
硅氮互作改善优质稻植株生理生态特性
2020-08-06 14:57潘韬文陈俣蔡昆争
热带作物学报 2020年4期
潘韬文 陈俣 蔡昆争
摘 要:本研究以2个水稻品种‘桂农占和‘黄华占为供试材料,设置3个氮水平(120、180、240 kg/hm2分别表示为低氮、中氮和高氮),3个硅水平(0、225、450 kg/hm2分别表示为低硅、中硅和高硅),交叉互作,研究硅氮互作对水稻植株形态、物质积累、光合作用及碳氮代谢的影响。结果表明:硅氮互作有助于增加水稻的穗数、株高及生物量。中氮或中硅浓度能显著提高水稻叶片的光合作用,以‘桂农占效果更为明显,光合速率分别提高了19.21%和8.75%。在不同氮水平下,施用中等浓度硅可以提高‘桂农占和‘黄华占叶片的硝酸还原酶活性,分别比低硅处理增加46.82%和85.43%;在成熟期低氮水平下,2个水稻品种硝酸还原酶活性随硅浓度的增加而增加。对于叶片蔗糖转化酶活性,2个水稻品种均为抽穗期>分蘖期>成熟期,且各处理下,‘黄华占蔗糖转化酶活性基本大于‘桂农占。总体上来讲,低氮高硅,中氮中硅的硅氮配比组合,有助于促进水稻生长发育和物质代谢。
关键词:水稻;硅;氮;光合作用;碳氮代谢
中图分类号:S511; S181 文献标识码:A
Abstract: Two rice varieties ‘Guinongzhan and ‘Huanghuazhan were used as the material to study the effects of N and Si application on rice plant growth and physiological traits. Three levels of nitrogen 120, 180 and 240 kg/hm2, indicated by N1, N2 and N3, respectively, and three levels of silicon application 0, 225 and 450 kg/hm2, indicated by Si1, Si2 and Si3, respectively, were selected. The interaction of silicon and nitrogen could increase the panicle number, plant height and biomass of rice. The N2 and Si2 treatment significantly improved the photosynthesis of leaves, especially for ‘Guinongzhan. The photosynthetic rate increased by 19.21% and 8.75%, respectively. Under different N application