
带状采伐对毛竹林林下植被物种多样性的影响
2020-07-31 08:21詹美春官凤英晏颖杰张美曼郑亚雄
生态学报 2020年12期
詹美春,官凤英,晏颖杰,张美曼,郑亚雄
国际竹藤中心,竹藤科学与技术重点实验室, 北京 100102
物种多样性是生物多样性在物种水平上的表现形式,它对生态系统功能的作用一直是生态学领域的重要研究问题之一,随着生态学研究在世界范围内的发展,物种多样性也愈发成为一个基础而重要的因素[1- 2]。α多样性指群落或生境内物种数及物种分布的均匀程度,其测度方法主要为物种丰富度、多样性指数、均匀度指数和等级-多度曲线四类,自生态学产生至今,生境内多样性研究都依以上四类方法演变和完善[3- 5]。干扰是影响自然生态系统演替的重要因素之一,许多植物群落物种组成与干扰具有密切关系,尤其在自然更新方面具有不可替代的作用,干扰对森林生态系统主要生态过程的影响以及森林生态系统对干扰的响应等问题成为森林生态研究领域的热点问题,干扰对物种多样性的影响取决于干扰的强度、频率等特征,如中度干扰假说、最大干扰水平等理论都描述对生物多样性有促进作用[6- 10]。植物群落的生物量是生态系统生物量的基础,植物生物量水平是森林生态系统功能的重要表现形式,研究物种多样性与生物量间的关系,对阐明植物物种多样性对森林生态系统功能的作用具重要意义[11- 13]。
毛竹(Phyllostachysedulis)用途广泛,在食品、医药、新材料、荒山造林及固碳等经济和生态方面展现出极大的价值[14- 18]。但由于毛竹林经营管理成本上升,竹材价格连年下滑,毛竹林经营管理日渐困难[19- 21]。基于竹子庞大、复杂的地下系统及其克隆生理整合的生态学特性,目前,已有部分学者通过改变竹林采伐方式来调整传统竹林经营模式,如带状采伐模式,此类研究将有可能实现竹林机械化经营,从而达到增产减投目的[22- 26]。大面积、高强度人为干扰对竹林生态系统产生极大扰动,特别是采伐形成的林窗空间对地面物种有着强烈影响,相关研究表明中度干扰对毛竹林物种多样性提升最具促进作用,但关于毛竹纯林不同面积林窗林下物种多样性的研究尚为空白[27- 31]。
