
BiIirubin and inflammation in neurodegenerative and other neuroIogicaI diseases
2020-07-28 09:01SriJayantiRitaMorettiClaudioTiribelliSilviaGazzin
Sri Jayanti, Rita Moretti, Claudio Tiribelli, Silvia Gazzin
1Italian Liver Foundation, Centro Studi Fegato, Trieste 34149, Italy.
2Neurology Clinic, Department of Medical, Surgical, and Health Sciences, University of Trieste, Trieste 34139, Italy.
Abstract Inflammation links neurodegenerative, neuropsychiatric and other neurological diseases (NDs) with acute brain events.It is responsible for the alteration of neurotransmission and circuity, brain architecture, and cell fate,affecting mood and personality (anxiety, depression and schizophrenia) and behavior (decline in cognitive, motor and speech abilities, altered sleep, fatigue, pain sensitivity and dementia).Inflammation is also a key component in systemic chronic diseases (cardiovascular disease, cancer, diabetes, and metabolic syndrome), in which bilirubin has been demonstrated to improve the diseases by acting as a multi-target antiinflammatory molecule, and where the evaluation of pharmacological modulation of the pigment level as a therapeutic approach has already started.While altered serum bilirubin levels have been reported in ND patients, the potential activity of bilirubin in the brain is vague.This review summarizes the available fragmentary information on the interplay of bilirubin with neuroinflammation, aiming to elucidate the pigment's role in the central nervous system environment.
Keywords: Neuroinflammatory diseases, bilirubin, brain, heme oxygenase, biliverdin reductase, inflammation,homeostasis
INTRODUCTION
Bilirubin (unconjugated - UCB) is the final metabolite of hemoglobin, which is processed in the liver (by conjugation to 1 or 2 molecules of glucuronic acid - CB: conjugated bilirubin) before elimination through urine and feces.Increased level of bilirubin (UCB and CB) in the blood is a well-recognized marker of hepatic damage.Recently, slightly elevated serum bilirubin concentration emerged as a biomarker of resistance versus chronic diseases[1].Epidemiological data have revealed a reduced prevalence of type 2 diabetes, obesity and metabolic syndrome, certain cancers, and especially cardiovascular diseases and related causes of mortality in Gilbert's syndrome patients,showing mild hyperbilirubinemia[2,3][Figure 1].The effect of the pigment has been demonstrated to be due mainly to the unconjugated (or indirect) moiety of bilirubin (UCB).UCB, especially in its free form [free bilirubin (Bf), the portion of UCB exceeding the serum albumin binding capacity], enters tissues from blood, acting as a powerful antioxidant molecule at nanomolar concentrations, where it is able to counteract 10,000 times higher levels of hydrogen peroxide (H2O2)[4].This capability has been related to the UCB-biliverdin cycle [Figure 2], which is able to regenerate the pigment consumed by oxidants and acts complementary to cellular glutathione (GSH)[4-6].In the last years,a more complex and fascinating scenario has emerged.First, various cells have been demonstrated to possess the complete enzymatic apparatus necessary for producing UCB themselves [Figure 2][7,8].A more expanded and intricate view has emerged from the discovery of the interplay of UCB (and the enzymes involved in its production and recycling, altogether called the “yellow players”) with cellular functions,signaling pathways, and defense/adaptation mechanisms (not restricted to redox state).Collectively, these findings suggest a greater role for the yellow players in cellular homeostasis and defense against diseases[2,3].The interplay of the yellow players with neurological and neurodegenerative diseases is still much less explored.In this article, we review the currently available evidence on the potential roles of bilirubin and other yellow players in neurological disease, with special emphasis on inflammation.We will also address the role of enzyme modulation in bilirubin metabolism, with the goal of increasing the systemic level of bilirubin and the protection it confers.
INFLAMMATION AND NEUROLOGICAL DISEASES: AN OVERVIEW
Chronic neurodegenerative pathologies are currently the most dominant clinical conditions.They comprise multiple sclerosis (MS), amyotrophic lateral sclerosis (ALS), Parkinson's disease (PD), Alzheimer's disease(AD) and vascular dementia.Consistent data show that the inflammatory process can be triggered by protein misfolding or protein accumulation, which are the initial events of a given pathology (i.e., amyloid for AD, tau protein for frontal dementia, alpha synuclein for PD,etc.)[9].