levels, medium Si level (Si2) significantly increased the nitrate reductase (NR) activity of leaves. Compared with Si treatment, Si2 treatment increased NR by 46.82% and 85.43% for ‘Guinongzhan and ‘Huanghuazhan, respectively. NR activity increased with the increase of Si level at maturity stage under low N level. For sucrose invertase (SI) activity, both varieties showed the trend of different growth stage: heading stage > tillering stage > mature stage, and SI activity in ‘Huanghuazhan was higher than that in ‘Guinongzhan. The results suggested that low N and high Si, medium N and medium Si treatment could significantly promote rice growth and metabolism.
Keywords: rice; silicon; nitrogen; photosynthesis; carbon and nitrogen metabolism
DOI: 10.3969/j.issn.1000-2561.2020.04.009
水稻是發展中国家最重要的作物之一,是全世界35%人口的主要粮食来源[1],在农业生产和粮食安全方面具有举足轻重的地位[2]。氮素是植物生长必需的养分元素,但是长期过量施用氮肥会造成土壤氮素的大量盈余和流失[3-4]。其氮素一部分残留于土壤中,为后季作物吸收利用;另一部分氮素通过淋溶、氨挥发和反硝化作用而损失,进入环境并造成农业面源污染[5-6]。研究表明,合理施用氮肥,不仅可提高水稻的产量、品质和氮肥利用率,而且还可减少因过量施用氮肥而带来的环境污染[7-9]。此外,氮肥还可显著影响作物的生理生化特征,如花后光合速率和叶片中叶绿素含量[10]。
