
不同类型稻田中全程氨氧化微生物的分异特征
2020-07-23 16:37张洁宋怡轩张鑫磊张耀鸿
江苏农业学报 2020年3期
关键词:稻田
张洁 宋怡轩 张鑫磊 张耀鸿

摘要:選用微碱性氮贫瘠的上海市崇明岛稻田和微酸性氮丰富的南京市稻田剖面(0~50 cm),比较研究稻田土壤中氨氧化微生物类群丰度的差异及其环境驱动机制,评价其与氨氧化潜力的内在关系。结果表明,崇明稻田的净硝化速率为12.82~22.30 mg/(kg·d),明显高于南京稻田[4.26~7.46 mg/(kg·d)]。崇明稻田土壤剖面的Comammox amoA基因总拷贝数(Clade A与Clade B之和)均值为1 g 8.8×106拷贝,是南京稻田的2.4倍,且Clade A与Clade B的比值范围为2.5~12.7,证实了Comammox存在于2种不同类型的稻田土壤中。崇明稻田和南京稻田剖面的氨氧化细菌(AOB)的amoA基因拷贝数均值分别为1 g 3.75×108拷贝和1.23×108拷贝,氨氧化古菌(AOA)的amoA基因拷贝数均值分别为1 g 2.05×107拷贝和0.35×107拷贝,这2种菌群基因拷贝数均在10.1~20.0 cm土层达到最高。回归分析发现,2个稻田中氨氧化细菌(AOB)对氨氧化潜力的贡献率达到90%~94%,而Comammox仅为3%左右,表明氨氧化细菌(AOB)在氨氧化过程中发挥主要作用。
关键词:稻田;全程氨氧化细菌;氨氧化细菌;氨氧化古菌;氨氧化潜力
中图分类号:S511.061文献标识码:A文章编号:1000-4440(2020)03-0584-07
Differentiation characteristics of complete ammonia-oxidizing microorganisms in different types of paddy soils
ZHANG Jie 1,2,SONG Yi-xuan 1,2,ZHANG Xin-lei 1,3,ZHANG Yao-hong1
(1.Nanjing University of Information Science and Technology, Collaborative Innovation Center for Forecast and Evaluation of Meteorological Disasters/Jiangsu Provincial Key Laboratory of Agricultural Meteorology, Nanjing 210044, China;2.State Key Laboratory of Soil and Sustainable Agriculture, Institute of Soil Science, Chinese Academy of Sciences, Nanjing 210008, China;3.Shanxi Climate Center, Taiyuan 030006, China)
Abstract:In this study, the alkaline coastal paddy field profile (0-50 cm) with low nitrogen and acidic Nanjing paddy field profile with high nitrogen were selected to compare the abundance of ammonia-oxidizing microorganisms in paddy soils and its environmental driving mechanism, and to evaluate its internal relationship with ammonium oxidation potential was evaluated. The results showed that the net nitrification rate of coastal paddy fields ranged from 12.82 mg/(kg·d) to 22.30 mg/(kg·d), which was significantly higher than that of Nanjing paddy fields [4.26-7.46 mg/(kg·d)]. The average number of total copies of comammox amoA gene (the sum of Clade A and Clade B) in the soil profile of coastal paddy fields was 8.8×106 copies per gram, which was 2.4 times greater than that of Nanjing paddy fields, confirming that comammox existed in two different types of paddy fields. In addition, the ratio of Clade A to Clade B ranged from 2.5 to 12.7. The average copy number of amoA gene of ammonia-oxidizing bacteria(AOB) in coastal and Nanjing paddy fields was 3.75×108 copies per gram and 1.23×108 copies per gram, respectively. The average copy number of amoA gene of ammonia-oxidizing archaea (AOA) was 2.05×107 copies per gram and 0.35×107 copies per gram, respectively. The abundance of these two bacteria were the highest in 10.0-20.0 cm soil layer. The results of regression analysis indicated that AOB contributed 90%-94% to the potential of ammonia oxidation in the two paddy fields, while comammox only contributed about 3%, indicating that AOB could play a major role in ammonia oxidation in the two soils.
