
西藏湖泊放线菌的分离鉴定及抗菌活性测定
2020-08-04 03:02潘文娟林家富王小桃郭义东褚以文刘超兰
生物技术通报 2020年7期
潘文娟 林家富 王小桃 郭义东 褚以文 刘超兰
(成都大学四川抗菌素工业研究所 抗生素研究与再评价四川省重点实验室,成都 610052)
抗生素耐药性已成为全球关注的医学与社会问题,超级耐药菌严重威胁着人类的健康,新型抗生素的研发迫在眉睫[1]。众所周知,土壤放线菌一直是新抗生素研发的重要来源[2-5]。但随着研究深入,从土壤中筛选得到新放线菌及新次级代谢产物的概率呈显著下降趋势[6-7],迫使人们从新的生境中筛选新的放线菌资源,如海洋放线菌、动物共生放线菌、植物内生放线菌、极端环境放线菌等[8-12]。而湖泊生境中的放线菌资源开发至今未得到充分重视,尤其是位于高海拔西藏湖泊的放线菌资源研究仍处于空白。
西藏自治区是我国湖泊最多的地区,这些湖泊分为内流湖和外流湖,主要包括造山运动在地层断裂处积水而形成的断层湖、冰川形成的冰蚀湖、泥石流使河道堵塞形成的堰塞湖等。西藏湖泊形成原因比较特别,造就了其独特的生物多样性,因此,研究西藏湖泊中的放线菌资源及其次级代谢产物对于发现新抗生素先导化合物具有重要意义。我们在西藏湖泊微生物资源调查过程中,对采集于7个湖泊的沉积物样品进行了放线菌的分离鉴定及抗菌活性测定,本文报道获得的初步研究结果。
1 材料与方法
1.1 材料
1.1.1 沉积物样品 菌株分离用沉积物样品采集自西藏7个湖泊,包括左用措、纳木措、昆仲措、戈芒措、芦布措、昂仁金措、吉力湖。采集点位于湖中心区域,用淤泥分层采集装置采集表层沉积物(1-10 cm),样品存放于无菌试管中,放置4℃保存。采样信息见表1。
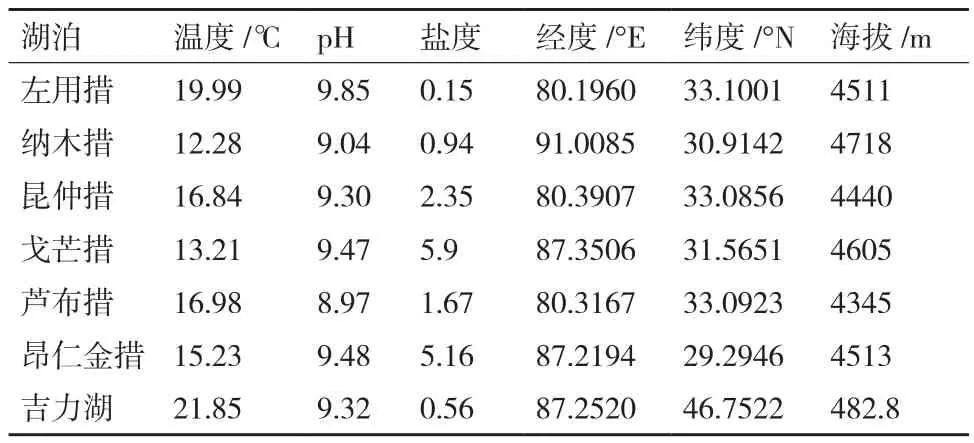
表1 西藏7个湖泊采样信息
1.1.2 培养基及溶液 (1)分离培养基:ISP2培养基[13]、ISP3 培养基[13]、SCA 培养基[14]、高氏一号培养基[15],各培养基中均含30 mg/mL萘啶酮酸和放线菌酮。(2)纯化培养基:ISP2培养基。(3)发酵培养基:葡萄糖10 g,糊精25 g,燕麦粉20 g,棉籽饼粉10 g,鱼粉5 g,糖蜜5 g,干酵母2 g,CaCO33 g,蒸馏水1 000 mL,pH 7.0-7.2,121℃高压灭菌15 min。(4)体外抗菌活性筛选用培养基:LB 细菌培养基[15],PDA 真菌培养基[15]。(5)NGM线 虫 培 养 基[16]。(6)M9缓 冲 液:Na2HPO46 g,KH2PO43 g,NaCl 5 g,MgSO4·7H2O 0.25 g,蒸馏水1 000 mL,121℃高压灭菌15 min,宜现用现配。