Aging has been associated with a low-grade sterile inflammatory status of the immune system, in which interleukin-6 (IL6), IL1β and tumor necrosis factor (TNF) are key players, more evident in an unhealthy state.Neuroinflammation can be considered one of the most important etiological factors in age-related neurodegeneration[10], associated with a reduction in neuron number, a decrease in neuronal arborization,and loss of spines[11].What fits particularly in this theory, is the strong evidence that in the aging brain,both macrophages and microglia react with a prolonged and overactive response to stimuli[12].This overactivation induces the production of reactive oxygen species and attracts peripheral leukocytes, and both these conditions can activate glial cells[13].The activation of glial cells promotes telomere shortening,which can be a contributor in different neurological conditions, such as AD[14].Resulting impaired phagocytosis alters the removal of toxic compounds, such as amyloid-beta (Aβ) and alpha-synuclein(aSyn)[15].Microglial activation is believed to be involved in the occurrence of deterioration in various neurodegenerative diseases, such as AD, PD, ALS, and MS[15].Microglia are the main immune cells of the central nervous system (CNS), and as the first line of defense, microglia play an important role in the inflammatory reaction[16].In AD, microglia are known for their role in cleaning up Aβ.Meanwhile, in PD,a pathological aSyn aggregation can induce microglial activation and dysfunction[17].Microglia are also directly involved in MS by producing cytokines such as tumor necrosis factor-a (TNF-a) and interferon-γ(IFN-γ), superoxide (O2-) or nitric oxide (NO), and release of proteolytic and lipolytic enzymes, which can destroy the myelin sheath[15,18].
Neuroinflammation and mitochondrial dysfunction are common features of chronic neurodegenerative diseases.Both conditions can lead to increased oxidative stress, which leads to excess reactive oxygen species and reactive nitrogen species, resulting in a cascade of events, with injury to polarized bilayers, lipid peroxidation, lysosomal intrinsic activity and autophagy; all these events together, permit self-potentiation of an inflammatory cascade[19].
Inflammation also plays a role in cerebral small-vessel disease (cSVD), which usually manifests as stroke,cognitive impairment, dementia, physical disability and depression[20].Inflammation in cSVD might be explained by the role of endothelial cells in the blood-brain barrier.Endothelial cells communicate with pericytes, astrocytes, microglia, and neural stem cells in the neurovascular unit.Damage to CNS tissues leads to the disruption of tight junctions that link endothelial cells[21].It is followed by the infiltration of neutrophils and monocytes, the activation of microglia and astrocytes and the invasion of T and B cells.This inflammatory response is mediated by the nuclear factor κ-light-chain-enhancer of activated B cells(NFκB) pathway and strongly associated with neuroinflammation in the acute phase of various vascular injuries such as stroke, diabetic retinopathy, and AD[22,23].
Bilirubin has been reported to act on all the above-mentioned molecular mechanisms.The following paragraphs review the state-of-the-art of what it is known about bilirubin and the brain.
BILIRUBIN: A MARKER FOR NEUROLOGICAL DISEASES
Cumulative clinical evidence demonstrates the alteration of total serum bilirubin (TSB) level in neuroinflammatory diseases, including schizophrenia, MS, PD, AD, ALS, stroke, diabetic retinopathy,etc.(for details, Table 1).For obvious reasons (limited CNS sample availability, especially in the early stages of diseases), few data on the potential molecular role of the pigment in these pathologies are available, and cause-effect studies are possible only by using experimental models.
Schizoprenia
Multiple contrasting studies exist on the potential correlation between the serum level of UCB and schizophrenia.Studies have reported an increase in UCB level, as in Gilbert's syndrome in schizophrenia compared to other psychiatric diseases (affective disorder and neuropsychosis) and healthy controls[24].