Key words:paddy soils;complete ammonia oxidizers(comammox);ammonia-oxidizing archaea(AOA);ammonia-oxidizing bacteria(AOB);potential of ammonia oxidation
硝化作用是自然界氮循环的关键转化过程,也是农田土壤氮素循环的重要环节[1]。氨氧化作用是硝化过程的第一步反应,也是整个过程的限速步骤。一直以来,普遍认为变形菌纲的氨氧化细菌(AOB)是其唯一承担者,直到氨氧化古菌(AOA)被发现[2],才证实了氨氧化过程是由AOB 和AOA 两类微生物共同承担。
根据经典动力学理论计算,一种微生物独自将NH3氧化为亚硝酸盐,再继续氧化为硝酸盐的两步氧化过程是可行的[3]。2015年底,科学家在水生生态系统中发现了全程氨氧化微生物(Complete ammonia oxidizers,Comammox),能够直接完成从NH+4到NO-3的氧化[4-5]。新近的研究结果证实,Comammox广泛存在于农田土壤、自然湿地、河流沉积物、废水污泥和饮用水处理系统中[4-7]。但是,Comammox对氨氧化速率的贡献以及与传统的AOA和AOB如何相互竞争/共存及其相对重要性,仍不清楚,是目前研究的热点。
稻田土壤因受到施肥、淹水、农业耕作等因素的影响,其氨氧化及硝化作用非常强烈。有报道认为,稻田土壤中NH3浓度的变化可能是驱动Comammox 和AOA及AOB的关键环境因子[8]。受农业施肥的影响,稻田土壤中的活性N水平差异极大;且施肥往往会导致土壤酸化,深刻影响着活性N尤其是NH+4的存在形态。受H+介导的化学平衡影响,酸性土壤中NH3浓度通常远低于中性和碱性土壤[9]。经典AOA和AOB的氨单加氧酶只能催化非离子态氨(NH3),而不能催化离子态铵(NH+4)[10];而Comammox是否也只能氧化非离子态氨仍不清楚。许多研究者认为,AOA对NH3的亲和力远高于AOB[11]。因此AOA 通常主导酸性土壤氨氧化[12],而AOB 则在中性和碱性土壤中发挥作用[13-14]。而最新的研究结果则表明,全程氨氧化细菌对NH3的亲和力远高于大多数可培养的AOA,能够更好地适应于极低NH3浓度的胁迫环境[15]。然而,在不同pH值、不同活性N水平的稻田土壤环境中,Comammox和AOA、AOB对稻田土壤氨氧化速率的相对贡献仍不清楚。
因此,本试验选取2个不同类型的稻田——微碱性N贫瘠的长江口崇明稻田和微酸性N丰富的南京稻田,研究这2个稻田土壤剖面的氨氧化潜势及各类氨氧化微生物的丰度,揭示两者之间的内在关系。
1材料与方法
1.1土壤样品采集
采样地点为上海市崇明岛围垦稻田(31.50°N, 121.92°E)和南京信息工程大学农业气象试验站稻田(32.16°N, 118.86°E)。两个稻田都处于北亚热带湿润气候区,年均降水量1 000~1 100 mm,年均气温15.6 ℃。在每个稻田内以S形设置6个采样点,各采样点间距约为10~15 m,用土钻取 0~10.0 cm、10.1~20.0 cm、20.1~35.0 cm、35.1~50.0 cm的土层鲜土,并将相同深度土层的土样充分混合,放入冰盒中运回实验室冷冻保存备用。