1.1.3 线虫 野生型秀丽隐杆线虫N2由加拿大戴尔豪斯大学Balakrishnan Prithiviraj博士惠赠,饲养于NGM线虫培养基中。
1.1.4 检定菌 金黄色葡萄球菌(Staphylococcus aureusSIIA1005)、大肠杆菌(Escherichia coliSIIA 1006)、白色念珠菌(Candida albicansSIIA 2284)、枯草芽孢杆菌(Bacillus subtilisSIIA 1007)、绿脓杆菌(Pseudomonas aeruginosaSIIA 2261)由中国医药集团总公司四川抗菌素工业研究所菌种保藏中心提供。
1.1.5 主要试剂和仪器 引物27F/1492R购自生工生物工程(上海)股份有限公司;PCR mix kit购自北京擎科新业生物技术有限公司;细菌基因组DNA提取试剂盒购自天根生化科技(北京)有限公司;万古霉素、庆大霉素、5-氟-2-脱氧尿嘧啶购自上海源叶生物科技有限公司;放线菌酮购自卡迈舒(上海)生物科技有限公司,萘啶酮酸购自阿法埃莎(天津)化学有限公司。使用仪器如下:超净工作台SW-CJ-1FD(苏州安泰空气技术有限公司),恒温培养箱HWS-150(北京中兴伟业仪器有限公司),振荡式摇床THZ-92A(上海博迅实业有限公司医疗设备厂),高速离心机H1650(湖南湘仪实验室仪器开发有限公司),PCR扩增仪T100(Bio-Rad,美国),凝胶成像仪Gel DocTMXR+(Bio-Rad,美国)、体视显微镜SMZ-168(杭州欧尔柏维科技有限公司)、96孔板(Corning,美国)。
1.2 方法
1.2.1 菌株分离 将沉积物样品自然干燥7 d后,称取样品2 g于60℃再干燥1 h后研磨过80目筛。将1 g干燥样品加入装有9 mL无菌蒸馏水的试管中,用涡旋振荡器振荡均匀,吸取1 mL土壤悬液至9 mL无菌蒸馏水中,进行10倍梯度稀释,使其终浓度为 10-2、10-3、10-4、10-5。吸取 0.2 mL 稀释液分别涂布于分离培养基上,于28℃培养30 d,从第7天开始,每天挑取新出现的菌落至纯化培养基上进行纯化,纯化后的菌株用30%甘油保藏于-80℃冰箱。