硅是土壤中第2大元素,但其有效性低,能被植物吸收、利用的很少[11]。硅自Sommer提出与水稻正常发育有关之后,很多研究也证实了硅在水稻生产上的必要性。硅能显著提高植株的茎秆坚硬度和抗倒伏能力[12-14],增加植物抗病性及光合作用效率[15-16],促进植物的健壮生长,提高作物产量和品质[17-18],以及增强植物对生物和非生物胁迫的抗性等[19-20]。郭彬等[18]研究发现,氮硅配施有利于水稻体内生理代谢的正常进行,明显地促进了水稻的生长。此外,氮硅配施效果优于氮硅单施,表明氮、硅配施对水稻生长发育有明显的交互作用。Fallah等[21]研究发现,在培养液(pH 5)中添加硅,无论氮的供应如何,都会增加组织中干物质的产生。适时施用氮和硅,可显著调节植物生长和产量性状[22]。本研究以南方推广面积较大的优质水稻为材料,研究硅氮互作后对不同水稻品种的植株形态、生物量、光合作用和碳氮代谢的影响,筛选合理的氮肥和硅肥施用量,从而为减氮、施硅的作物生产技术提供参考依据和实践指导。
1 材料与方法
1.1 材料
1.1.1 植物材料 华南地区优质稻‘桂农占和‘黄华占,由广东省农业科学院水稻研究所提供。
1.1.2 土壤及肥料 试验在华南农业大学试验农场进行。土壤主要理化性质如下:pH 6.48、全氮0.91 mg/g、速效磷31.70 mg/g、速效钾165.38 mg/g、土壤有效硅含167.88 mg/kg。试验所用硅肥为山西省曲沃县瑞丰硅肥厂生产的特种硅钙钾肥(SiO2≥20%)。
1.2 方法
1.2.1 试验设计 2011年进行早季和晚季大田试验,设3种供氮水平和供硅水平。3个供氮水平为:低氮(N1),中氮(N2),高氮(N3),分别为120、180、240 kg/hm2。3个供硅水平为:低硅(Si1),中硅(Si2),高硅(Si3),分别为0、225、450 kg/hm2。每个品种有9个处理,每个处理设3个重复,随机区组排列,共54个小区。小区面积为12 m2(长6 m,宽2 m),每个小区共300株秧苗,每小区设10行,株行距均为20 cm,每行插秧苗30株。
所有小区所施用的磷肥与钾肥相同,磷肥(过磷酸钙)施用量为375 kg/hm2,钾肥施用量为150 kg/hm2。磷肥作为基肥,插秧前整地时期1次性施用。钾肥基肥施用总量的40%,分蘖肥(插秧后15 d)追施30%,穗肥(插秧后30 d)追施30%。
由于2个品种的生育期大体一致,采取同时取样。2011年早季和晚季试验于分蘖期测定光合作用指标,抽穗期测定植株株高、穗数、生物量,分蘖期、抽穗期和成熟期测定不同处理下的叶片酶活性。由于早季和晚季实验结果一致,本研究仅展示2011年早季的实验结果。
1.2.2 植株生长参数的测定 于抽穗期测定植株的高度、穗数和生物量。
1.2.3 光合作用参数的测定 在晴天,选择水稻分蘖期主茎最上部完全展开叶中部用光合仪(美国Gene公司生产的LI-6400)分别测定各处理条件下第2片功能叶的光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Ts)、细胞间隙CO2浓度(Ci),记录数据。
1.2.4 碳氮代谢指标的测定 (1)硝酸还原酶(NR)活性的测定。采用活体法[23],进行显色测定并计算酶活性。