1.2土壤理化性质测定
土壤全氮(TN)含量采用半微量凯氏定量法测定。土壤总有机碳(TOC)含量采用浓硫酸-重铬酸钾消煮-硫酸亚铁滴定法测定。土壤铵态氮和硝态氮含量用2 mol/L KCl溶液浸提后,采用AA3流动分析仪测定。土壤pH采用水土比为2.5∶1.0(质量比)提取水溶液后,用数字酸度计测定。
用净硝化速率反映稻田剖面土壤的好氧氨氧化潜力。净硝化速率的测定采用好气培养法。取沉积物土样10 g(干基),加15 ml去离子水,将土样分散均匀,形成悬浊液。称取总质量并记录。25 ℃避光预培养7 d以便激活土壤微生物。预培养结束后,将每个土样分为2组:第1组的3个锥形瓶,直接测定正式培养起始时刻(t0)的硝态氮浓度。第2组的6个锥形瓶中,用移液枪加入(NH4)2SO4溶液使N最终浓度达到200 mg/L,再根据质量法补加去离子水,使之去离子水总量为20 ml,然后在恒温培养箱中25 ℃避光培养7 d(t)后,取出锥形瓶,测定其中的硝态氮浓度。硝化速率的计算公式为:
N=[(NO-3)t-(NO-3)t0]/(t-t0),式中,N为硝化速率,单位为mg/(kg·d),(NO-3)t和(NO-3)t0 分别为t d和0 d时NO-3-N含量。
1.3土壤DNA提取
利用Fast DNA Spin kit for soil提取试剂盒 (MP Biomedicals, USA) 提取土壤样品中的总DNA。取0.5 g土壤样品,按试剂盒说明书的步骤提取DNA,取部分DNA提取液用分光光度计(NanoDrop -1000 UV-Vis)测定DNA浓度和纯度。土壤DNA保存于-80 ℃冰箱待用。
1.4实时荧光定量PCR
在C1000TM Real-Time System扩增仪上进行实时荧光定量PCR扩增。测定氨氧化古菌(AOA)、氨氧化细菌(AOB)和全程氨氧化细菌分支A与分支B(Comammox Clade A与Comammox Clade B)的amoA基因拷贝数。AOB、AOA、Comammox的 amoA基因定量 PCR 所用擴增引物和扩增条件参照文献[16]、[17]和[18]。反应体系均为20.0 μl,包括DNA样品1.0 μl、Taq DNA聚合酶10.0 μl、前后引物各0.5 μl、无菌水8.0 μl。提取氨氧化微生物的amoA基因的重组质粒,并进行测序验证,再用分光光度计( NanoDrop ND-1000 UV-Vis)测定质粒浓度,并用无菌水将质粒稀释6~8个梯度,用于制作定量PCR的标准曲线,根据标准曲线计算目的基因的拷贝数。
1.5数据统计分析
用SPSS 16.0软件进行统计分析,通过单因素方差分析(one-way ANOVA)和Pearson分析进行土壤理化性质、氨氧化微生物丰度的差异性以及相关性检验,显著性水平α=0.05。
2结果与分析
2.1不同类型稻田土壤的理化性质