本研究对毛竹林进行带状采伐,本质上是对毛竹林进行不同程度的干扰。禾本科植物生长特性致使毛竹林采伐后形成的大面积林窗能再短时间内完成闭合,因林窗面积大小、恢复时间不同,林下植被在此期间内呈现不同的生长、竞争规律。研究带状采伐毛竹林林下植被物种多样性,一方面,能为促进竹林生物多样性提供一定的参考;另一方面,可为竹林经营管理过程中协调生产与生态之间的关系提供一定的数据支撑。
1 研究区及研究方法
1.1 研究区概况
研究区位于江苏省宜兴国有林场内(31°15′1″—31°15′12″N,119°44′2″—119°44′8″E & 31°15′21″—31°15′40″N, 119°44′41″—119°43′52″E),在该地区毛竹林主要基于传统人工经营管理,并不对竹林进行施肥。主要的人为活动有竹林灌草清理、冬笋春笋采挖、竹材采伐和旅游观赏,试验区内的所有人类行为可控,在研究区域设置护栏,对试验样地进行保护。
本研究依靠江苏省宜兴竹林生态系统定位观测研究站开展,该站位于北方散生竹区和江南竹区交界,是毛竹分布区东北边缘地带。研究区地处典型的亚热带季风气候区,受太湖和海洋影响,年降水量和大雨的时空分布不均匀,夏季降水集中,年平均最高温度在20—24℃之间,年平均最低温度在12—14℃之间[32]。研究区独特的水热环境为当地中小径型毛竹提供了良好的生长环境。除洪涝灾害外,该地区受台风影响明显。
1.2 样地设置及取样方法
通过预调查选取林分条件一致的毛竹林,考虑毛竹地下系统对竹笋生长发育及竹林恢复影响的范围,设置5种不同宽度的毛竹林带状采伐样地(表1)。经过一年的自然恢复,分采伐带和保留带,对每个条带进行了物种多样性调查和生物量调查,试验地采伐及恢复过程如图1所示。
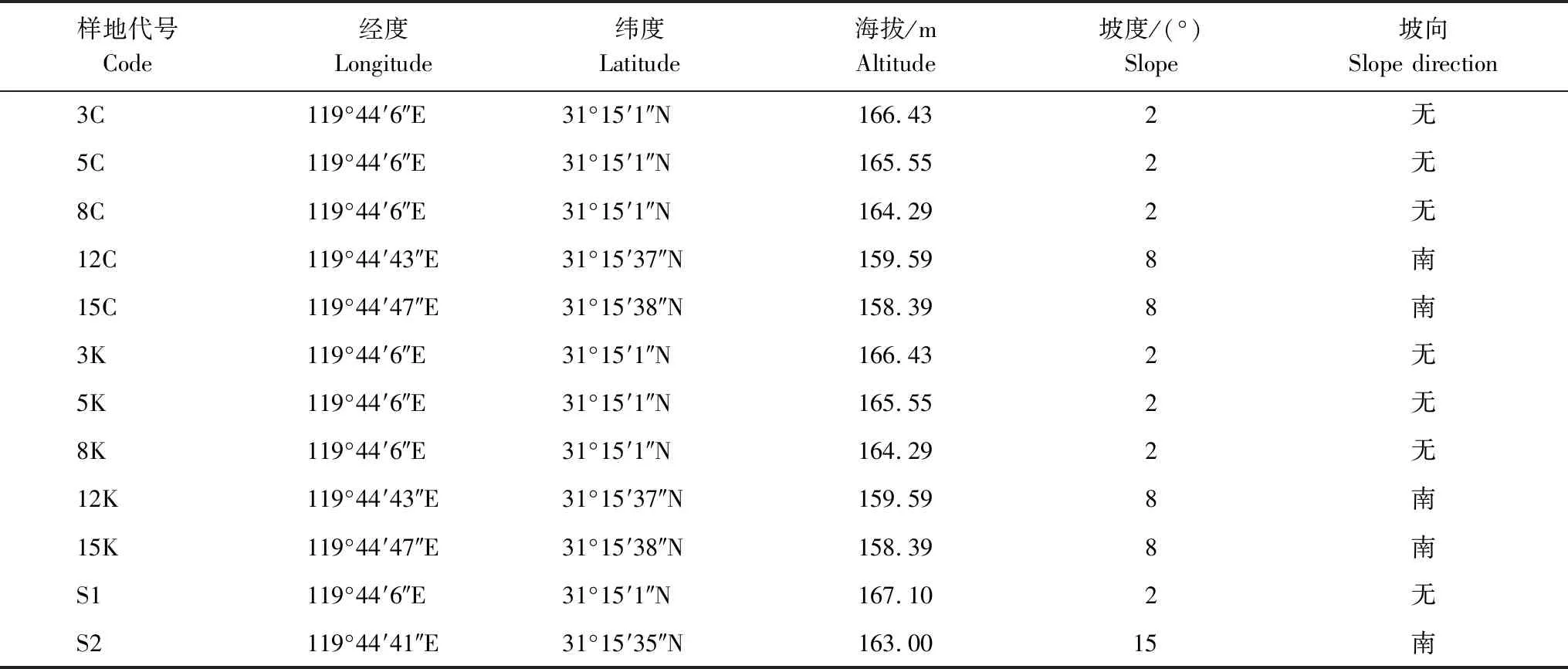
表1 样地地理信息表
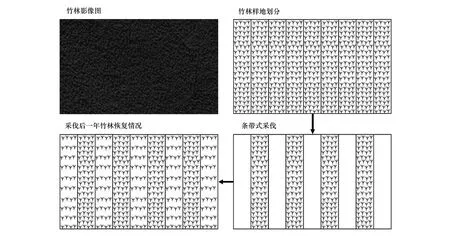
图1 带状采伐毛竹林恢复过程示意Fig.1 Recovery process of bamboo forest
样地采用GPS定位,测定相关地理位置。每个宽度的样地总面积控制在600 m2左右。共设置33块样地,其中采伐带和保留带样地计27块,对照样地计6块(20 m×20 m),采伐带带宽分别为3 m、5 m、8 m、12 m、15 m,长度皆为20 m。3 m采伐带11个重复,5 m采伐带6个重复,8 m采伐带4个重复,12 m采伐带3个重复,15 m采伐带3个重复;保留带的宽度与采伐带相同。样地设置详细信息如表2所示。