1.2.2 菌株分类鉴定及系统发育树的构建 采用细菌基因组DNA提取试剂盒提取基因组DNA,采用16S上游引物27F(5'-AGAGTTTGATCCTGGCTCAG-3')、下游引物1492R(5'- TACGGCTACCTTGTTACGACTT-3')进行PCR扩增。PCR扩增体系(25 μL):PCR mix 12.5 μL,ddH2O 8.5 μL,DNA 模板 2 μL,引物各 1 μL,扩增程序为:95℃ 5 min,94℃ 45 s,56℃ 45 s,72℃ 2 min,34个循环,4℃无限延伸。扩增完成后用1%琼脂糖凝胶电泳检测PCR反应产物。将结果为阳性的PCR反应产物送至生工生物工程(上海)股份有限公司进行测序。利用NCBI数据库中BLAST程序对测序结果进行比对分析,选择GenBank中与之同源性较高的模式菌株的16S rRNA基因序列,分析菌株与参比菌株间的序列相似度,利用MEGA6.0[17]软件,选择Tamura-Nei模型,构建菌株Maximum Likelihood系统发育树。
1.2.3 菌株发酵及粗提液制备 将菌株接种于ISP2斜面培养基中于28℃生长10-14 d后,接种到发酵培养基中,于28℃、220 r/min发酵7 d。取20 mL发酵液加入20 mL乙酸乙酯进行萃取,吸取乙酸乙酯层10 mL,旋转蒸发干燥,加入2 mL甲醇溶解后,备用。
1.2.4 体外抗菌活性筛选 采用纸片扩散法进行抗菌活性检测。吸取60 μL发酵粗提液至直径为6 mm的圆形灭菌滤纸片上,待甲醇挥发后,贴于含有检定菌的培养基上,37℃培养24 h,然后观察并记录抑菌圈大小,将抑菌圈直径大于1 cm作为阳性结果。
1.2.5 体内抗菌活性筛选 (1)线虫培养:线虫按照国际标准流程进行培养[18]。(2)线虫同期化:挑取处于产卵期的成年线虫于新的NGM平板,于15℃恒温培养箱培养12 h,待产卵后将成虫挑出,于20℃恒温培养箱培养48 h使虫卵孵化并成长至L4期线虫。(3)菌悬液制备:挑取金黄色葡萄球菌和大肠杆菌菌落于5 mL的LB液体培养基中,37℃、200 r/min振荡培养12 h,备用。(4)线虫抗感染实验:吸取金黄色葡萄球菌和大肠杆菌菌液100 μL接种至NGM培养基(含50 μg/mL 5-氟-2-脱氧尿嘧啶,用于抑制线虫产卵),37℃培养24 h。挑取已同期化的L4期线虫,于M9缓冲液中反复清洗干净后备用。将线虫分别挑取到含有金黄色葡萄球菌和大肠杆菌的平板上,感染时间为14-16 h。96孔板中每孔加入20条已感染的L4期线虫和190 μL M9缓冲液,其中阳性对照组加入10 μL抗生素(万古霉素、庆大霉素,终浓度为50 μg/mL),阴性对照组加入10 μL甲醇,空白对照组加入10 μL M9缓冲液,样品组加入10 μL发酵粗提液,每个发酵粗提液做3个复孔,并于培养箱中20℃静置培养。(5)线虫观察:每隔2 h观察1次,观察时间为20 h。正常线虫保持着正弦S形,并活跃地移动[19],而死线虫呈笔直状。记录存活和死亡线虫数量,计算存活率,线虫存活率(%)=线虫存活数/线虫总数×100%。以存活率>50%为阳性结果。
