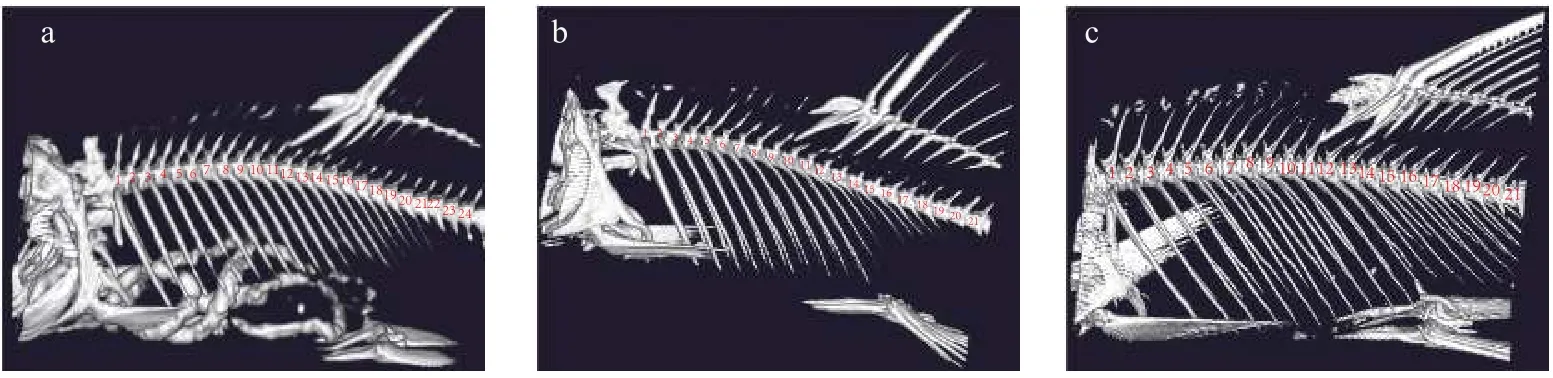
图7 不同水系样品背鳍支鳍骨与椎骨相对位置的部分展示图Fig.7 The relative position of the dorsal fin branch bone and vertebra in different drainage of sample
猜你喜欢
计算机测量与控制(2024年2期)2024-02-29 04:22:22
传感器与微系统(2021年3期)2021-03-26 04:35:38
第二课堂(课外活动版)(2021年6期)2021-01-18 18:55:58
浙江临床医学(2020年9期)2020-12-20 22:29:55
作文周刊·小学二年级版(2019年12期)2019-04-26 12:37:56
测控技术(2018年12期)2018-11-25 09:37:20
传感器与微系统(2018年7期)2018-08-29 00:44:24
广东海洋大学学报(2015年3期)2015-12-22 10:05:26
大灰狼(2015年5期)2015-07-25 20:57:36
计算机工程与设计(2014年9期)2014-12-23 01:16:00