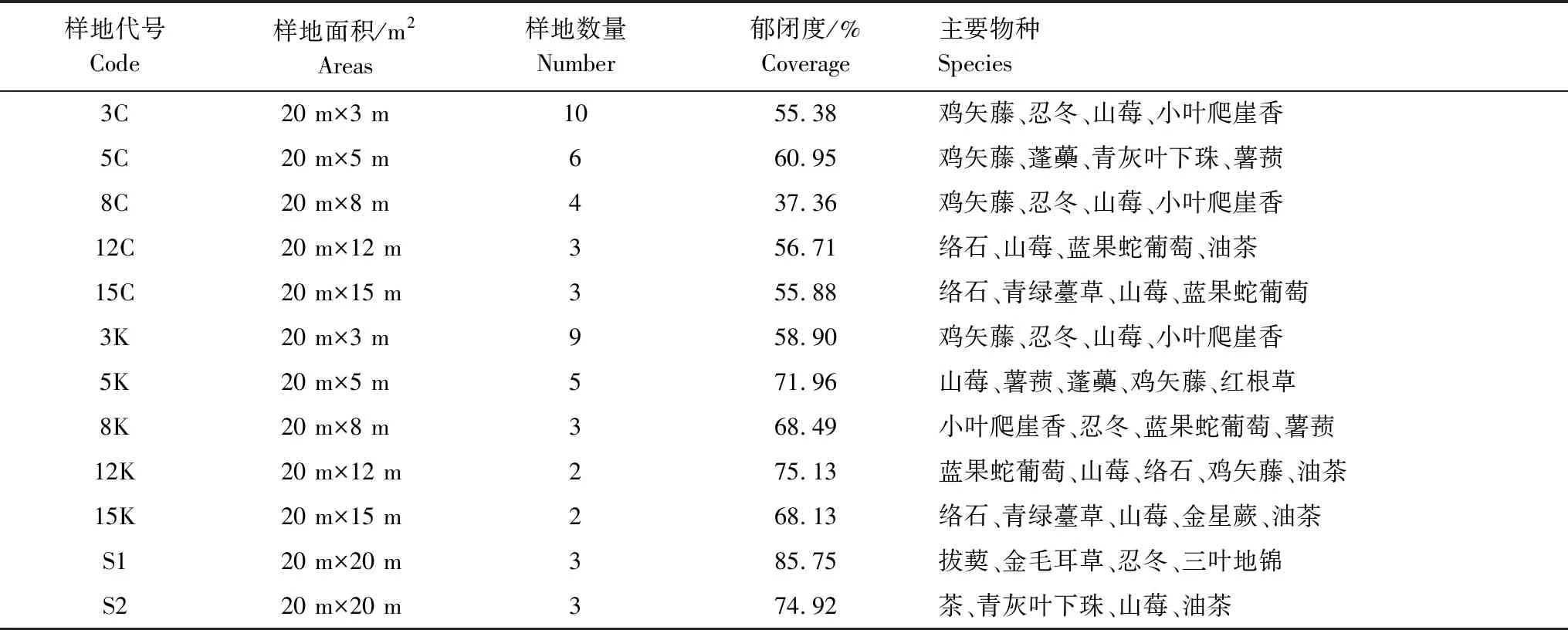
表2 样地布设及林下植被恢复情况表
样地代号同表1;主要物种根据重要值(>0.05)进行判断
1.3 统计分析方法
在2018年8月中旬至10月上旬,对林下植被进行全部调查,并参照5 m×5 m标准灌木样方、1 m×1 m的草本样方进行数据统计,具体调查内容包括种名、冠幅(盖度)、高度和数量,进行物种多样性研究。设置1 m×1 m样方进行生物量典型取样,在野外进行鲜重测定,在室内对其余指标进行测定。
(1)林下植被以匍匐藤本和禾本科矮小草本为主,因此未将植株高度纳入重要值内进行计算,重要值测定方法如下[12,33-34]:
灌木层重要值IV=(相对密度+相对盖度+相对频度)/3
草本层重要值IV=(相对密度+相对盖度+相对频度)/3
(2)毛竹群落物种多样性测定以重要值作为α多样性指数测度依据,选用Shannon-wiener指数、Simpson指数、Pielou均匀度指数和Gleason物种丰富度指数四种常用测度方法对物种多样性进行描述[33- 35]:
Gleason物种丰富度指数(Dg):
Dg=S/lnA
上述各式中,S为样地中物种数;Pi为种i的相对重要值;A为面积。
1.4 数据处理与图表制作
数据统计采用Excel 2016软件,采用SPSS 22.0软件进行相关性分析、T检验及方差分析,图表制作采用Graph Pad Prism 8软件。
2 结果与分析
2.1 林下植被物种丰富度
调查统计,记录到研究地共计有53科95属110种物种,其中,灌木计37科64属76种,草本计18科30属34种。如图2所示,林下植被以蔷薇科、菊科、大戟科、茜草科、百合科、唇形科、禾本科、马鞭草科、木兰科和安息香科等为主,物种丰富度较高,各科物种构成相对均匀。
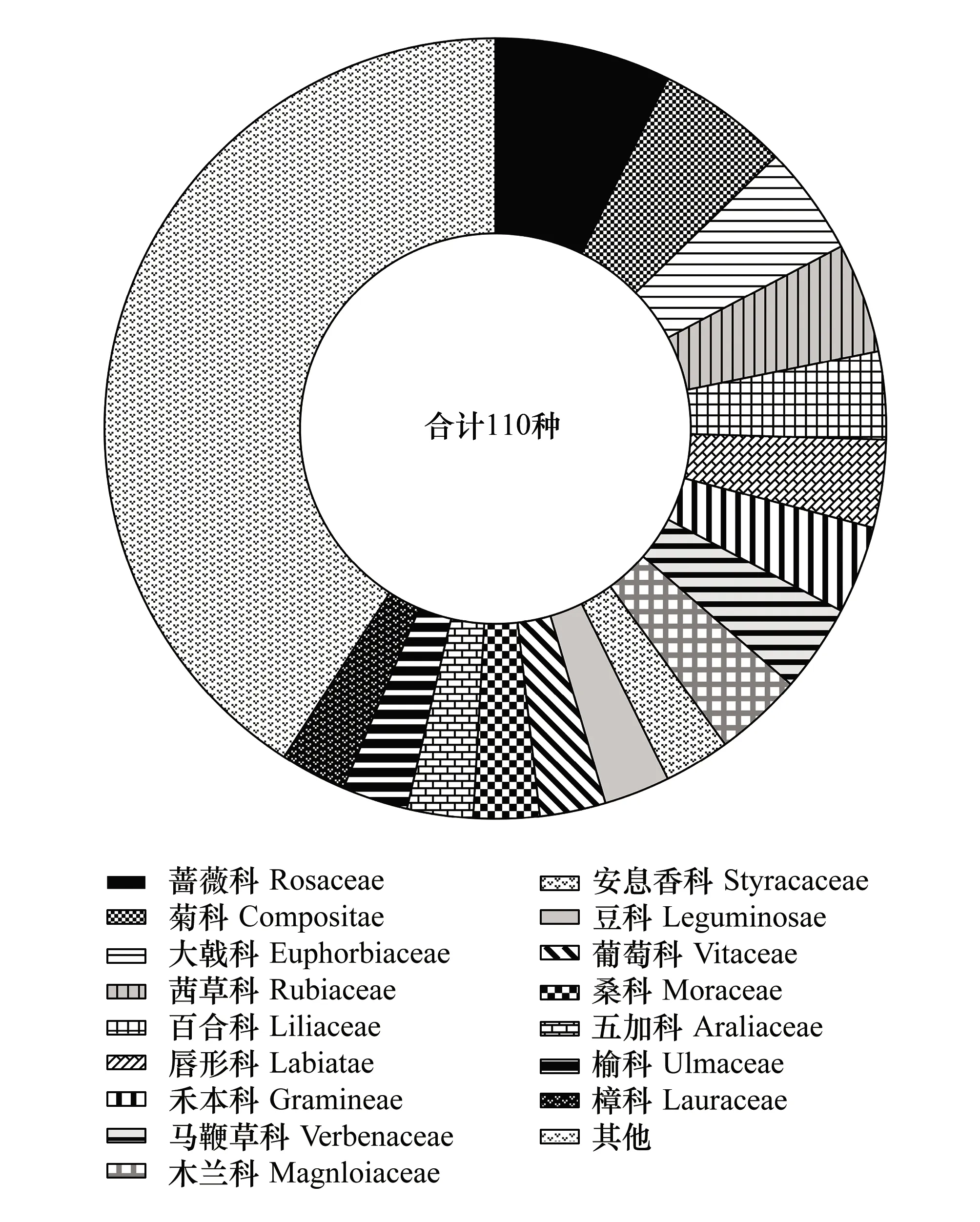
图2 物种构成Fig.2 Species composition