2 结果
2.1 菌株分离
从西藏7个湖泊沉积物样品中分离得到73株放线菌(图1),其中分离自芦布措湖的菌株数最多,共23株,占比31.5%;分离自昂仁金措湖的菌株数最少,为5株,占比6.8%。

图1 西藏7个湖泊沉积物样品分离菌株数
采用不同分离培养基所获菌株数有差异,ISP2、ISP3、SCA、高氏一号培养基分离菌株数分别为30株、25株、8株和10株,其中ISP2和ISP3培养基是分离西藏湖泊沉积物中放线菌的适用培养基。
2.2 菌株鉴定及系统发育树构建
根据16S rRNA基因序列,利用Maximum Likelihood和Tamura-Nei模型构建系统发育树(图2),结果表明,73株放线菌分属于5个科8个属,以链霉菌科为主,其次为小单胞菌科、诺卡菌科、链孢子囊科、拟诺卡菌科等。链霉菌属(Streptomyces)为优势属,共59株,占80.8%,小单孢菌属(Micromonospora)次之,共7株;其他属包括诺卡氏菌属(Nocardia,1株)、北里孢菌属(Kitasatospora,1株)、小双孢菌属(Microbispora,1株)、野野村菌属(Nonomuraea,1株)、游动放线菌属(Actinoplanes,1株)、拟诺卡氏菌属(Nocardiopsis,2株)。
根据Kim[20]提出的新种16S rRNA临界值98.65%,分离的菌株中存在潜在新种3株,分别为菌株PF1(Streptomyces pratensisch24T,相似度为98.4%)、 菌 株 PF26(Streptomyces pseudovenezuelaeDSM 40212T, 相 似 度 为 98.25%)、 菌 株 PF188(Nonomuraea guangzhouensisNEAU-ZJ3T,相似度为98.13%)。
不同湖泊所分离到的放线菌种类具有明显差异。各湖泊所分离菌株的属水平相对丰度(图3)表明,Streptomyces是各湖泊的优势菌属。而在芦布措湖的沉积物样品中发现最多稀有放线菌属,包括Kitasatospora、Microbispora、Nonomuraea、Actinoplanes,在昆仲措湖沉积物样品中还分离到Nocardiopsis。

图2 基于16S rRNA基因序列构建湖泊放线菌的系统发育树
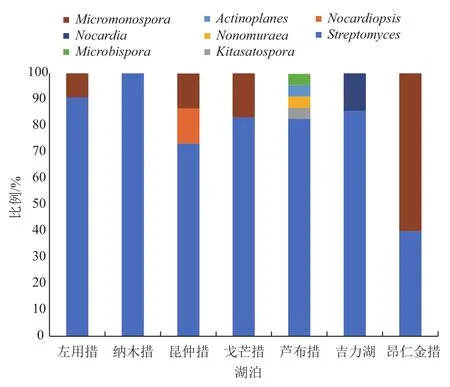
图3 西藏7个湖泊沉积物样品的放线菌属相对丰度