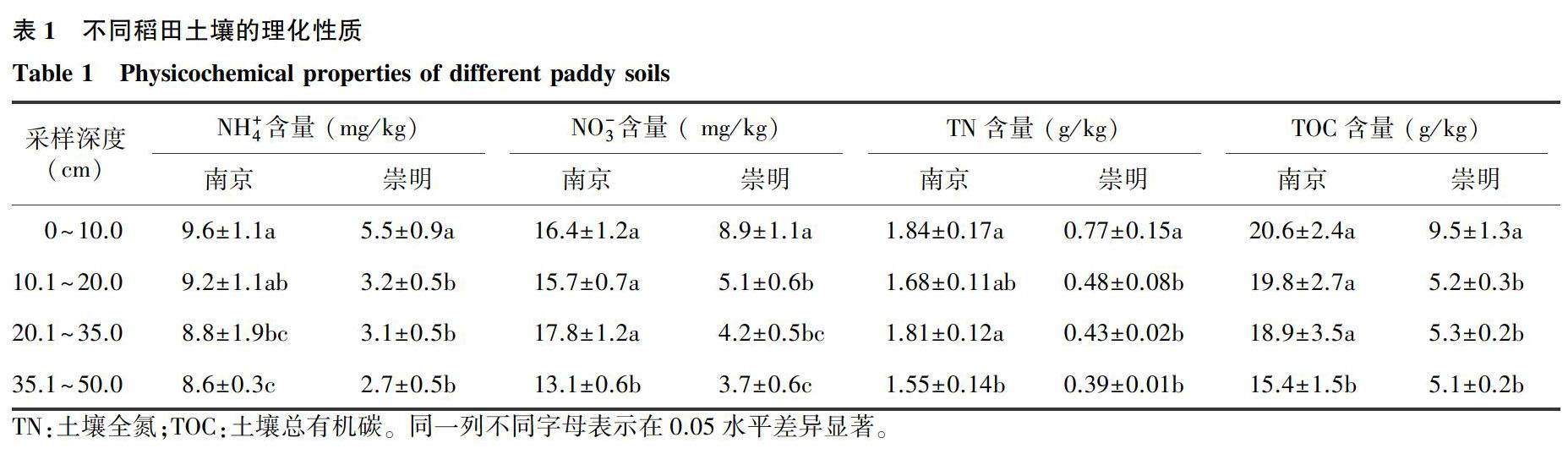
南京稻田和崇明稻田剖面的土壤理化性质如表1所示。南京稻田土壤剖面中NH+4和NO-3含量范圍分别为8.6~9.6 mg/kg和13.1~17.8 mg/kg,明显高于崇明稻田土壤剖面,且随土壤深度的变化较小。而崇明稻田剖面中NH+4和NO-3含量变化幅度较大,表现为0~10.0 cm土层的硝态氮含量显著高于底层,其垂直分布特征与南京稻田土壤明显不同。南京稻田的TN和TOC均值分别为1.72 g/kg和18.68 g/kg,分别是崇明稻田的3.3倍和2.9倍。南京稻田剖面的pH值为6.3~6.5,崇明稻田剖面的pH值为7.6~7.9。总体而言,南京稻田为微酸性氮丰富土壤,且活性氮和总氮含量随剖面深度增加而变化较小;崇明稻田为微碱性氮贫瘠土壤,且表层土壤活性氮和总氮含量显著高于底层土壤。这为进一步深入分析两个稻田的氨氧化过程提供了很好的试验材料。
TN:土壤全氮;TOC:土壤总有机碳。同一列不同字母表示在0.05水平差异显著。
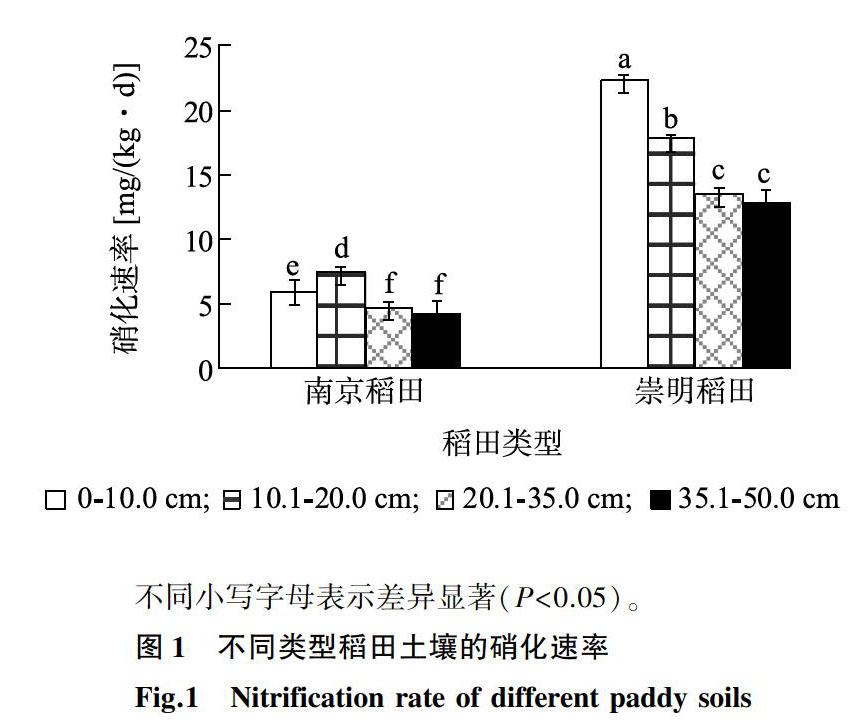
2.2不同类型稻田土壤的硝化速率
图1显示,南京稻田的硝化速率为4.26~7.46 mg/(kg·d),其中,10.1~20.0 cm土层的土壤硝化速率显著高于深层。崇明稻田的硝化速率为12.82~22.30 mg/(kg·d),表现为表层硝化速率最高,随深度增加而逐渐减小。相关性分析发现,两个稻田的硝化速率与土壤氨态氮含量呈显著正相关,相关系数分别为0.66*和0.92*,与土壤TOC也呈显著正相关,相关系数分别为0.70*和0.87*。
不同小写字母表示差异显著(P<0.05)。
2.3不同类型稻田土壤的氨氧化细菌和古菌丰度特征
由图2可知,南京稻田和崇明稻田的氨氧化古菌(AOA)的amoA基因拷贝数(以干土计)均值分别为1 g 0.35×107拷贝和2.05×107拷贝,其中,在10.1~20.0 cm土层最高,分别为1 g 0.82×107拷贝和4.18×107拷贝。两个稻田土壤的氨氧化细菌(AOB)的amoA基因拷贝数均值分别为1 g 1.23×108拷贝和3.75×108拷贝,其中,也在10.1~20.0 cm土层达到最高,分别为1 g 2.41×108拷贝和6.21×108拷贝。通过相关性分析发现,崇明稻田剖面中氨氧化古菌(AOA)与氨氧化细菌(AOB)的amoA基因拷贝数显著正相关,相关系数为0.92*。
不同小写字母表示差异显著(P<0.05)。
2.4不同类型稻田土壤的全程氨氧化细菌丰度特征
由图3可知,两个稻田土壤中全程氨氧化细菌Comammox Clade A的丰度差异很大。南京稻田和崇明稻田土壤的Comammox Clade A amoA基因拷贝数均值分别为1 g 0.29×107拷贝和0.87×107拷贝,其中,在10.1~20.0 cm土层达到最高,分别为1 g 0.73×107拷贝和1.97×107拷贝。两个稻田剖面的Comammox Clade B amoA基因拷贝数均值分别为1 g 0.75×106拷贝和1.04×106拷贝,其中,也在10.1~20.0 cm土层达到最高,分别为1 g 2.15×106拷贝和2.22×106拷贝。通过相关性分析发现,两个稻田剖面中全程氨氧化细菌Comammox Clade A与Comammox Clade B的amoA基因拷贝数均呈显著正相关,相关系数分别为0.98*和0.94*。