由表3和表4可以看出,参考样地S1林下灌木和S2林下草本差异性大于S1林下草本和S2林下灌木;不同宽度带状采伐样地及相同宽度采伐样地条带间差异极显著,林下植被物种丰富度差异明显;3K、5K、8K保留样地及S1样地林下灌木条带间差异极显著,12K、15K保留样地及S2样地林下灌木差异显著;3K、5K、8K保留样地及S2样地林下草本条带间差异极显著,S1样地林下草本差异显著,12K与15K保留样地林下草本不具显著差异。
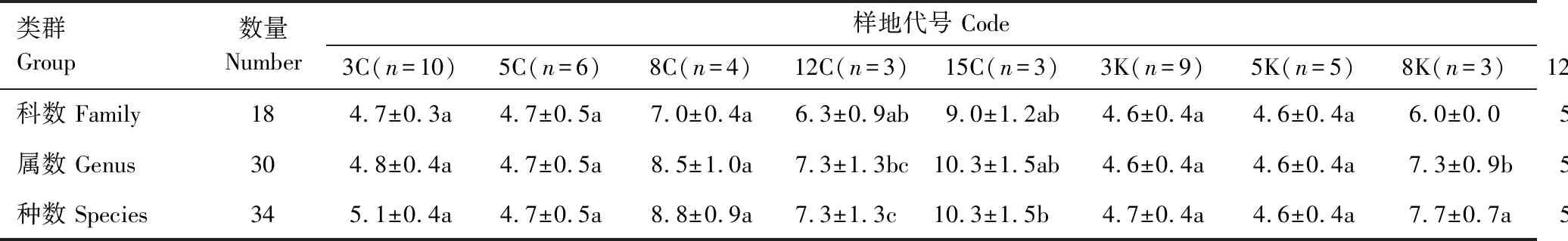
表4 草本物种数一览表
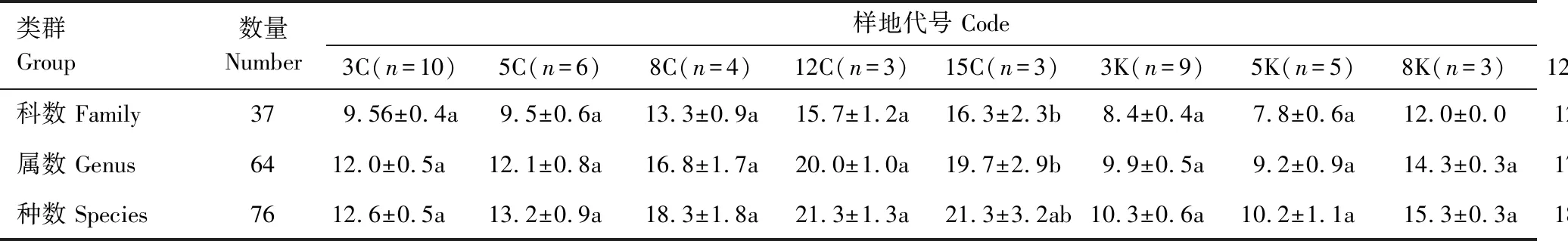
表3 灌木物种数一览表
由图3可以看出,采伐样地间,8C与15C样地林下植被物种丰富度较其余样地高,8C样地灌木丰富度高于15C样地,但草本丰富度低于15C样地,物种丰富度最低为S1样地;保留样地间,3K、15K与S2样地灌木丰富度高于其余样地,3K与8K样地草本丰富度高于其余样地,物种数量大小排序为3K>8K>15K>5K>S2>12K>S1。
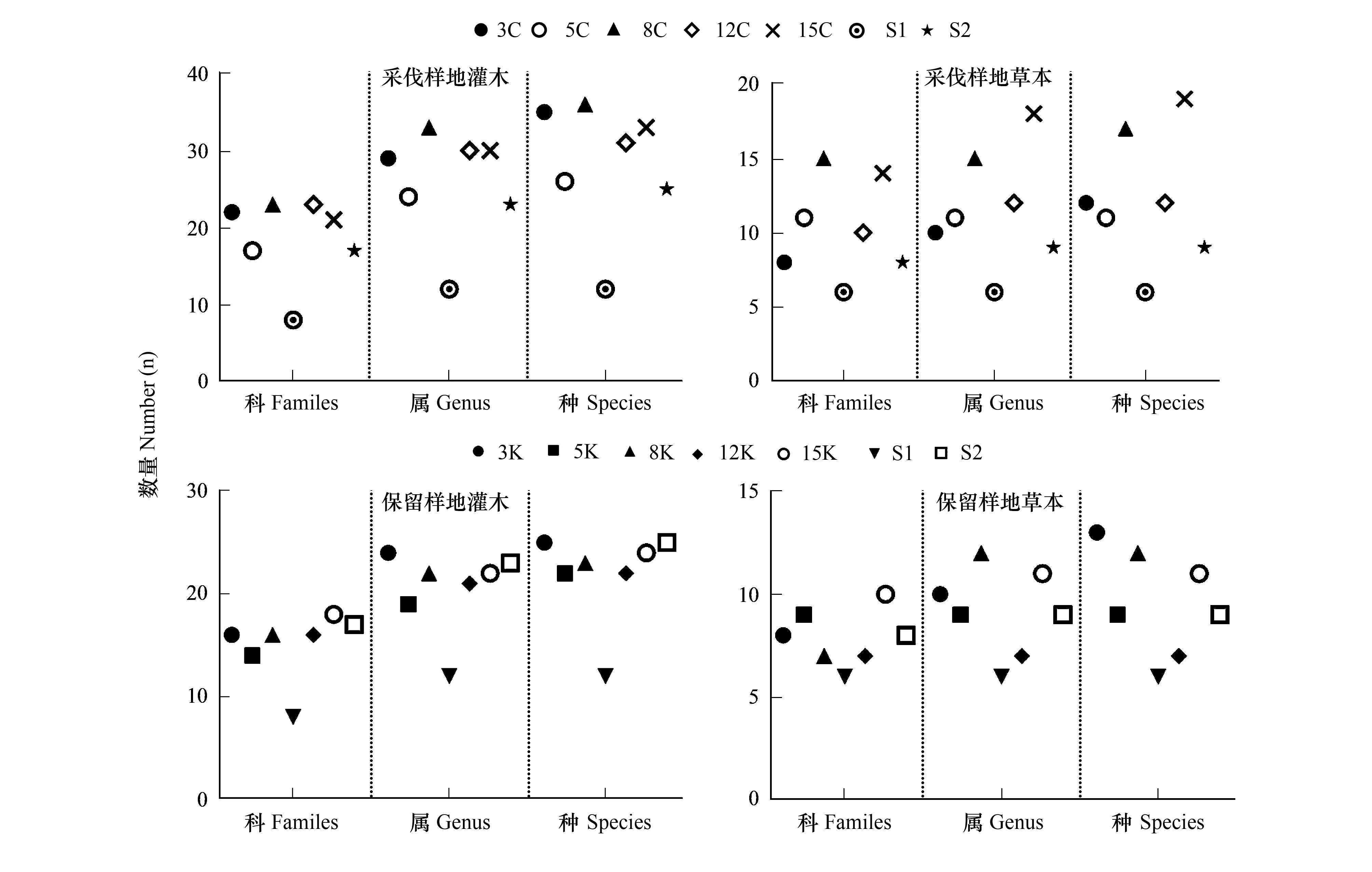
图3 类群数量Fig.3 Number of group样地代号中3、5、8、12、15表示采伐宽度,C:采伐区域(采伐带);K:保留区域(保留带);S1:传统经营1号样地,对应3 m、5 m、8 m宽度带状样地;S2:传统经营2号样地,对应12 m、15 m宽度带状样地
2.2 灌草物种多样性分析
从图4可以看出,林下植被在不同宽度采伐作业的影响下,呈现出巨大的差异。Shannon-wiener(H′)指数最高值出现在8 m样地、15 m样地及S2样地,其中,15 m采伐带(3.17)、8 m保留带(3.05)及S2样地(2.97)达到最高;最低的为5 m采伐带(2.51)和S1样地(2.49)。Simpson(D)指数在8 m保留带(0.96)、15 m采伐带(0.94)及S2样地(0.93)达到最高;最低的为5 m采伐带(2.51)和12 m采伐带(2.49)样地。林下植被Pielou(E)均匀度指数介于0.70—0.86之间,S1样地(0.86)及8 m保留带(0.86)物种分布相对最均匀,5 m采伐带(0.70)及3 m采伐带(0.71)最低,物种分布相对不均匀。 