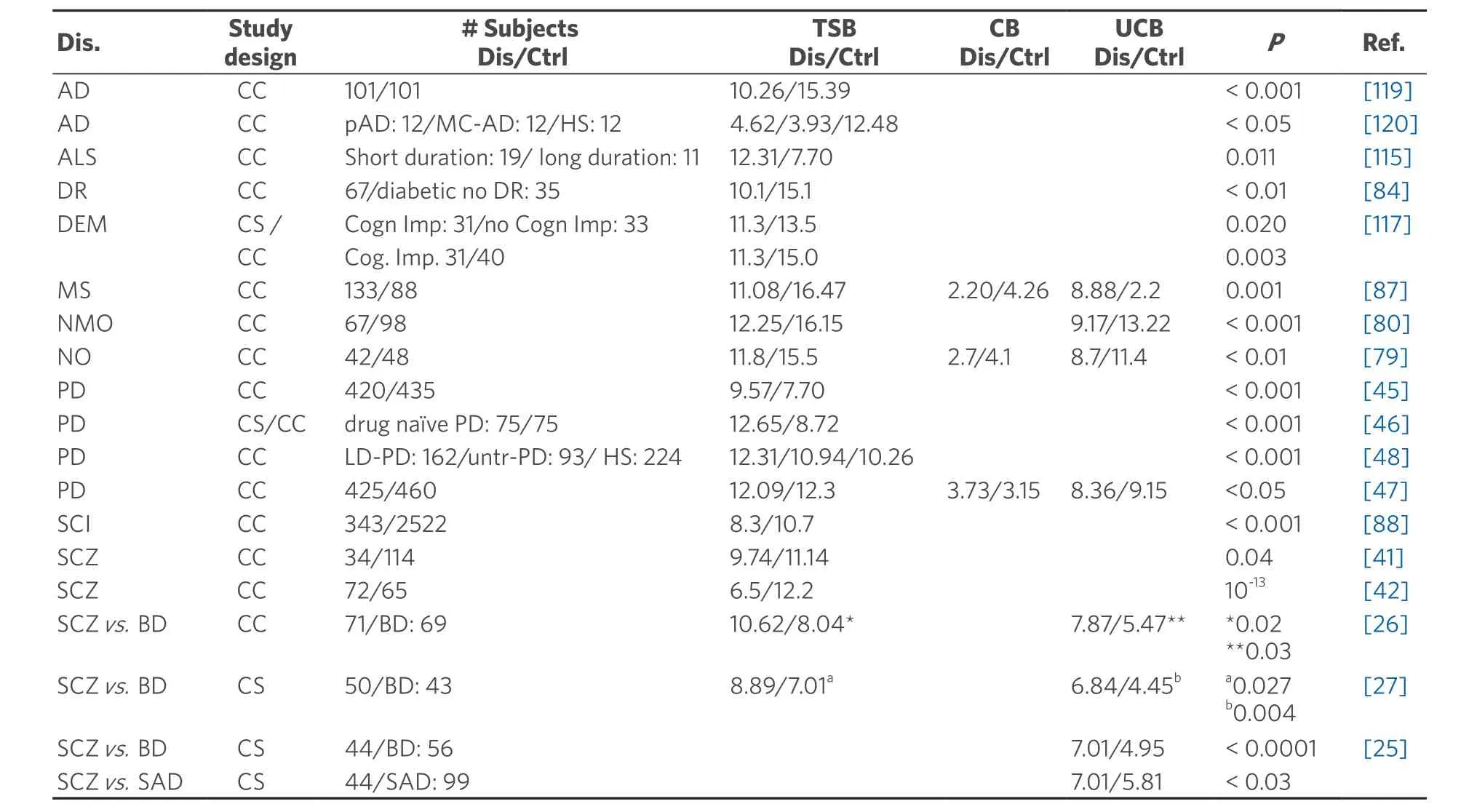
Table 1.TSB level in neurological diseases
More interestingly, after antipsychotic treatment, UCB decreases in 80% of subjects.Gama Marqueset al.[25]also reported that mean UCB levels are clearly higher in patients with schizophrenia than patients with schizoaffective disorder and bipolar disorder.Additionally, retrospective and recent prospective studies indicate that serum UCB level in patients with schizophrenia is higher than those with bipolar affective disorder (1.40- and 1.53-fold, respectively)[26,27].Notably, all these studies included only subjects with normal liver enzymes and no pre-existing liver disease or other confounding factors, further emphasizing the pivotal role of UCB increase.UCB can be viewed as a potential biomarker to distinguish schizophrenia from other psychiatric disorders[28].
UCB level also shows a correlation with schizophrenia symptoms.Patients with hyperbilirubinemia have shown significantly higher scores on the positive and general psychiatric subscales of the PANSS (Positive and Negative Syndrome Scale)[24].In concordance with the previous finding, UCB elevation has been found frequently in psychotic episodes of schizophrenia[25,29].Radhakrishnanet al.[26]reported that UCB levels are higher in paranoid schizophrenia than non-paranoid schizophrenia.Meanwhile, a recent study by Pradeepet al.[27]found no association between serum UCB levels with the severity of psychopathology in schizophrenia subjects.