(2)蔗糖转化酶(INV)活性的测定。参考刘卫群等[24]的方法。取水稻新鲜叶片,除去叶脉,将叶片剪碎,准确称取0.5 g样品2份,在研钵中加入少量蒸馏水研磨成匀浆,然后定容到50 mL,摇匀,取10 mL 4 000 r/min离心15 min,分别吸取上清液2 mL加入2试管中,各加入pH 6.0磷酸缓冲液5 mL及10%蔗糖溶液1 mL,37 ℃水浴保温30 min,分别从2试管中吸取2 mL反应液于2刻度试管中,各加入1.5 mL 3,5-二硝基水杨酸试剂,沸水浴5 min,冷却定容,540 nm波长下测定OD值。所有结果均以鲜重计算。
1.3 数据处理
所有数据均采用Microsoft Excel软件进行计算,SPSS 16.0软件进行统计分析,Origin 9.0软件制图。
2 结果与分析
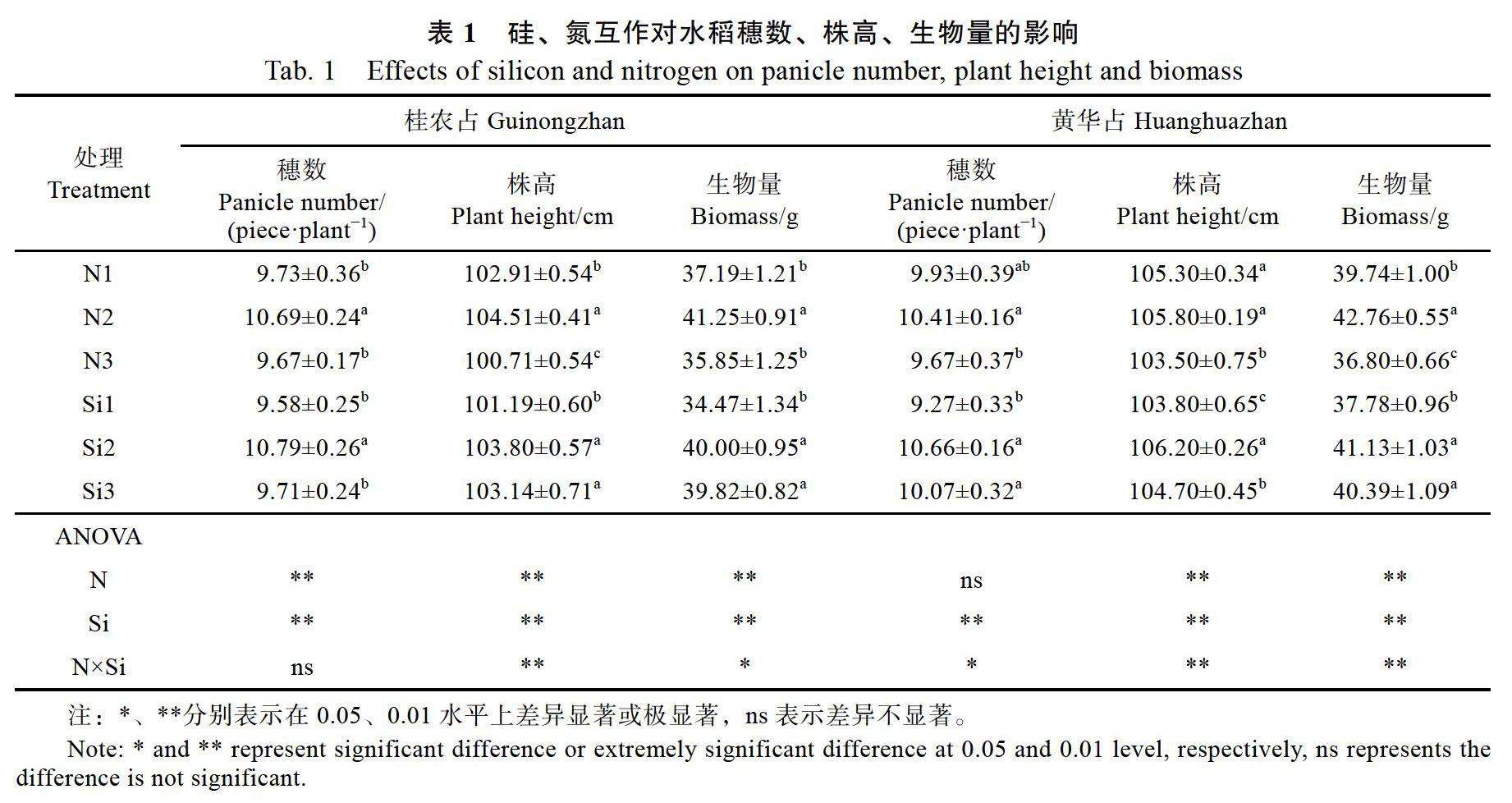
2.1 硅氮互作对水稻形态指标及干物质积累的影响
在抽穗期,‘桂农占在N2、Si2处理下显著增加穗数、株高和生物量(表1)。方差分析结果表明,除氮硅互作对穗数无显著差异外,对穗数、株高和生物量均存在显著影响。同样,‘黄华占在N2、Si2处理下穗数、株高和生物量达到最大(表1)。方差分析结果表明,氮对其穗数无显著差异外,其他处理对其穗数、株高和生物量均存在显著差异影响。‘桂农占在N2、Si2处理下的生物量比N3,Si1处理分别增加了13.1%和13.8%。
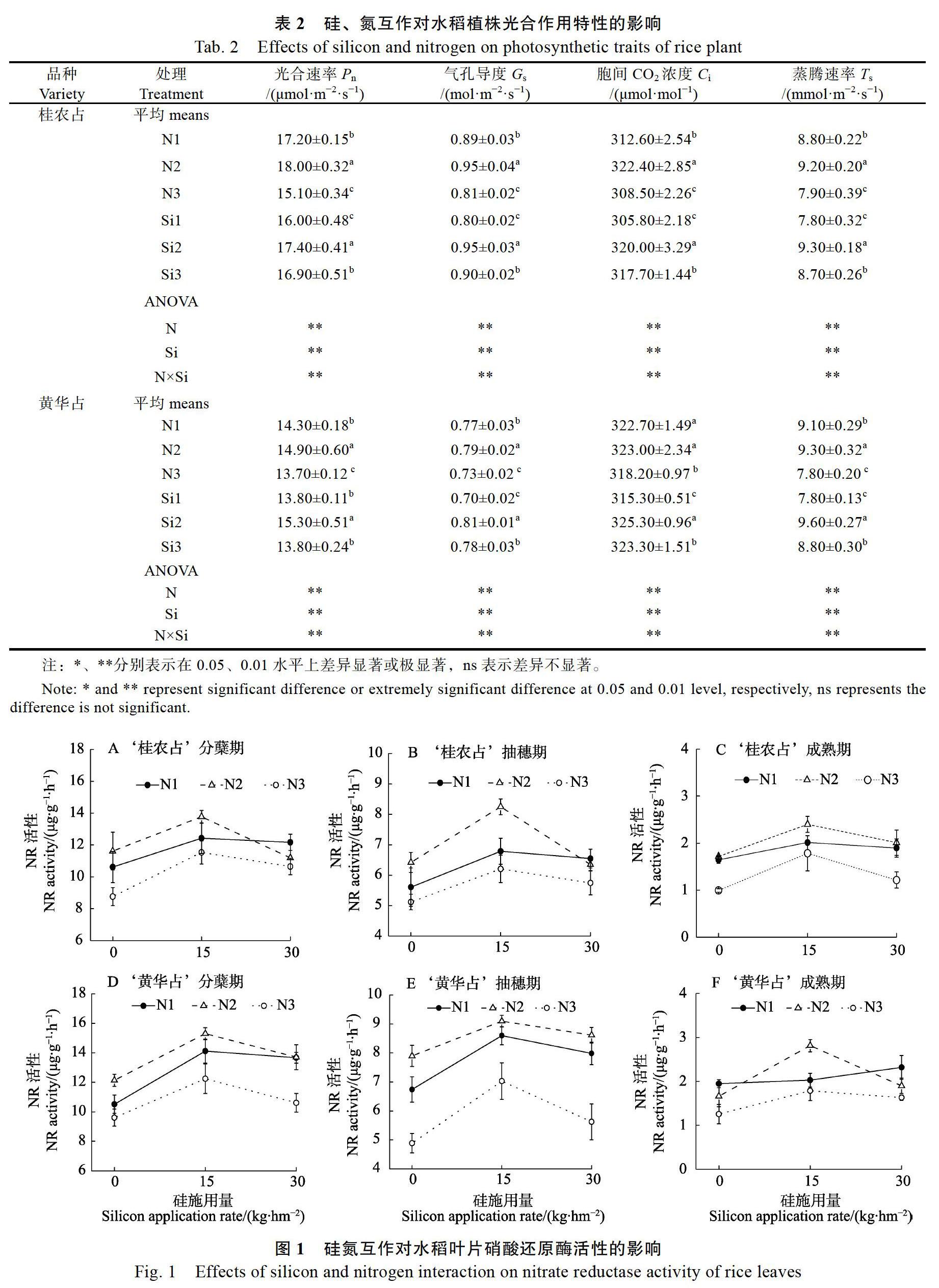
2.2 硅氮互作对水稻植株光合作用的影响
在水稻分蘖期,2个水稻品种叶片的光合速率、气孔导度、胞间CO2浓度、蒸腾速率规律变化一致,均在N2、Si2处理下其值最大(表2)。例如,‘桂农占在低、中、高3个氮浓度下,光合速率分别为17.2、18.0、15.1 μmol/(m2·s),‘黄华占在低、中、高3个硅浓度下分别为13.8、15.3、13.8 μmol/(m2·s)。同时,‘桂农占的光合速率以及气孔导度总体大于‘黄华占。方差分析结果表明,氮、硅、氮×硅互作对2个水稻品种的光合速率、气孔导度、胞间CO2浓度、蒸腾速率均有显著影响。
2.3 硅氮互作对水稻碳氮代谢的影响
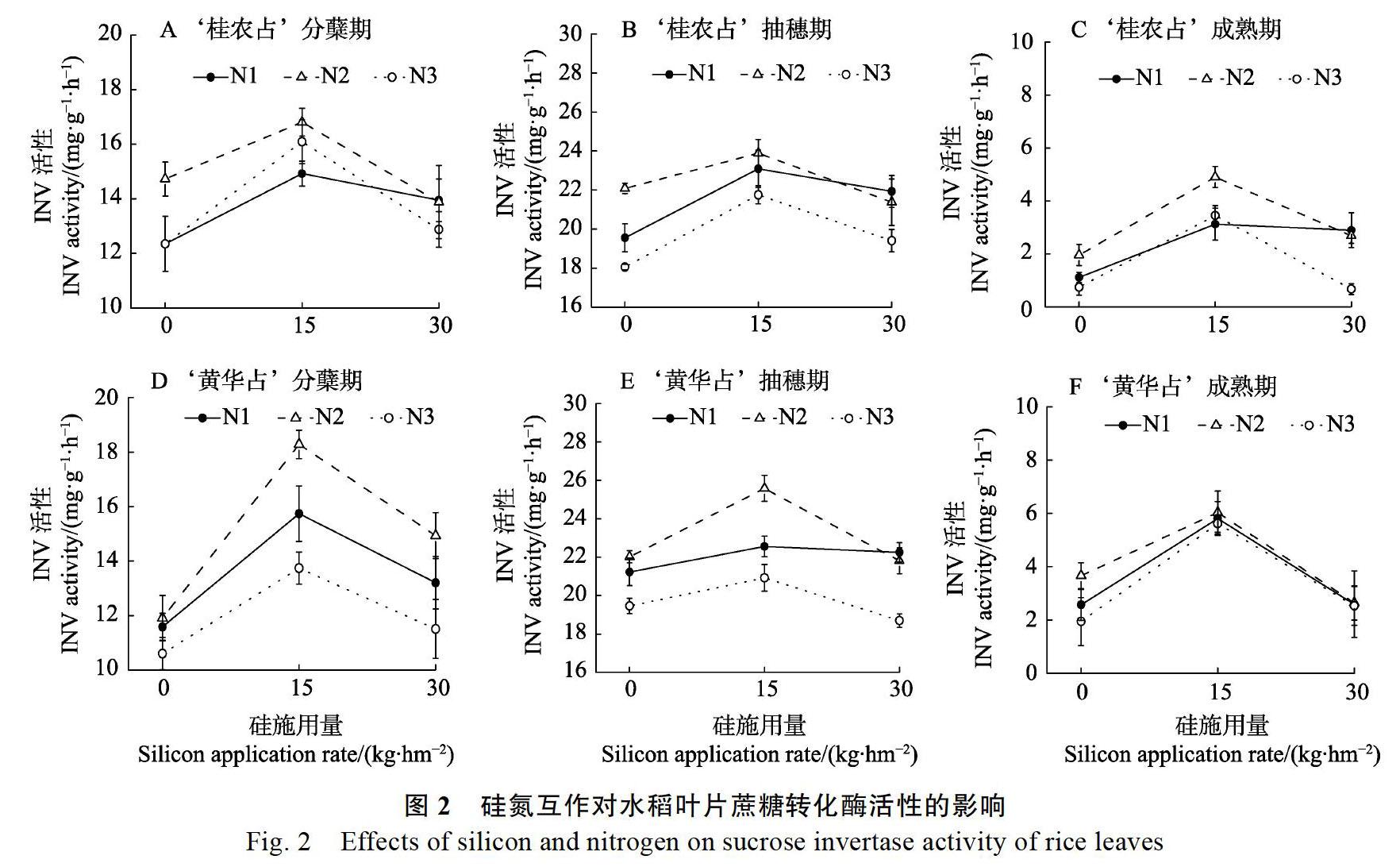
2.3.1 叶片硝酸还原酶(NR)活性 硝酸还原酶是一种氧化还原酶,可作为植物碳氮代谢的关键性酶[25],可催化硝酸离子还原成亚硝酸离子。硝酸还原酶可分为参与硝酸盐同化的同化型还原酶和催化以硝酸盐為活体氧化的最终电子受体的硝酸盐呼吸异化型(呼吸型)还原酶,是水稻植株氮代谢的限速酶,对水稻氮代谢的强弱起到关键作用。水稻叶片中的硝酸还原酶随着水稻生育期的进行也是在不断变化。