Gleason(Dg)物种丰富度指数在8 m采伐带(8.20)、15 m采伐带(7.50)和3 m采伐带(7.35)达到最高,S1样地(2.40)最低。对带状样地间物种多样性进行显著性分析得知(图5),采伐带H′、D、Dg值之间差异大,保留带H′、D、Dg差异较小,E值在采伐带的差异略大于保留带。
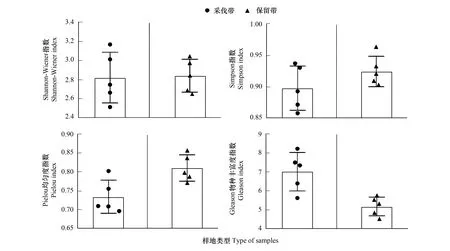
图5 带状样地物种多样性指数显著性(SD)Fig.5 Species diversity index significant on strip plots (standard deviation)
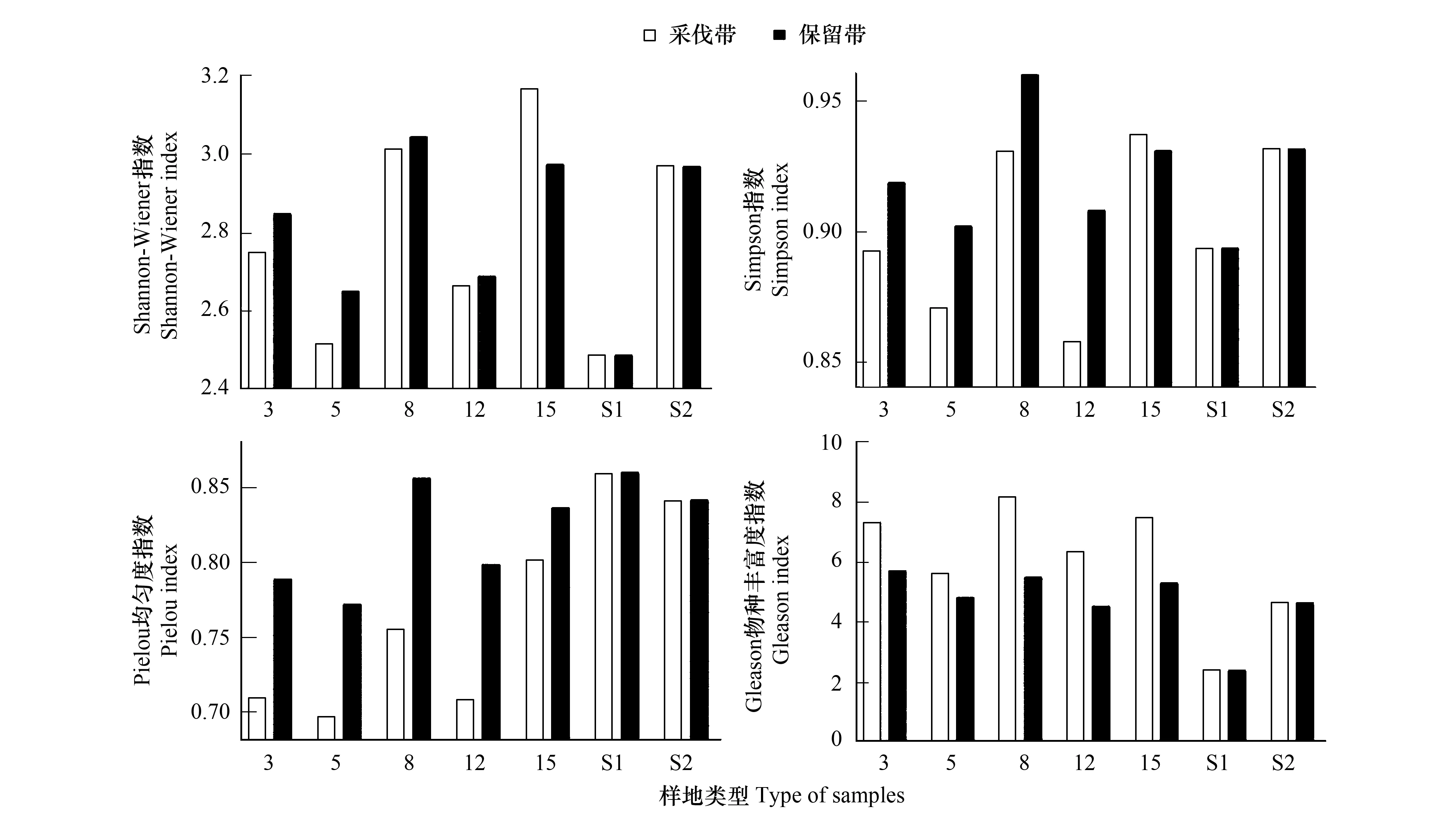
图4 物种多样性指数Fig.4 Species diversity index
由上可知,林窗(干扰)越大,物种多样性表现越优,α多样性各项指标走向一致,8 m及15 m带状样地指数最高。从物种多样性结果分析可知,速生快繁类、生态位广的木质藤本及禾本科草本植物在样地中占据重要位置,它们的生物特性和活动形成相应规律。3 m、5 m和8 m带状样地采伐带及保留带间差距明显,8 m带状样地物种多样性指数最优,人为采伐对8 m带状样地物种多样性的影响最大。
2.3 灌草生物量
受林地自身条件和毛竹林竞争的影响,江苏宜兴林下植被生物量偏小,如表5所示,不同类型样地生物量介于36.67—117.50 g/m2之间。总体而言,采伐样地林下生物量随着采伐宽度增加而增加,保留样地林下生物量随保留宽度变化不大,且生物量总量与参考样地相近;林下植被生物量以8 m保留带最低,15 m采伐带最高;由于林下植被种类不同,其生物量在根、茎、叶三部分也存在着差异,3 m、5 m、8 m样地生物量集中于茎部及叶部,S1、S2、12 m、15 m样地生物量集中于根部及叶部。
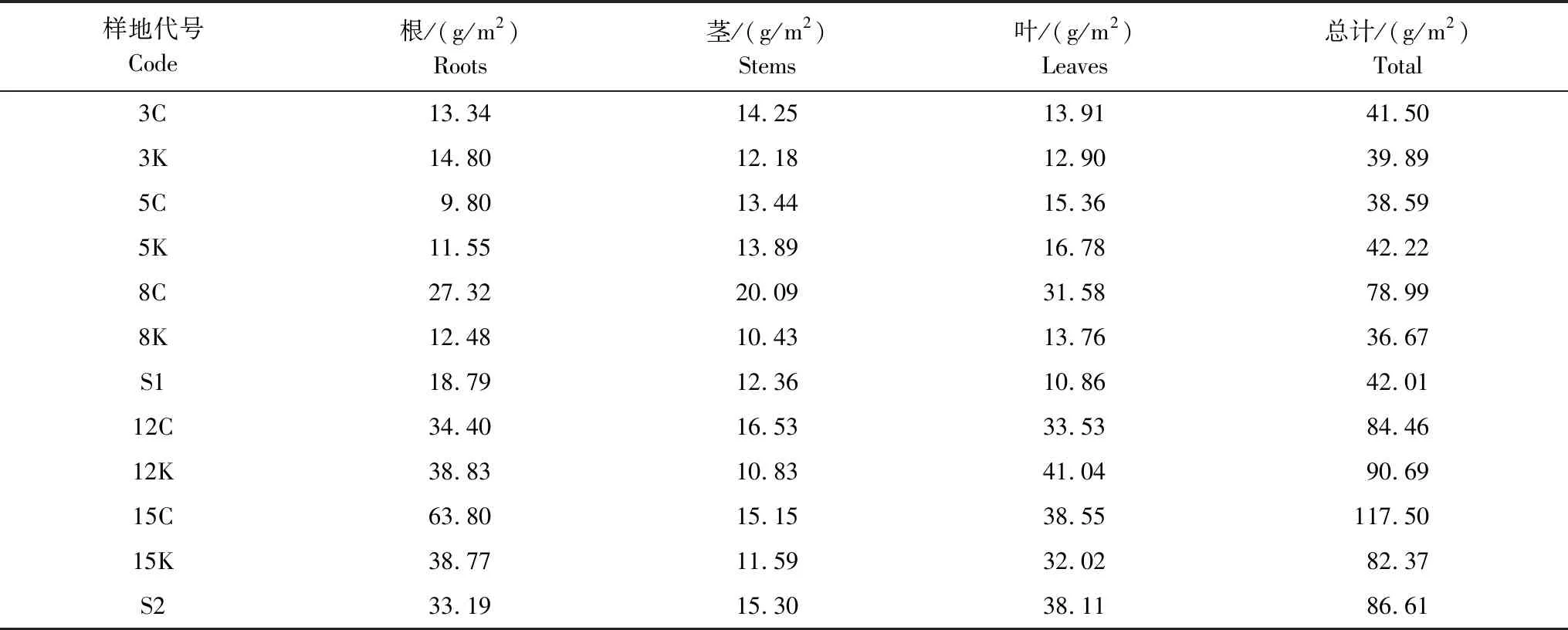
表5 各类型样地林下植被生物量