Schizophrenic subjects with unconjugated hyperbilirubinemia show a significant enlargement of cerebral ventricles[30]and abnormalities of brain metabolism compared to both normobilirubinemic schizophrenia patients and normal controls[31].Neuroinflammation in schizophrenia is characterized by increased serum concentrations of proinflammatory cytokines, including IL1β, IL6, and transforming growth factor-β(TGFβ) and microglial activation[32,33].This inflammatory picture is similar to that observed in kernicterus spectrum disorders, the neurological sequel due to severe neonatal hyperbilirubinemia[34].Dalman and Cullberg[35]reported that neonates experiencing severe neonatal hyperbilirubinemia (TSB > 15 mg/dL)might later have an increased frequency of mental disorders.This clinical hypothesis is supported by the finding in the Gunn rat, the animal model for severe neonatal hyperbilirubinemia[36], showing severe hyperbilirubinemia in the first weeks of life[37].The Gunn rat displays a stereotypical behaviour,a supposedly typical symptom of schizophrenia, associated with microglia activation (indicative of inflammation), decreased ribosomal protein synthesis activity in neuronal cells, decreased neurogenesis,and increased apoptosis[38,39].
On the other hand, multiple studies have reported lower TSB levels among schizophrenia patients compared to patients suffering from some other psychiatric disorder and healthy controls[40-42].To reconcile these contradictory findings, Víteket al.[42]evaluated the correlation of bilirubin level with variability of the promoter of the gene forUGT1A1(uridine-diphospho-glucuronosyltransferase 1A1) (responsible for hepatic conjugation of UCB) between schizophrenia patients and controls.They noticed that an increase in bilirubin of 1 mM (0.06 mg/dL) could reduce the odds for schizophrenia status up to 19%.
Notably, UGTs participate in the metabolism of dopamine[26], a critical neurotransmitter, the loss of which is responsible for motor PD, and dopaminergic over activity has recently been suspected responsible for psychosis in schizophrenia[43].An additional potential link between these two pathologies might be heme oxygenase-1 (HO-1).The overexpression of this enzyme in the astrocytes of glial fibrillary acidic proteinheme oxygenase-1 (GFAP-HMOX1) transgenic mice resulted in increased subcortical oxidative imbalance with the induction of mitochondrial damage and autophagy, augmented dopamine and serotonin levels in the basal ganglia, and reduced dopamine 1 receptor (D1) in the nucleus accumbens, and enzyme overexpression caused degeneration of axons in the hippocampus and hyperkinetic behavior[44].However,there is still no clear evidence of HO-1 alteration in the clinical setting for schizophrenia.
PD
PD is another neurological disease linked to UCB.A study comparing 420 PD patients and 435 healthy control showed not only a significant increase in bilirubin in PD patients but also a negative correlation between bilirubin level and progression from a less to more severe staging of the disease[45].Complementary with this study, Mocciaetal.[46]also reported higher TSB concentration in drug-naive PD patients compared to controls, and Qinet al.[47]also observed an upregulation of levels of direct bilirubin (conjugated bilirubin, in clinical terminology), accompanied by reduced UCB (indirect bilirubin) levels in PD patients compared to the healthy group.Notably, L-DOPA treatment (the most used therapeutic approach to PD,able to improve symptomatology temporarily), has been found to increase TSB by about 20%[48].Because L-DOPA may increase the oxidative stress causing dopaminergic neuron loss in PD, and since bilirubin is a well-known antioxidant, an increased bilirubin level has been interpreted as a possible protective response to the disease[48].
As noted in the Introduction, almost every cell in the body possesses the full enzymatic equipment for producing UCB itself[7][Figure 2].Modulation of the enzymes responsible for UCB production in the brain of subjects with neurological/neurodegenerative diseases has been reported, including PD patients.An increased HO-1 signal has been detected in reactive astrocytes and affected dopaminergic neurons showing Lewy bodies[49].Although, the authors suggested that the HO-1 induction was uniquely due to its antioxidant properties, it is clear now that the elevation of HO-1 levels also represents an attempt to downregulate inflammation[50].
A direct proof of the protective effect of HO-1 induction has been obtained in experimental models.The injection of adenovirus containing human heme oxygenase-1gene (Hmox1) into the substantia nigra of methyl-4-phenylpyridinium (MPP+) treated rats (a model for PD) revealed that the overexpression ofHmox1protects dopaminergic neurons by reducing the expression of TNF-a and IL1β, and increasing brain-derived neurotrophic factor (BDNF) and glial cell line-derived neurotrophic factor (GDNF)expression[51].Both growth factors, BDNF and GDNF, are able to prevent injury to dopaminergic neurons and improve the behavioural deficits in PD[52-54].A recent work based on organotypic cultures of substantia nigra (challenged with rotenone to induce PD) demonstrated that inflammation and redox imbalance are early and simultaneous triggers for dopaminergic neuron loss[55].In this study,Hmox1upregulation occurs only at the very first phases of neurodegeneration (3 hours after rotenone challenging, a time which represents the pre-diagnosis stage in human PD based on dopaminergic neuron loss).Similarly,Bdnfexpression initially increased but then rapidly declined below the control level accompanying dopaminergic neuron demise.The modulation trend of both markers (Hmox1andBdnf) was interpreted as a failed tentative reaction to the insult[55], supporting the hypothesis of the interplay between L-DOPA and bilirubin(see above).This interpretation may also agree with a recent finding of Songetal.[56], who reported that the expression levels of HO-1 in the saliva of PD patients with early-stage (stage 1) based on the Hoehn and Yahr scale were significantly higher compared to control subjects or PD patients at stage 3 (advanced PD).The authors proposed an additional value of HO-1 as a potential biomarker in idiopathic PD patients.Notably, hyperactivation of HO-1 may exacerbate oxidative stress by the deposition of iron (Fe), one of its products [Figure 2], a phenomena frequently observed in neurological lesions and known to worsen the disease[49].This aspect is discussed in detail later on in the review.