2.3 菌株发酵粗提液的体外抗菌活性
放线菌发酵粗提液的体外抗菌活性结果总结于表2,73株放线菌中有55株放线菌至少对一种检定菌表现出抗菌活性,阳性率为75.3%,其中抗金黄色葡萄球菌比例为57.5%,抗枯草芽孢杆菌比例为54.8%,抗白色念珠菌比例为37.0%,抗绿脓杆菌比例为17.8%,抗大肠杆菌比例为11.0%,菌株发酵产物的体外抗菌活性以抗革兰氏阳性细菌为主。
2.4 线虫体内抗菌活性
73株菌株发酵提取液的线虫体内活性结果(表3),显示8个(11%)发酵粗提液为阳性,其中3株菌株的代谢产物使线虫存活率达到80%-100%,2株菌株的代谢产物使线虫存活率达到65%-80%,3株菌株的代谢产物使线虫存活率达到50%-65%。结合体外抗菌活性结果,其中2个发酵粗提液同时具有体内和体外活性,6个发酵粗提液仅有体内活性而无体外活性。
菌株PF188(Nonomuraeasp.)的代谢产物对感染金黄色葡萄球菌和大肠杆菌的线虫均有100%的保护作用,其对感染金黄色葡萄球菌的线虫体内活性与50 μg/mL的万古霉素相当,但该代谢产物无体外抗大肠杆菌活性。菌株PF187(Microbispora tritici)的代谢产物具有良好的体外抗金黄色葡萄球菌活性,对感染金黄色葡萄球菌的线虫有90%保护作用,同时该代谢产物无体外抗大肠杆菌活性,但对感染大肠杆菌的线虫有60%保护作用。菌株PF33(Streptomyces sparsus)的代谢产物对金黄色葡萄球菌和大肠杆菌均无体外活性,但对感染金黄色葡萄球菌的线虫有80%保护作用,对感染大肠杆菌的线虫有50%保护作用(图4)。预示菌株PF188、PF187、PF33的代谢产物具有不同于传统抗生素的作用机制。
3 讨论
七个西藏湖泊沉积物中链霉菌属菌株为丰度最高的放线菌优势菌属,与罗红丽等[21]和夏占峰等[22]报道的研究结果基本一致。小单孢菌属是仅次于链霉菌属的优势菌属,在左用措、昆仲措、戈芒措、昂仁金措、吉力湖中均有分布,而在纳木措和芦布措中未分离得到。比较各湖泊生态环境参数,推测小单孢菌属的分离频度与湖泊的pH值呈正相关,与张学武[23]报道的小单孢菌属是分布于低温、碱性水体生境中的优势菌属的研究结果一致。近年来,国内外许多学者从湖泊生境分离获得大量放线菌。如夏占峰等[22]采用不同盐浓度的9种选择性培养基从艾丁湖沉积物中分离放线菌,获得8个属的55株放线菌;Zothanpuia等[24]从印度东北部淡水湖泊分离得到9个属的68株放线菌。而本研究分离菌株数相比其他类似研究偏少[25],其原因除了生态环境因素外,可能与预处理方法单一、分离培养基设计未考虑盐度因素等有关。深入挖掘西藏湖泊放线菌资源的多样性,需要结合湖泊水体的物理化学参数,从培养基设计、样品预处理方法等多个环节建立优化的分离体系。