如图6所示,3 m、5 m、8 m样地茎部及叶部生物量较参考样地略多,3 m样地、5 m样地和8 m保留带根部生物量都比参考样地略少,12 m、15 m样地根部生物量较参考样地略多,但12 m采伐带叶部、15 m保留带茎部及叶部生物量都比参考样地略少; 15 m采伐带根部生物量较参考样地多30.60 g/m2,8 m采伐带根、茎、叶生物量分别多8.52、7.73、20.72 g/m2。
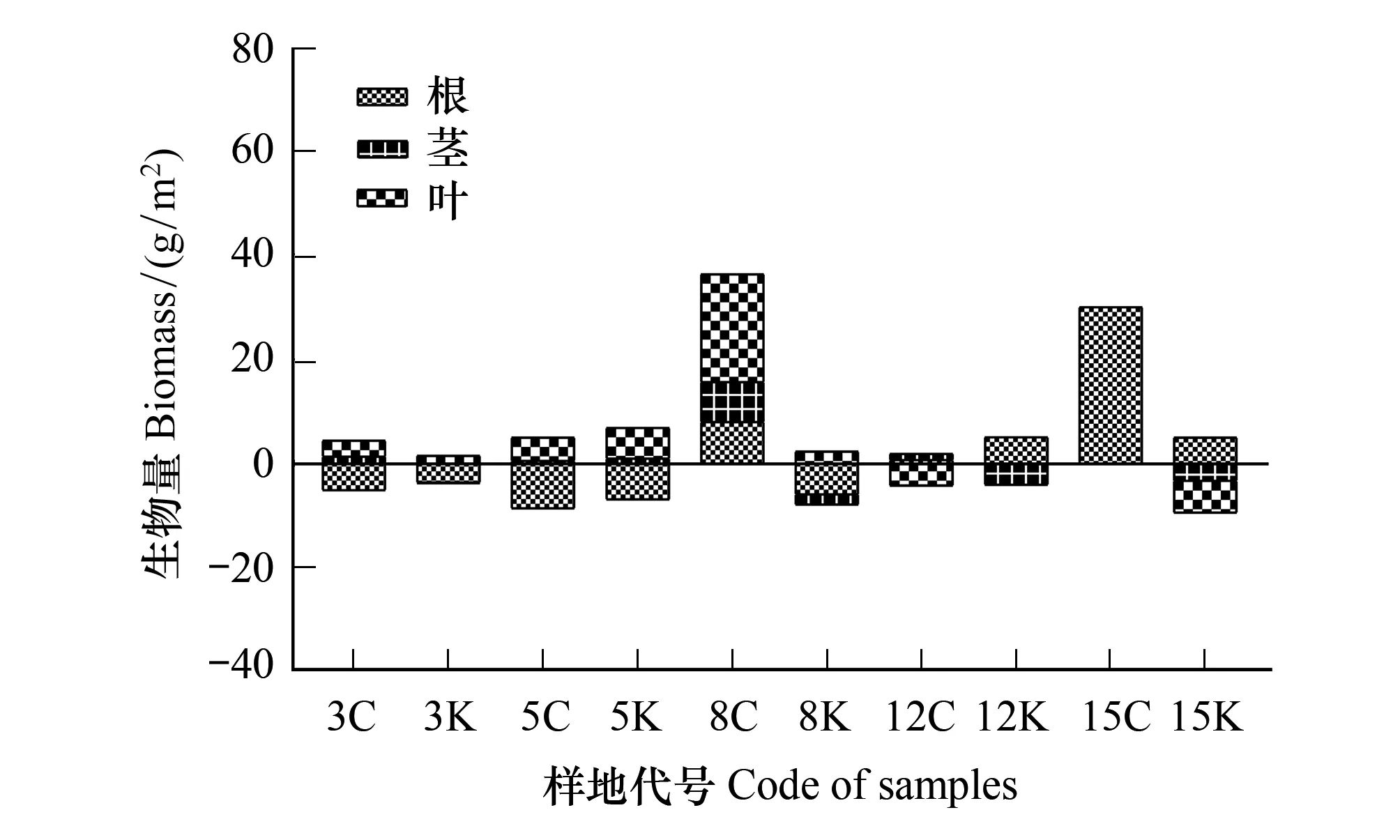
图6 带状样地与参考样地生物量比较Fig.6 Comparison value about strip plots and samples
如图7所示,3 m、5 m及12 m样地采伐带和保留带间生物量差异小,3 m样地采伐带生物量比保留带略多,5 m样地采伐带生物量比保留带略少,12 m样地采伐带茎部生物量比保留带多,但根部和叶部生物量比之要少;8 m样地采伐带生物量比保留带多42.31 g/m2,以根部及叶部为主;15 m样地采伐带生物量比保留带多35.13 g/m2,以根部为主。
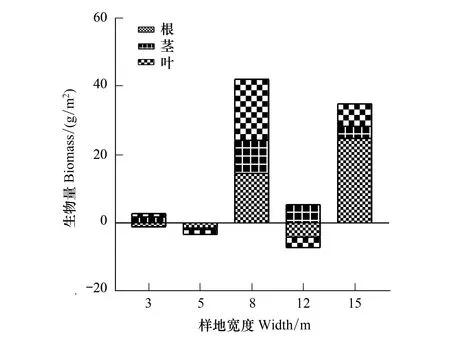
图7 采伐样地与保留样地生物量比较Fig.7 Comparison value about cutting plots and keeping plots
3 讨论
3.1 带状采伐模式与人为干扰
对毛竹林进行带状采伐,本质上是从经营层面对毛竹林进行不同程度的人为干扰处理,及在培育层面上对毛竹林采取不同模式的方法探究。在控制毛竹林样地总面积前提下,采伐样地越窄,采伐样地数量越多,换而言之,采伐宽度为3 m的样地干扰频率高、单个带状样地干扰强度小,采伐宽度为15 m的样地干扰频率低、单个带状样地干扰强度大。因此,无论是经营层面还是培育层面,最适宜毛竹林的带状采伐模式(人为干扰)要从生态价值(生物多样性、固碳、保水等)、经济价值(新生竹数量、新生竹质量、生物量、竹林机构、后续繁育等)方面进行综合判断,才能得出毛竹林带状采伐的最佳模式,即符合中度干扰理论。
3.2 物种多样性与带状采伐关系