Contrary to PD and schizophrenia, other neurological conditions have been associated with a reduced TSB level.
MuItipIe scIerosis
MS is a chronic inflammatory disease characterized by the destruction of myelin in the brain and spinal cord likely due to loss of immune system tolerance to myelin[57].TSB levels have been reported to be lower (vs.healthy controls) also in MS subjects with clinically isolated syndrome (predominately by neuroinflammation) and relapsing-remitting multiple sclerosis (predominately by neurodegenerative disorder), where predominate neurodegenerative disorders[58].Interestingly, bilirubin levels have been shown to be significantly lower in relapsing-remitting multiple sclerosis compared to clinically isolated syndrome patients[59], suggesting a relationship between increased disease severity and decreased TSB.This hypothesis seems to be confirmed by the study of Ljubisavljevicet al.[59]who found a significant correlation between higher TSB and lower disability status, fewer MRI lesions, and shorter disease duration in both study groups.In the brain tissue from MS patients, enhanced nitrotyrosine staining, one of the markers of oxidative damage[60], was found in demyelinated regions, specifically in hypertrophic astrocytes and foamy macrophages on inflammatory lesions[61].The damaged sites also showed an upregulation of antioxidant enzymes, including HO-1, compared to normal-appearing white matter and white matter tissue from control brains with no neurological disease, and HO-1 immunoreactivity was particularly confined to microglia[61].These findings emphasize the close interaction between oxidative stress and inflammation in MS.
The effects of bilirubin on MS have also been studied in the animal model for the disease, the so-called experimental autoimmune encephalomyelitis (EAE) model, where the infiltration of lymphocytes, the activation of CD4+T cells, and death of oligodendrocytes lead to the destruction of myelin sheaths[62,63].Figure 3 illustrates the most relevant mechanisms of action of bilirubin in EAE, one of the best-known models unravelling the interplay between MS and bilirubin.Bilirubin administration in the EAE model effectively prevented both acute and chronic EAE, even better than did glucocorticoid treatment, the most commonly used therapy for MS[64,65].
Histological analyses demonstrated that bilirubin interfered with the infiltration of inflammatory cells into the CNS by protecting the blood-brain barrier from free radical-induced permeability changes[64],stressing the intimal connection between redox stress and inflammation in CNS diseases.Further study supported the antiinflammatory potential of bilirubin in the EAE model[64].In vitroexperiments using spleen-harvested CD4+T cells showed that bilirubin at non-apoptotic concentrations (20-150 mM)inhibits CD4+T cell proliferation through various mechanisms.It suppresses the production of T helper-1(Th1) cytokines (IL2 and IFNγ) and Th2 cytokines (IL4 and IL10), reduces costimulatory molecule activity (CD28 on CD4+T cell, the co-receptor B7-1 activity in macrophages and dendritic cells), inhibits NFkB activation, which is a key transcription factor involved in T cell receptor-mediated signaling, and downregulates inducible MHC (major histocompatibility complex) class II expression.Bilirubin effectively downregulates EAE in SJL/J mice as confirmed by the reduction of the proliferation capacity of CD4+T cell.Meanwhile, the reduction of endogenous bilirubin synthesis by zinc-protoporphyrin, a specific inhibitor of the bilirubin producing-enzyme HO-1, dramatically exacerbates this disease[66].In contrast, induction of HO-1 by cobalt protoporphyrin IX (CoPPIX) inhibits EAE effectively[67].