研究结果表明,在3个不同生育期,2个水稻品种基本上满足在N2-Si2处理下,NR活性最高(图1),可以看出氮和硅均是中等浓度时,水稻氮代谢最旺盛,为水稻的营养生长奠定基础,从而提高其产量等。‘桂农占和‘黄华占在N1-Si3处理下的NR活性比N1-Si1处理下分别提高了14.7%和29.8%,可见分蘖期低氮条件下,施用硅有利于水稻NR活性的提高,从而提高氮代谢能力。在成熟期,N2-Si2处理下,2个水稻品种的NR活性最高,2个水稻品种的叶片NR活性基本都降低到1~3个活力单位,这表明在水稻成熟期时,体内氮代谢已经逐渐减缓,甚至停止。
随着水稻生育期的进程,硝酸还原酶活性逐渐降低,‘桂农占分蘖期到抽穗期平均下降了45.1%,抽穗期到成熟期平均下降了72.2%,同时‘黄华占分蘖期到抽穗期平均下降了40.6%,抽穗期到成熟期平均下降了74.0%,因此水稻体内氮代谢也随着生育期的进程而逐渐减缓。
2.3.2 叶片蔗糖转化酶(INV)活性 正常条件下,叶片光合固定的CO2前期主要以淀粉、蔗糖的形式贮藏在叶绿体和细胞质中,水稻叶片在进行光合作用时,蔗糖必须经过转化酶分解为葡萄糖和果糖而进入三羧酸循环。所以转化酶在水稻体内对于蔗糖的吸收和利用有着至关重要的作用。
从水稻叶片蔗糖转化酶活性可以看出,2个水稻品种从分蘖期到抽穗期再到成熟期,INV活性均是先增强再迅速减弱的(图2)。并且在分蘖期和抽穗期的不同氮处理下,均是在中等硅浓度下INV活性最强。此外,‘黄华占在3个生育期的INV活性总体大于‘桂农占。
3 讨论
秦遂初等[26]研究表明,在高氮条件下,施硅可抑制水稻过量摄入氮,降低水稻高氮的负面影响。此外,植物体内硅含量的减少会导致谷物尤其是水稻的坏死、叶片光合效率的紊乱、生长迟缓和籽粒产量的降低[27]。本研究表明,施硅能显著增加抽穗期植株的生物量,促进作物生长,而以中等施用水平(Si2)的处理效果最为明显。光合作用所固定的二氧化碳在生长前期都以淀粉、蔗糖的形式贮藏在叶绿体和细胞质中[28]。硅处理可以提高叶肉细胞胞间CO2 浓度,增加气孔导度,从而提高水稻叶片光合效率[29]。本研究表明,在不同氮处理下,添加硅显著增加2个水稻品种的叶片光合作用,使水稻积累了足够的光合产物,为水稻生长发育奠定了基础。汪本福等[30]研究发现,在低氮条件下(施纯氮112.5 kg/hm2),增施硅肥显著提高了叶片光合速率,在高氮条件下(施纯氮225.0 kg/hm2),增施硅肥使得叶片光合速率先增后减,即叶绿素含量在一定范围内能增加光合速率,叶绿素SPAD超出一定范围后会出现过剩现象,光合速率不再增加。本研究中,2个水稻品种在低氮条件下,光合速率随硅添加量的增加而增加;在中、高氮条件下,中等浓度硅有利于光合速率的增加。