2.5不同类型稻田土壤氨氧化微生物拷贝数比较及对氨氧化速率的贡献
对南京稻田和崇明稻田土壤的氨氧化微生物拷贝数进行比较。结果(表2)表明,两个稻田中氨氧化古菌(AOA)与全程氨氧化细菌的比值分别为1.54和2.32,说明两者的拷贝数为同一数量级。相反,氨氧化细菌(AOB)与全程氨氧化细菌的比值均大于40,而且氨氧化细菌(AOB)与氨氧化古菌(AOA)的比值也均大于10,说明3类氨氧化微生物中,氨氧化细菌(AOB)的数量占有绝对优势。值得注意的是,两个稻田中全程氨氧化细菌Clade A与Clade B的比值均高于1.0,说明Clade A在全程氨氧化细菌中数量上占有明显优势。
将各类氨氧化微生物amoA基因拷贝数与硝化速率进行线性回归分析(表3),发现南京稻田和崇明稻田中,随土壤硝化速率的增加氨氧化细菌AOB的响应度均最大,其线性方程的斜率分别达到7.67和2.54;其次是氨氧化古菌(AOA),响应度最小的是全程氨氧化细菌Clade A和Clade B。根据响应方程进一步计算氨氧化微生物对氨氧化速率的贡献率,结果显示两个稻田中氨氧化细菌(AOB)对硝化速率的贡献率达到90%~94%,而全程氨氧化细菌(Clade A+Clade B)的贡献率仅为2.9%~3.4%。
不同小写字母表示差异显著(P<0.05)。
AOA:氨氧化古菌;AOB:氨氧化细菌;Comammox Clade A:全程氨氧化细菌分支A;Comammox Clade B:全程氨氧化细菌分支B。
AOA、AOB、Comammox Clade A、Comammox Clade B见表2注。
3讨论
已有研究结果表明,全程氨氧化微生物可能广泛分布于土壤、沉积物、水等生态环境中[4-7]。本研究在微酸性氮丰富的南京稻田和微碱性氮贫瘠的崇明稻田土壤中,均检测出高数量级的全程氨氧化细菌(Comammox)amoA基因拷贝数,分支A(Comammox Clade A)与分支B (Comammox Clade B)amoA 基因拷贝数分别为1 g 0.5×106 ~2.0×107拷贝和1.1×105 ~2.2×106拷贝,证实了全程氨氧化细菌与氨氧化细菌(AOB)和氨氧化古菌(AOA)一样,广泛分布于不同类型的稻田土壤中。而且,两个稻田土壤中Comammox amoA基因拷贝数(分支A 与分支B 之和)均值与AOA的数量级相同,说明全程氨氧化细菌可能在这两种类型的水稻土硝化过程中发挥着一定的作用。