According to the previous data, it seems beneficial to increase bilirubin synthesis through HO-1 induction.Several clinically used drugs have been reported to induce HO-1, among them are nonsteroidal antiinflammatory drugs (e.g., coxibs, acetylsalicylic acid) and hypolipidemic agents (e.g., niacin, fibrates,statins)[68].Atorvastatin and rosuvastatin treatment in mice demonstrated protection by increasing not only plasma bilirubin concentrations (up to 70%) but also cardiac tissue bilirubin content (up to 119%)[69].On the contrary, HO-1 induction in astroglia promotes oxidative mitochondrial membrane damage, iron sequestration, and mitophagy (macroautophagy)[56].These reasons then increase the doubt of bilirubin synthesis through HO-1 induction as a good strategy to counteract the neuroinflammatory process.
In another part of inflammation signalling pathways, both UCB and biliverdin are known as activators of aryl hydrocarbon receptor (AhR), a ligand-activated transcription factor that plays critical modulatory roles in various immune cells during innate and adaptive immune responses[2,70,71].AhR has critical roles in MS by modulating IL17 producing T-helper (Th17) cells, and regulatory T cells[72-74], as well as B cells,macrophages and dendritic cells.In macrophages, AhR regulates IL1β production and also IL6, IL12 and TNFa expression[73,75].AhR also inhibits the transcriptional activity of NFκB in stimulated macrophages[76].Meanwhile, in dendritic cells, AhR mediates the generation of T regulatory cells and Th17 cells from naive T cells and also IL10 production[77].Notably, the injection of low doses of UCB (20 μg/kg body weight,injected into C57Bl/6 mice) increased the population of regulatory T cells by more than 50% compared to controls, prolonging graft survival[78].Regulatory T cells are also inducible by HO-1 activity, CO, and UCB[78]and all “yellow players” [Figure 2].
Optic neuritis and neuromyeIitis optica
Reduced TSB has been noticed in both optic neuritis, an acute inflammatory demyelinating disorder of the optic nerve, and neuromyelitis optica, also known as Devic disease, a severe autoimmune demyelinating disease that selectively affects the optic nerve and spinal cord[79-82].Although the main focus was on the potential antioxidant role of bilirubin, suggesting that the low TSB is a result of the overconsumption of bilirubin due to oxidative stress, immunohistochemistry of damaged nerves is similar as in MS[83], with an evident inflammatory component.Thus, the interplay between bilirubin and inflammation described in MS also applies to optic neuritis and neuromyelitis optica.Further studies are needed.
Diabetic retinopathy and stroke
TSB levels have been found to be decreased also in patients with diabetes mellitus and diabetic retinopathyvs.subjects with diabetes mellitus without retinopathy, suggesting TSB as a biomarker of diabetic retinopathy[84].In the pathogenesis of diabetic retinopathy, particularly in the non-proliferative type,hyperglycemia exposure causes the alteration of retinal microvasculature with pericyte loss[85], endothelial apoptosis and endothelial thickening of the basement membrane, which further lead to capillary occlusion and ischemia[86].
Similarly, damage and even destruction of the cerebral vasculature are landmarks of stroke and related vascular events[87].In a large study conducted by Liet al.[88], 343 subjects with silent cerebral infarct presented with lower TSB levels.In another study, patients with deep white matter lesions (DWMLs) had a decreased TSB level compared to non-DWMLs subjects.Again, the low and intermediate bilirubin groups showed a higher prevalence of severe DWMLs than did the group with higher bilirubin levels[89].Notably,DWMLs are a recognized predictor for the development of impaired cognitive function and stroke[90,91].
A recent study also explored the association of UCB and intracranial atherosclerosis, which was found in approximately 50% of patients with transient ischemic attack and up to 47% of ischemic stroke patients in Asia[92-94].In this study, UCB was significantly negatively associated with intracranial atherosclerosis.The odds of intracranial atherosclerosis was 0 0.67-fold lower in participants in the high UCB concentration group (≥ 10.10 mMol/L) when compared with those in the low UCB concentration group[94].An increased bilirubin level has been proposed as a novel independent predictor for hemorrhagic transformation and symptomatic intracranial hemorrhage after mechanical thrombectomy[95].
The role of bilirubin in all previously mentioned conditions might be explained by a large number of studies suggesting bilirubin as a protector against microvascular complications.Bilirubin exhibits potent antiinflammatory effects [via HO-1, nuclear factor erythroid 2-related factor 2 (Nrf2) and NFκB] by the inhibition of monocyte transmigration [through a decrease in TNFa-induced monocyte chemoattractant protein-1 (MCP-1) secretion], reducing endothelial vascular cell adhesion molecule-1 (VCAM-1)expression, and improving endothelial cell dysfunction and hyperproliferation after damage[96-99].UCB also appears to affect the immune system by inhibiting the activation of the complement cascade[100], and also by modulating the phagocytic and antigen-presenting function of macrophages (by changing the expression of Fc receptors)[101].