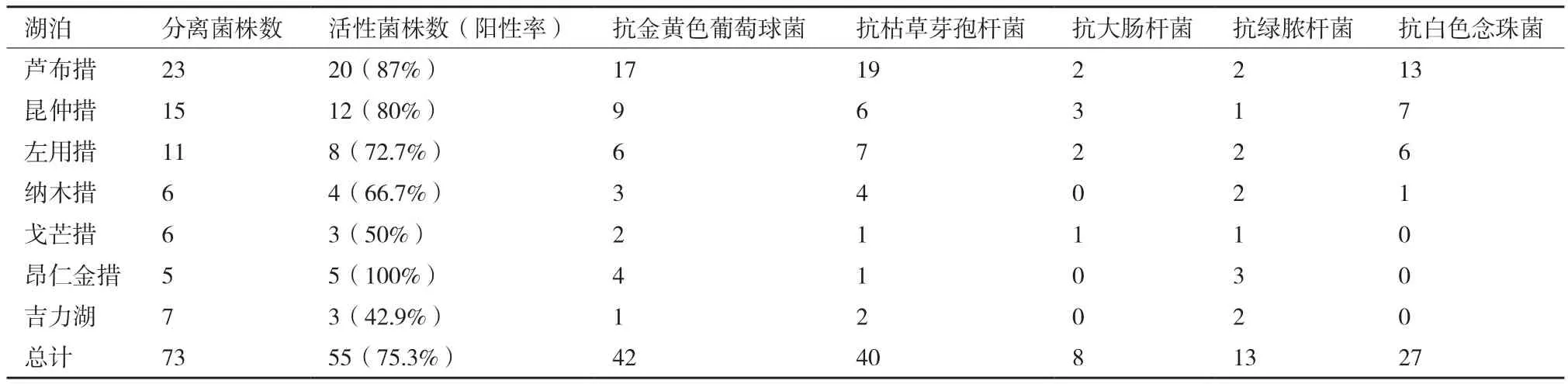
表2 西藏湖泊放线菌的体外抗菌活性统计结果
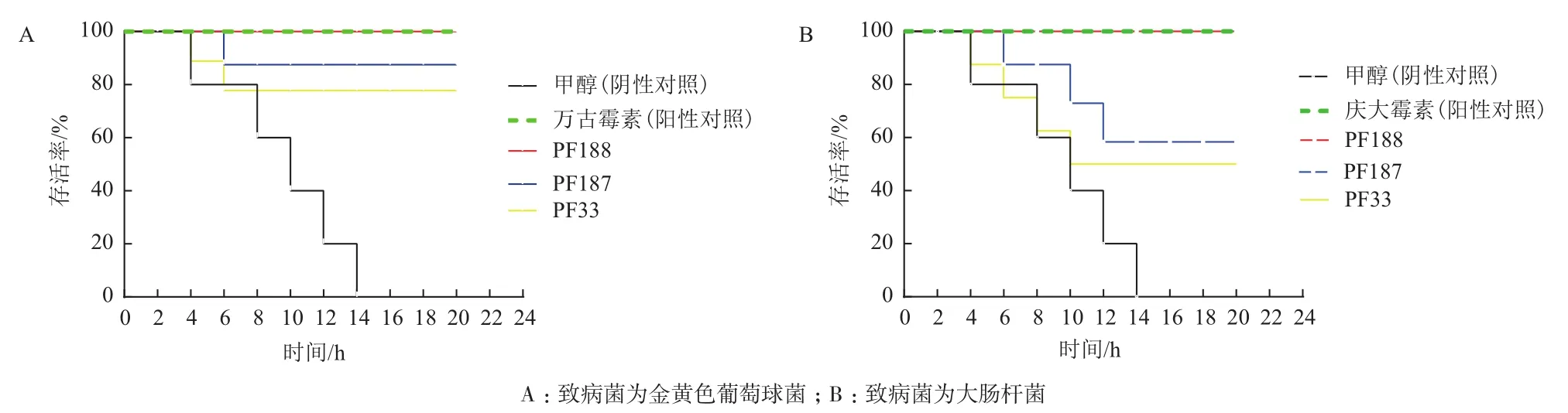
图4 三株菌株代谢产物对感染致病菌的线虫的保护作用
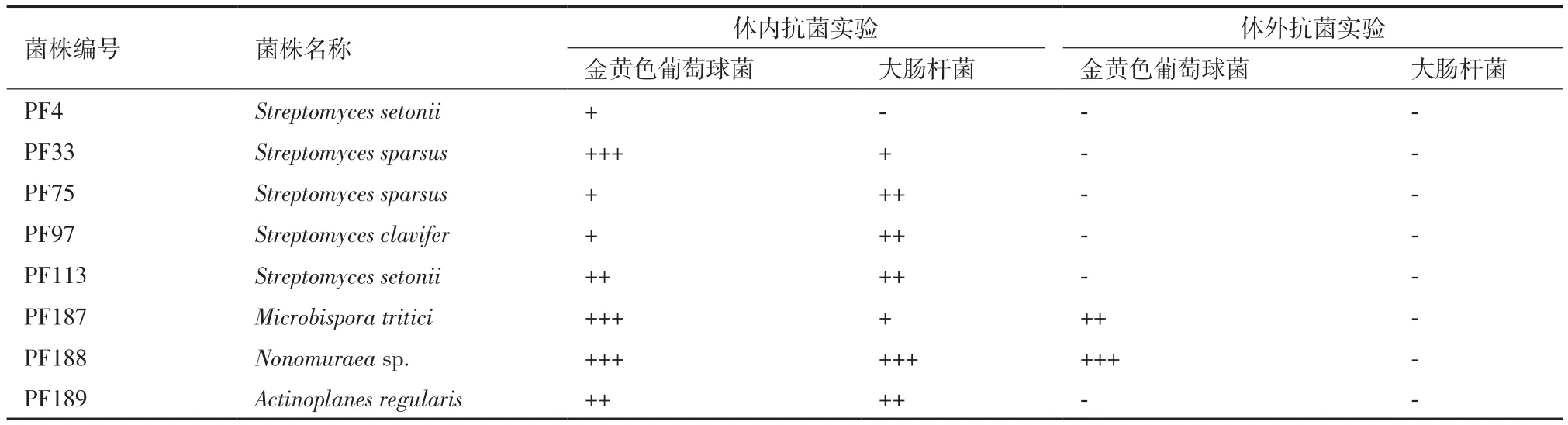
表3 西藏湖泊放线菌阳性菌株的体内外抗菌活性统计结果
秀丽隐杆线虫作为一种模式生物,因虫体较小、通体透明、生命周期短、易于实验室培养,近年来被广泛应用于药物高通量筛选中[26-27]。利用秀丽隐杆线虫感染模型,除了可以筛选得到体内外均有抗菌活性的化合物外,还可以获得仅有体内活性而无体外活性的化合物,包括作用于致病菌毒力因子或增强宿主免疫力的抗菌物质[28-29]。线虫感染方法分为固体平板法和液体法,大多数学者采用液体感染法的原因主要由于其操作简便、数据偏差较小。本研究采用秀丽隐杆线虫模型筛选获得的活性菌株次级代谢产物对金黄色葡萄球菌和大肠杆菌有良好的抑制作用,对这些次级代谢产物开展活性化合物分离纯化、结构解析和作用机制研究,有望获得具有新作用机制的抗菌先导化合物。
4 结论
(1)从西藏7个湖泊中分离得到73株放线菌,分属于4个目5个科8个属,其中含有潜在新种3株,链霉菌属和小单孢菌属为优势菌属。(2)放线菌发酵粗提液体外抗菌活性结果中,对至少一种检定菌具有抗菌活性的菌株数为55株。采用秀丽隐杆线虫体内抗菌活性筛选模型,筛选获得阳性菌株数8株,其中菌株PF33(Streptomyces sparsus)、PF187(Microbispora tritici)、PF188(Nonomuraeasp.)具有强大的线虫体内广谱抗菌活性,值得深入研究。
猜你喜欢
今日农业(2022年14期)2022-09-15
现代装饰(2020年10期)2020-10-28
天然产物研究与开发(2018年10期)2018-11-06
中成药(2018年2期)2018-05-09
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17
小学阅读指南·低年级版(2017年2期)2017-03-23
天然产物研究与开发(2016年6期)2016-06-05
天然产物研究与开发(2014年7期)2014-04-27
天然产物研究与开发(2014年6期)2014-04-27
中国医学科学院学报(2013年2期)2013-03-11