判断森林功能,乔木层作为森林主体,起最主要作用,但林下植被层在维持生物多样性方面却是有着决定性作用的,特别是乔木层结构单一的情况下。以毛竹为主体的森林乔木层结构单一,偶有单一阔叶树、针叶树出现,且因毛竹林郁闭度大,致使林下植被种类、数量、盖度、高度等形态特征偏劣。带状采伐产生的大面积林窗改善了林下植被生长环境,林窗尚未闭合期间林下植被繁育的自然条件十分优渥,从而使得毛竹林生物多样性急剧上涨,但目前我国缺乏相关研究,因而探究带状采伐模式对毛竹林林下植被的影响具必要性。
4 结论
4.1 林下植被与毛竹郁闭度相关性讨论
表6表明,林下植被数量、物种丰富度和盖度均与竹林郁闭度呈显著负相关,以植被数量受郁闭度负面影响最大。由主成分分析可知,林下植被数量也是相应其余因子的决定因子,决定系数为0.391,因此,林下植被数量是带状样地内与竹林郁闭度响应最突出的因素。
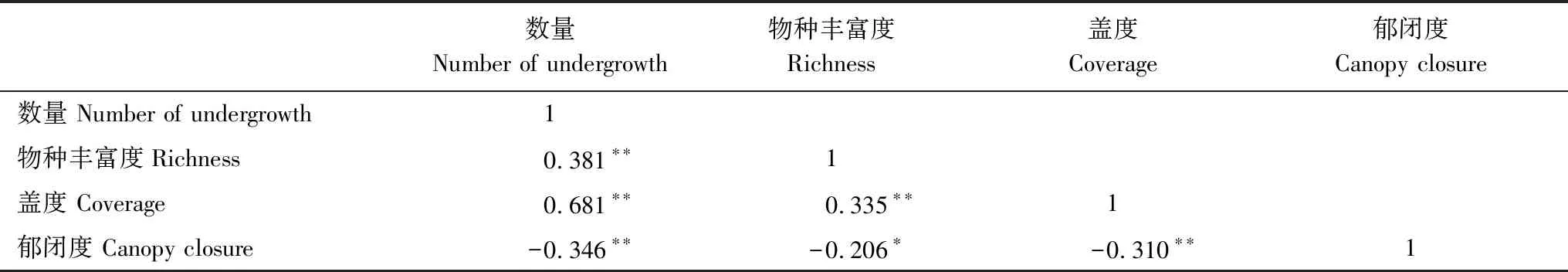
表6 林下植被与竹林郁闭度相关性及因子分析
4.2 采伐对物种多样性的影响
由表7可知,带状样地采伐宽度与E呈显著正相关,采伐频次与E呈显著负相关;样地数量与Dg呈典型正相关;H′与D呈显著正相关。因此,在采伐作业的影响下,带状样地宽度和样地数量是物种分布均匀程度重要因素,样地越宽,物种均匀度越高,样地数量越多,物种均匀度越低,所以参考样地物种均匀度最高。此外,从Dg与采伐频次的相关性可以看出,人为采伐对物种丰富度有促进作用,人为干扰促进了物种丰富度提高。但经过分析得知郁闭度与物种多样性不具典型相关性。
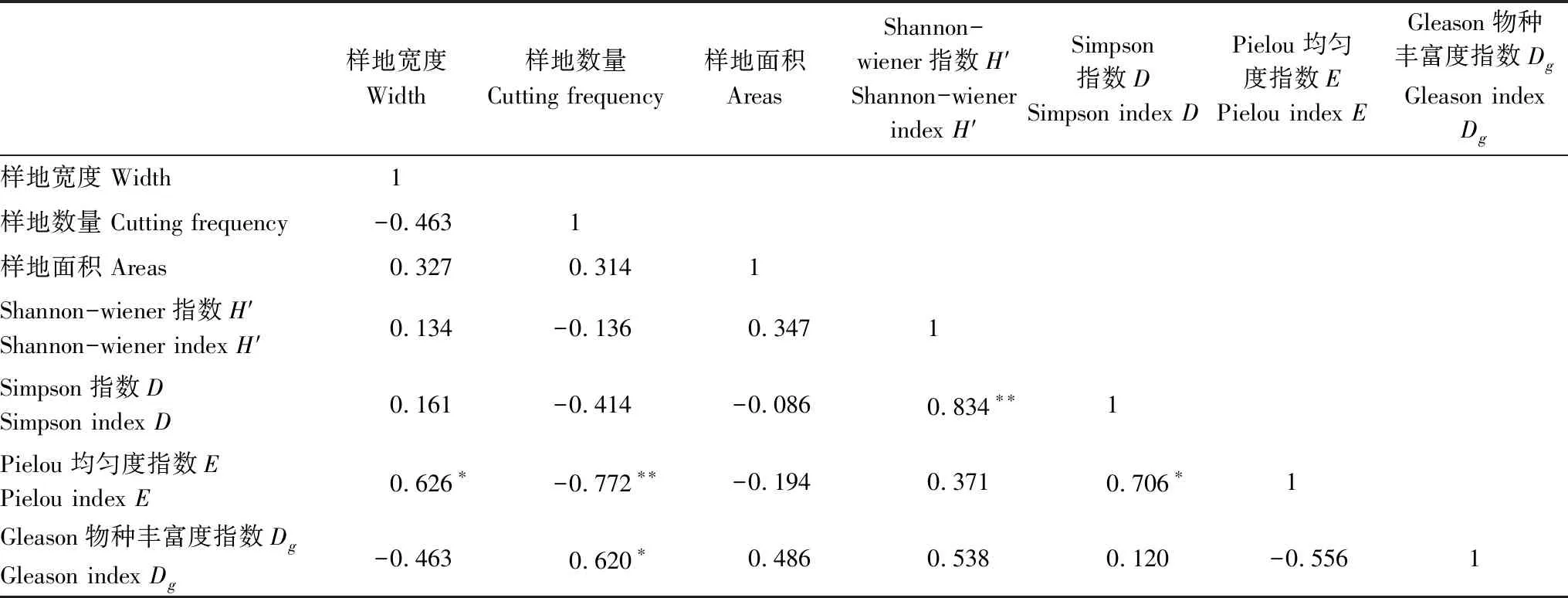
表7 采伐作业与物种多样性相关性分析
4.3 生物量与物种多样性及采伐作业相关性讨论
经过分析,江苏宜兴毛竹林群落林下植被生物量与物种多样性不呈显著相关关系,这可能是由林下植被生物量偏小且主要植被生物量增生类型所导致的。表8表明,生物量与样地带宽呈显著正相关,采伐频次及样地面积对生物量影响不显著。
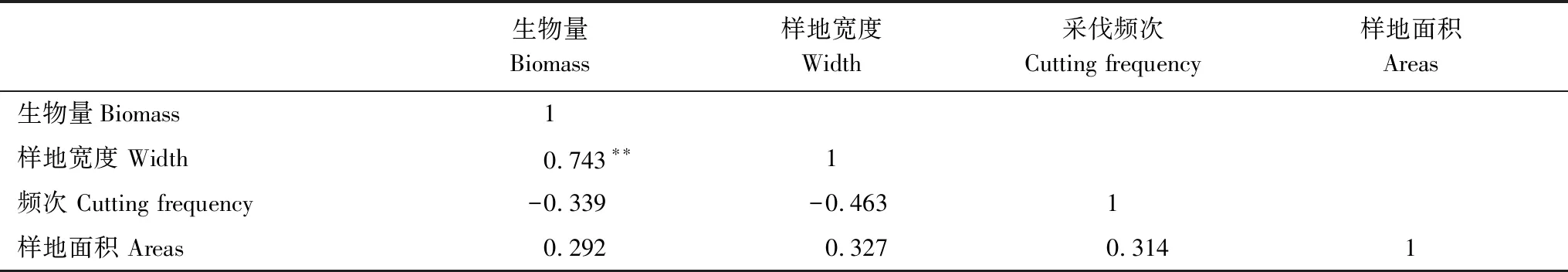
表8 生物量与采伐作业相关性分析