The potential protective role of HO-1 induction has been demonstrated in several studies.HO-1 increases early (24 h) after brain trauma[102], and in intracerebral haemorrhage models, after infiltration of the brain by blood components activating the inflammatory response[103].HO-1 activity is observed in both endothelial cells and microglia surrounding the haematoma site.The protective effects of HO-1 in neurons was studied by Orozco-Ibarraet al.[104], who revealed that the upregulation of HO-1 prevented the death of cerebellar granule neurons due to mitochondrial toxicity.Further investigation found that the HO-1 products, bilirubin and CORM2 (carbon monoxide releasing molecule), were involved in preventing cell death.
HO-1 has neuroprotective effects by regulating the phosphatidylinositol 3-kinase (PI3K/AKT) signalling pathway and by reducing apoptosis in rats with cerebral haemorrhage[105].Fenget al.[106]showed that the PI3K/AKT and extracellular signal regulated kinase pathways are involved in oleanolic acid induced HO-1 expression by activating Nrf2 in vascular smooth muscle cells.As one of the signalling pathways for cell survival, the PI3K/AKT signal transduction pathway plays an important role in cell proliferation and differentiation and inhibition of neuronal apoptosis[107].Therefore, HO-1 may protect the nerves of rats with cerebral haemorrhage by regulating the PI3K/AKT signalling pathway.
A second isoform of HO exists in the brain, called heme oxygenase-2 (HO-2).Considered constitutive, it looks like HO-2 in the brain plays the vital function of maintaining adequate levels of UCB to guarantee normal cellular homeostasis, participating in brain protection[108].The protective activity of HO-2 has been reported in intracerebral haemorrhage models, where heme oxygenase-2 gene (Hmox2) deletion led to greater brain injury volumes and neurological deficits than in wild-type mice after intracerebral haemorrhage[109].In brain cultures, Doréet al.[108]expanded our knowledge on the role of HO-2 and bilirubin as neuroprotective factors by showing increased neuronal death in cerebellar granule cultures ofHmox2knockout mice (Hmox2-/-).On the contrary,Hmox2transfection in human embryonic kidney 293 (HEK293) cells rescued cells from apoptotic death.In another study, the induction of HO-2 activity by phorbol esters enhanced the production of bilirubin, which protected primary hippocampal and cortical neuronal cultures from the neurotoxicity of H2O2.Of note, HO-2 was immunolocalised in neurons both before and after traumatic brain injury, whereas HO-1 was highly upregulated in glia only after traumatic brain injury.Cell loss was significantly greater inHmox2-/-mice in areas including the cortex,hippocampus and lateral dorsal thalamus[110].
Biliverdin (BV), another yellow player, also showed neuroprotective effects by ameliorating cerebral reperfusion injury in rats most probably via its antiinflammatory activity[111].Two pathways are known to be involved in the antiinflammatory mechanism of BV: by activating the nitric oxide-dependent biliverdin reductase, BV reduces the expression of toll-like receptor-4 (TLR-4) in murine macrophages[112], and BV regulates the expression of complement C5a receptor[113].Furthermore, a study by Zouet al.[111]found the downregulation of miR-204-5p and its target gene, ETS protooncogene 1 (Ets1), in cerebral ischemiareperfusion injury rats following BV administration.Ets1 is known to be responsible for inducing proinflammatory Th1 type response and causing neuronal death[114].Therefore, BV may play a crucial role in preventing injury in stroke by interfering in miRNAs levels.
AdditionaI neuroIogicaI conditions underpinned by reduced TSB IeveIs
ALS
In ALS, the reduction of TSB levels correlates with both the clinical state and disease duration.Patients with long-lasting ALS (where motor neuron degeneration is noticed) have lower TSB levels than do patients with a shorter duration[115].
Seasonal affective disorder
Nocturnal plasma bilirubin levels were evaluated in individuals with unipolar recurrent winter seasonal major depressive disorder and age-/gender-matched controls.Lower bilirubin levels were found in patients with seasonal affective disorder.The reduction in bilirubin level in this study was proposed as a vulnerable risk for depression, although only an associative and not a causative link was provided[116].
Dementia
A significant reduction in TSB levels was reported in patients with cognitive impairment compared to those with normal cognitive function, although no significant correlation between bilirubin and disease duration was found[117].In a further study by Baierleet al.[118], the authors showed that the significant increase in the proinflammatory cytokines IL1β and TNFa correlated with the increase in oxidative stress and inversely associated with low cognitive performance in elderly patients.