一般认为,氨氧化过程是硝化反应的第一步,也是限速步骤[19];氨氧化过程的反应底物为非离子态氨(NH3)而不是离子态铵(NH+4)[10],因此氨氧化过程是对pH高度敏感的典型生物学过程[20]。酸性土壤中NH3质量分数很低,所以一直以来科学家们认为硝化过程不能在酸性土壤中发生。但随后越来越多的证据表明酸性土壤中也存在硝化作用[21],但普遍低于中性或碱性土壤。本试验中,微酸性南京稻田土壤的硝化速率为5.58 mg/(kg· d),仅为微碱性崇明稻田的34%,原因可能是pH值导致南京稻田中硝化底物NH3含量降低而影响氨氧化作用,最终硝化速率减小。此外,已有许多研究结果表明,AOB 对氮肥更敏感,高氮条件下土壤中氨氧化过程的主要承担者是AOB而非AOA[13-14]。本研究发现,两个稻田土壤中,氨氧化细菌(AOB) amoA 基因拷贝数比AOA高出一个数量级,而且对硝化速率的贡献率高达90%~94%,表明本试验稻田中氨氧化速率的主要贡献者是AOB,而非AOA。
已有研究结果表明,全程氨氧化细菌(Comammox)可能更适应低氮的贫营养环境。Kits 等通过氨氧化的动力学分析发现,全程氨氧化细菌(Comammox)生长的氨底物Km值仅为0.05 μmol/L,远低于氨氧化古菌的Km值(0.7~4.4 μmol/L),表现出极强的氨底物捕获能力[15]。此外,Palomo 等发现[22],全程氨氧化细菌(Comammox)能编码低氧、磷饥饿下高效表达的基因phoD。本试验中,尽管检测到全程氨氧化细菌(Comammox)存在于两个稻田中,但对氨氧化潜力的贡献率仅为3%左右。Wang等[23]对长期施肥19年农田中的全程氨氧化细菌进行分析,发现全程氨氧化细菌对土壤氨氧化速率和亚硝酸盐还原速率的贡献极小,认为Comammox对底物NH3具有高亲和力,而氨氧化速率很低,非常适应于氨浓度极低的贫营养环境中,但在长期施肥的农田中,氨浓度远高于这类微生物的需求量,其数量和酶促速率不随施肥量增加而增加,表现出较小的氨氧化贡献率。本试验的崇明稻田母质尽管是河口沉积物,土壤养分较为贫瘠,但该围垦稻田已有20年左右的种稻施肥历史,其土壤全氮和活性氮含量明显高于海堤之外的自然滩涂湿地[24]。因此,与其他养分贫瘠生态系统相比,两个稻田中长期施肥导致的土壤氮含量增加,可能是全程氨氧化细菌(Comammox)对氨氧化潜力贡献率降低的重要原因之一。
本研究中还发现,两个类型稻田土壤中,全程氨氧化细菌(Comammox)Clade A与Clade B amoA基因拷贝数的比值均大于1.0,最高达12.7。Pjevac 等在意大利水稻土中检测到全程氨氧化细菌分支A(Comammox CladeA)的丰度明显高于分支B (Comammox Clade B)[25],与本试验结果一致。王梅等报道在重庆中性紫色水稻土中全程氨氧化细菌以分支A(Comammox Clade A)为主,且Comammox Clade A 对施肥更敏感,随施肥量增加而增加,而Comammox Clade B 却无显著变化[7]。本试验中两个类型稻田土壤中全程氨氧化细菌(Comammox)也是以分支A(Comammox Clade A)為主,对硝化速率的贡献可能远高于分支B(Comammox Clade B)。
参考文献:
[1]贺纪正,张丽梅. 氨氧化微生物生态学与氮循环研究进展[J].生态学报,2009,29(1):406-415.
[2]KNNEKE M,BERNHARD A E,DE LA TORRE J R,et al. Isolation of an autotrophic ammonia-oxidizing marine archaeon[J]. Nature, 2005, 437(7058):543-546.
[3]COSTA E, PREZ J, KREFT J U. Why is metabolic labour divided in nitrification?[J] Trends in Microbiology, 2006, 14(5): 213-219.
[4]DAIMS H, LEBEDEVA E V, PJEVAC P, et al. Complete nitrification by Nitrospira bacteria[J]. Nature, 2015, 528(7583): 504-509.
[5]VAN KESSEL M A H J,SPETH D R,ALBERTSEN M, et al.Complete nitrification by a single microorganism[J]. Nature,2015, 528(7583): 555-559.
[6]PALOMO A, JANE F S, GLAY A, et al. Metagenomic analysis of rapid gravity sand filter microbial communities suggests novel physiology of Nitrospira spp[J]. ISME Journal, 2016, 10(11): 2569-2581.
[7]王梅,王智慧,石孝均,等. 长期不同施肥量对全程氨氧化细菌丰度的影响[J]. 环境科学, 2018, 39(10): 4727-4734.
[8]曹彦强,王智慧,莫永亮,等. 施肥和淹水管理对水稻土氨氧化微生物数量的影响[J]. 土壤学报, 2019, 56(4): 1004-1011.
[9]PROSSER J I, NICOL G W. Archaeal and bacterial ammonia-oxidisers in soil: the quest for niche specialisation and differentiation[J]. Trends in Microbiology, 2012, 20(11): 523-531.
[10]SUZUKI I, DULAR U, KWOK S C. Ammonia and ammonium ion as substrate for oxidation by Nitrosomonas europaea cells and extracts[J]. Journal of Bacteriology, 1974, 120(1): 556-558.