水稻的氮代谢开始于根细胞对土壤中硝酸盐和铵盐的吸收,通过特定转运蛋白吸收的氮素必须经过一系列代谢酶参与的反应与转化,才能被作物吸收与利用[31],硝酸还原酶是植物同化NO3过程中的关键酶,其活性高低与植物体内氮同化能力密切相关[32],对植物生长发育、产量形成和蛋白质产量都有重要影响。此外,氮的供给和代谢水平直接影响植物叶片衰老的程度[33]。徐国伟等[34]研究发现,水稻叶片氮代谢酶活性随着生育进程先提升后降低,而高氮降低了叶片氮代谢酶活性。本研究结果表明,分蘖期硝酸还原酶活性最强,随生育进程推进活性逐渐降低,在成熟期最弱,表明随着抽穗的来临,水稻氮代谢基本停止,开始碳代谢,促进源向库的转移。并且在不同硅浓度下,低、中浓度氮处理的硝酸还原酶活性均大于高氮处理。在水稻籽粒灌浆时期,氮代谢主要是功能叶分解蛋白质产生的氨基酸向籽粒运输而合成籽粒蛋白质的过程。水稻生育后期生理代谢变化最为剧烈的是氮代谢,早衰也是从体内氮代谢功能衰退开始的[35]。此外,关于氮、硅互作条件下,硝酸还原酶的具体推移时间,还应继续研究。
水稻的碳代谢,追其根本是糖代谢,水稻的有效穗、结实率和千粒重是构成产量的重要因素,这些因素与糖代谢有着一定的关系[36],然而蔗糖转化酶作为水稻碳代谢的关键性酶,对水稻碳代谢有很重要的影响。一般认为随着生育期的推进,水稻叶片转化酶活性会出现先升高再降低的趋势,这说明叶片对二氧化碳的固定作用逐渐减弱。本研究结果趋势与其大概一致。前人关于水稻碳氮代谢的研究主要集中在不同肥料水平和栽培模式下产量水平、根系活力、籽粒灌浆和品质等方面。以碳氮代谢为研究核心,本研究结果发现,减氮加硅处理的酶活性要高于减氮不加硅处理,表明硅可以合理调节水稻碳氮代谢,从而促进植物的生长发育。碳氮代谢是植物体内最基本的2大代谢过程,调节好两者关系对水稻碳、氮代谢平衡,提高水稻产量和氮素利用效率具有十分重要的意义[37]。
水稻施硅时,不仅要考虑土壤有效硅含量的高低,还要考虑施氮量和土壤供氮能力,有关这方面问题需要进一步深入研究。此外,氮肥和硅肥的互作效应已经得到很多学者的认同,但目前对氮硅的交互作用的应用仍远不能满足水稻可持续生产的需要,应进一步加强氮硅互作在水稻吸收、运输和体内代谢的机制及稻谷产量和品质等方面的研究。
参考文献
[1] Becker M, Asch F. Iron toxicity in rice-conditions and management concepts[J]. Journal of Plant Nutrition and Soil Science, 2005, 168(4): 558-573.
[2] 祝锦霞, 陈祝炉, 石媛媛, 等. 基于无人机和地面数字影像的水稻氮素營养诊断研究[J]. 浙江大学学报(农业与生命科学版), 2010, 36(1): 78-83.
[3] 侯云鹏, 韩立国, 孔丽丽, 等. 不同施氮水平下水稻的养分吸收、转运及土壤氮素平衡[J]. 植物营养与肥料学报, 2015, 21(4): 836-845.
[4] 刘晓伟, 王火焰, 朱德进, 等. 氮肥施用方式对水稻产量以及氮、磷、钾养分吸收利用的影响[J]. 南京农业大学学报, 2017, 40(2): 203-210.