综合分析江苏宜兴毛竹林林下植被物种多样性在带状采伐作业下的结果,表现出以下特征:(1)研究区毛竹林林下植被物种丰富度较高;林下植被受郁闭度影响大,在数量、物种丰富度和盖度指标上均与郁闭度呈显著负相关;林下植被以匍匐藤本(数量和丰富度)和禾本科(数量)为主,如鸡矢藤(Paederiascandens)、忍冬(Lonicerajaponica)、山莓(Rubuscorchorifolius)、小叶爬崖香(Piperarboricola)、蓬蘽(Rubushirsutus)、青灰叶下珠(Phyllanthusglaucus)、薯蓣(Dioscoreaopposita)、络石(Trachelospermumjasminoides)、蓝果蛇葡萄(Ampelopsisbodinieri)、油茶(Camelliaoleifera)、茶(Camelliasinensis)、红根草(Lysimachiafortunei)、菝葜(Smilaxchina)、青绿薹草(Carexbreviculmis)等。(2)林下植被物种多样性本底数据小,但人为干扰在短时间内促进了其物种多样性发展;样地宽度与物种均匀度带呈显著正相关,采伐频次与物种均匀度呈显著负相关;采伐频次与物种丰富度呈典型正相关;Shannon-wiener指数与Simpson指数呈显著正相关。因此,采伐作业对物种分布均匀程度有着极大的影响,林窗环境改变了物种对环境的响应,种间竞争致使物种分布范围变广,在自然条件好的林窗下聚集式分布,物种数量剧增;但参考样地由于受人为干扰相对较小,在人为调控下,其物种均匀度反而高。(3)物种多样性与生物量不具典型相关性,可能为生物量偏小且主要植被生物量增生类型不同所导致。(4)经过研究,8 m和15 m带状样地对采伐作业的响应最佳,物种多样性各方面表现最优,生物量最大,表示其林下植被对林窗环境的利用最为全面,但具体原因还待进一步研究,建议从土壤肥力及太阳辐射方面进行补充。
猜你喜欢
林业资源管理(2022年6期)2023-01-18
天津农林科技(2022年2期)2022-04-19
现代园艺(2021年23期)2021-12-01
新农业(2020年18期)2021-01-07
花卉(2020年24期)2021-01-04
浙江农林大学学报(2020年5期)2020-11-05
河南畜牧兽医(2020年21期)2020-01-10
乡村科技(2018年8期)2018-06-27
北京航空航天大学学报(2017年3期)2017-11-23
中国药业(2014年17期)2014-05-26