AD
As in the majority of neurological conditions, TSB levels are also decreased in AD[119].The lower concentration of bilirubin is supposed to have a role in the transition from mild cognitive impairment to clear AD[120].Similarly, reducedHmox1mRNA[121]and protein[122]levels were detected in the serum of fullblown AD subjects, and proposed to correlate with cognitive decline[120].Notably, the diagnosis of AD and cognitive decline is usually a late event and performed after the pathological alteration has already existed for some time.In the early stages, the brain tries to protect itself from the oxidative and inflammatory insult of AD by increasing the endogenous production of UCB.Both HO-1 and biliverdin reductase A(BLVR-A), the two key enzymes in biliverdin and bilirubin production [Figure 2], have been reported to be upregulated in the brain of AD patients[103,123].However, the upregulation of HO-1 will also cause Fe deposition in the brain, which worsens and accelerates disease progression by enhancing the redox imbalance in ongoing AD lesions[49,124-126].It should be recalled that the brain possesses very low antioxidant capacity compared to other organs, being much lower in neonatal life[127,128]and in the elderly[129,130].Thus, the enhanced pro-oxidant milieu due to HO-1 hyperactivation leads to an increased oxidative and nitrosative post-translational modification of cellular enzymes, with their consequent inactivation[131].One of the targets of this mechanism is BLVRA, and thus, UCB production will be stopped[123].
The chains of events described here, leading to the disruption of UCB-mediated protection and contributing to the progression of neurological damage fits well with what we described in PD.
IS THE BRAIN DIFFERENT FROM THE REST OF THE BODY?
Differently from extra-CNS diseases involving oxidative imbalance and inflammation, where a positive correlation between higher TSB level (Gilbert syndrome-like) and reduced disease incidence has been frequently found[2,3,68], the most readily evident finding in this review is that brain diseases have lower TSB levels in common.Two major explanation have been given: (1) lower TSB levels in subjects with neurological disease may reflect the consumption of UCB due to oxidative stress, although it seems unlikely that the local redox imbalance occurring in the brain may affect the systemic levels of bilirubin; and (2)individuals with lower TSB, thus a lower systemic antioxidant status, might be more vulnerable to oxidative stress[118]and related diseases (both in the brain and extra-CNS organs) [Figure 1].
Experimental evidence strongly supports the notion that an increased concentration of bilirubin in the CNS exerts antiinflammatory and antioxidant effects.On the other hand, it should be recalled that hyperactivation of HO-1 in the CNS, reported to be a tentative reaction against brain insults, may enhance the damage[68,104,131-148].This specificity of the CNS has to be taken into account in the growing research aimed at modulating the bilirubin players to increase levels of bilirubin and the protection it confers.
CONCLUSION
Studies assessing clearly the molecular events correlating bilirubin and neurological damage are still lacking, and many conclusions are based on clinical series.Understanding the role of bilirubin and all the other yellow players in the development and progression of the different neurological disorders may help in deciphering the efficacy of the modulation of bilirubin level to prevent CNS diseases.However, due to the side effects of the drugs inducing HO-1 activity, caution must be taken before their recommendation.More intriguing and promising is thein situCNS elevation of UCB level achievable by the development of molecules modulating BLVR.The future will hopefully provide the much-needed answer due to the medical and social burden of neurological disorders.
DECLARATIONS
Authors' contributions
Made substantial contributions to the search of the literature, the writing of the manuscript, and the revision and discussion of the contents of the paper: Jayanti S, Moretti R, Tiribelli C, Gazzin S
AvaiIabiIity of data and materiaIs
Not applicable.
FinanciaI support and sponsorship
Jayanti S was supported in part by a fellowship from the Lembaga Pengelola Dana Pendidikan of Ministry of Finance of Indonesia and an internal grant from the Italian Liver Foundation.The funders had no role in data analysis and interpretation and also writing of the manuscript.
ConfIicts of interest
All authors declare that there are no conflicts of interest.
EthicaI approvaI and consent to participate
Not applicable.
Consent for pubIication
Not applicable.
Copyright
© The Author(s) 2020.
 Neuroimmunology and Neuroinflammation2020年2期
Neuroimmunology and Neuroinflammation2020年2期
- Neuroimmunology and Neuroinflammation的其它文章
- Favorable and unfavorable roles of microglia and macrophages in the pathologic central nervous system
- COVID-19 neurotropism and implications for therapy
- An association between mitochondria and microglia effector function: what do we think we know?
- Phf15 - a novel transcriptional repressor regulating inflammation in a mouse microgIiaI ceII Iine
- AUTHOR INSTRUCTIONS
- CerebrospinaI fluid amyIoid beta and tau proteins in atypicaI Parkinsonism: a review