[5] Sutton M A, Bleeker A. Environmental science: The shape of nitrogen to come[J]. Nature, 2013, 494(7438): 435-437.
[6] Zhang F, Chen X, Vitousek P. Chinese agriculture: An experiment for the world[J]. Nature, 2013, 497(7447): 33-35.
[7] 吴建富, 潘晓华, 石庆华. 施氮量对免耕抛栽稻产量及氮素吸收利用的影响[J]. 江西农业大学学报, 2011, 33(6): 1031-1036.
[8] 张维理, 田哲旭, 张 宁, 等. 我国北方农用氮肥造成地下水硝酸盐污染的调查[J]. 植物营养与肥料学报, 1995(2): 82-89.
[9] 唐启源, 邹应斌, 米湘成, 等. 不同施氮条件下超级杂交稻的产量形成特点与氮肥利用[J]. 杂交水稻, 2003(1): 47-51.
[10] 安 宁, 范明生, 张福锁. 水稻最佳作物管理技术的增产增效作用[J]. 植物营养与肥料学报, 2015, 21(4): 846-852.
[11] 田艳洪, 刘玉娥, 任 毅, 等. 硅氮肥配施生物菌粉对水稻生长及产量的影响[J]. 现代化农业, 2019(4): 19-21.
[12] Hossain M T, Mori R, Soga K, et al. Growth promotion and an increase in cell wall extensibility by silicon in rice and some other Poaceae seedlings[J]. Journal of Plant Research, 2002, 115(1): 23-27.
[13] 張玉龙, 王喜艳, 刘鸣达, 等. 植物硅素营养与土壤硅素肥力研究现状和展望[J]. 土壤通报, 2004, 35(6): 785-788.
[14] Ma J F, Yamaji N. Silicon uptake and accumulation in higher plants[J]. Trends in Plant Science, 2006, 11(8): 392-397.
[15] 薛高峰, 孙万春, 宋阿琳, 等. 硅对水稻生长、白叶枯病抗性及病程相关蛋白活性的影响[J]. 中国农业科学, 2010, 43(4): 690-697.
[16] 张国良, 戴其根, 霍中洋, 等. 外源硅对纹枯病菌(Rhizoctonia solani)侵染下水稻叶片光合功能的改善[J]. 生态学报, 2008, 28(10): 4881-4890.
[17] 丁文斌, 蒋建华, 陶吉平, 等. 禾丰氮钾硅镁肥在水稻上的应用效果初报[J]. 上海农业科技, 2010(2): 58-59.