[11]MARTENS-HABBENA W,BERUBE P M,URAKAWA H,et al. Ammonia oxidation kinetics determine niche separation of nitrifying Archaea and Bacteria[J]. Nature, 2009, 461(7266): 976-979.
[12]LU L, HAN W, ZHANG J, et al. Nitrification of archaeal ammonia oxidizers in acid soils is supported by hydrolysis of urea[J]. ISME Journal, 2012, 6(10): 1978-1984.
[13]JIA Z J, CONRAD R. Bacteria rather than Archaea dominate microbial ammonia oxidation in an agricultural soil[J]. Environmental Microbiology, 2009, 11(7): 1658-1671.
[14]XIA W, ZHANG C, ZENG X, et al. Autotrophic growth of nitrifying community in an agricultural soil[J]. ISME Journal, 2011, 5(7): 1226-1236.
[15]KITS K D, SEDLACEK C J, LEBEDEVA E V, et al. Kinetic analysis of a complete nitrifier reveals an oligotrophic lifestyle[J]. Nature, 2017, 549(7671): 269-272.
[16]YU C, HOU L, ZHENG Y, et al. Evidence for complete nitrification in enrichment culture of tidal sediments and diversity analysis of clade a comammox Nitrospira in natural environments[J]. Applied Microbiological Biotechnology, 2018, 102: 9363-9377.
[17]WANG M, HUANG G, ZHAO Z, et al. Newly designed primer pair revealed dominant and diverse comammox amoA gene in full-scale wastewater treatment plants[J]. Bioresource Technology, 2018, 270: 580-587.
[18]BEMAN J M, FRANCIS C A. Diversity of ammonia-oxidizing archaea and bacteria in the sediments of a hyper-nutrified subtropical estuary: Bahía del Tóbari, Mexico[J]. Applied and Environmental Microbiology, 2006, 72(12): 7767-7777.
[19]PROSSER J I. Autotrophic nitrification in bacteria. Advances in microbial physiology[J]. Advances in Microbial Physiology,1990,30(1):125-181.
[20]DE BOER W, KOWALCHUK G A. Nitrification in acid soils: micro-organisms and mechanisms[J]. Soil Biology and Biochemistry, 2001, 33(7): 853-866.
[21]DAHLGREN R A. Soil acidification and nitrogen saturation from weathering of ammonium-bearing rock[J]. Nature, 1994, 368(1): 838-841.
[22]PALOMO A, PEDERSEN A G,FOWLER S J, et al. Comparative genomics sheds light on niche differentiation and the evolutionary history of comammox Nitrospira[J]. The ISME Journal, 2018, 12: 1779-1793.
[23]WANG J, WANG J L, RHODES G, et al. Adaptive responses of comammox Nitrospira and canonical ammonia oxidizers to long-term fertilizations: Implications for the relative contributions of different ammonia oxidizers to soil nitrogen cycling[J]. Science of the Total Environment, 2019, 668:224-233.
[24]林黎,崔軍,陈学萍,等. 滩涂围垦和土地利用对土壤微生物群落的影响[J]. 生态学报, 2014, 34(4): 899-906.
[25]PJEVAC P, SCHAUBERGER C, POGHOSYAN L, et al. AmoA-targeted polymerase chain reaction primers for the specific detection and quantification of comammox Nitrospira in the environment[J]. Frontiers in Microbiology, 2017, 8: 1508-1514.
(责任编辑:张震林)
猜你喜欢
青年文学家(2020年22期)2020-08-31
澳门月刊(2020年7期)2020-07-14
农产品市场周刊(2019年21期)2019-12-20
农产品市场周刊(2019年16期)2019-09-27
环球时报(2019-08-09)2019-08-09
作文大王·低年级(2019年2期)2019-01-23
学苑创造·A版(2018年9期)2018-10-22
创新作文(1-2年级)(2018年7期)2018-01-22
当代工人(2016年16期)2016-11-04
数学大王·中高年级(2016年2期)2016-09-10