[18] 郭 彬, 娄运生, 梁永超, 等. 氮硅肥配施对水稻生长、产量及土壤肥力的影响[J]. 生态学杂志, 2004(6): 33-36.
[19] 韩永强, 魏春光, 侯茂林. 硅对植物抗虫性的影响及其机制[J]. 生态学报, 2012, 32(3): 974-983.
[20] 王会方, 於朝广, 王 涛, 等. 硅缓解植物重金属毒害机理的研究进展[J]. 云南农业大学学报(自然科学版), 2016, 31(3): 528-535.
[21] Fallah A, Visperas R M, Alejar A A. The interactive effect of silicon and nitrogen on growth and spikelet filling in rice (Oryza sativa L.)[J]. Philippine Agricultural Scientist, 2004, 87(2): 174-176.
[22] Rukhsana J, Farooq A a, Fayaz A B, et al. Effect of nitrogen and silicon on growth and yield attributes of transplanted rice (Oryza sativa L.) under Kashmir conditions[J]. Journal of Pharmacognosy and Phytochemistry, 2018, 7(1): 328-332.
[23] 张志良, 瞿伟菁. 植物生理学实验指导[M]. 北京: 高等教育出版社, 2005: 41-43.
[24] 刘卫群, 陈良存, 甄焕菊, 等. 烟叶成熟过程中碳氮代谢关键酶对追施氮肥的响应[J]. 华北农学报, 2005, 23(3): 74-78.
[25] 黄树永, 陈良存. 烟草碳氮代谢研究进展[J]. 河南农业科学, 2005, 5(4): 8-11.
[26] 秦遂初, 马国瑞. 水稻生产中的硅[M]//孙 羲. 土壤养分、植物营养与合理施肥. 北京: 农业出版社, 1983.
[27] Shashidhar H E, Chandrashekhar N, Narayanaswamy C, et al. Calcium silicate as silicon source and its interaction with nitrogen in aerobic rice[C]//Silicon in Agriculture: 4th International Conference, South Africa, 2008: 93.
[28] 李潮海, 刘 奎, 连艳鲜. 玉米碳氮代谢研究进展[J]. 河南农业大学学报, 2000, 4(34): 318-323.
[29] 王 显, 张国良, 霍中洋, 等. 氮硅配施对水稻叶片光合作用和氮代谢酶活性的影响[J]. 扬州大学学报(农业与生命科学版), 2010, 31(3): 44-49.
[30] 汪本福, 黄金鹏, 赵 锋, 等. 硅氮配施对水稻光合特性、葉绿素荧光及产量的影响[J]. 中国稻米, 2016, 22(1): 30-34.
[31] Han M, Okamoto M, Beatty P H, et al. The genetics of nitrogen use efficiency in crop plants[J]. Annual Review of Genetics, 2015, 49(1): 269-289.
[32] 谷 岩, 胡文河, 徐百军, 等. 氮素营养水平对膜下滴灌玉米穗位叶光合及氮代谢酶活性的影响[J]. 生态学报, 2013, 33( 23): 7399-7407.
[33] Sui Y H, Gao J P, Shang Q Y. Characterization of nitrogen metabolism and photosynthesis in a stay-green rice cultivar[J]. Plant, Soil and Environment, 2019, 65(6): 283-289.
[34] 徐国伟, 陆大克, 刘聪杰, 等. 干湿交替灌溉和施氮量对水稻内源激素及氮素利用的影响[J]. 农业工程学报, 2018, 34(7): 137-146.
[35] 吴岳轩, 吴振球. 杂交水稻根系代谢活性与叶片衰老进程的相关研究[J]. 杂交水稻, 1992, 5(6): 36-39.
[36] 阳 剑, 时亚文, 李宙炜, 等. 水稻碳氮代谢研究进展[J]. 作物研究, 2011, 25(4): 383-387.
[37] 从夕汉, 施伏芝, 阮新民, 等. 施氮量对不同品种水稻氮素利用及碳氮代谢关键酶的影响[J]. 河南农业大学学报, 2019, 53(3): 325-330.
猜你喜欢
世界热带农业信息(2021年7期)2021-08-11
好孩子画报(2021年3期)2021-04-02
农民致富之友(2020年8期)2020-05-11
农民致富之友(2019年23期)2019-08-16
学苑创造·A版(2018年5期)2018-05-28
农产品市场周刊(2017年35期)2017-10-25
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
少儿科学周刊·儿童版(2015年10期)2015-11-07
中学生物学(2008年2期)2008-07-07
