
原花青素对小鼠血脂代谢紊乱与肠道菌群干预的影响
2020-07-23 04:23杨昌铭吴其国尹志婷周一鸣周小理
食品科学 2020年13期
杨昌铭,肖 瀛*,吴其国,尹志婷,周一鸣,周小理
(上海应用技术大学香料香精技术与工程学院,上海 201418)
随着我国经济水平的提高,居民饮食结构也发生了很大的变化,高脂膳食已成为目前大部分人的饮食习惯,由高脂膳食引起的血脂代谢紊乱成为普遍现象。脂质在维持人体内正常代谢平衡方面做出很大贡献。血脂代谢紊乱与心脑血管、糖尿病、高血压以及代谢综合征等关系密切[1],有效控制血脂代谢紊乱,可改善人们的健康水平,提升幸福指数。
肠道菌群是指寄居在人体肠道内并长期与人体相互依存的各种微生物,它数量庞大、种类繁多,与人体细胞总数比例约为1∶1[2],其对维持人体生理功能具有非常重要的作用,还与许多免疫性疾病、代谢性疾病等都有着紧密的联系[3-5]。高脂饮食会使肠道微生态发生改变[6],有研究报道高脂饮食在易肥胖鼠中造成肠杆菌增多,而在所有表型鼠中均可造成拟杆菌和梭菌目菌落的增多[7]。
原花青素是一种多酚类物质,它具有抗氧化性、降血脂和预防心血管疾病等活性[8]。原花青素的消化吸收主要发生在小肠部位,肠道菌群赖以生存的养料主要来自于未被小肠消化吸收的部分物质,原花青素及其代谢产物对肠道菌群有明显影响,原花青素与肠道菌群的相互作用可能是原花青素发挥生物活性作用的重要原因。二代测序Illumina MiSeq是目前应用最多的技术,具有低成本、时间短、通量高等优点[9]。本研究采用Illumina高通量测序为主要技术手段,以食源性原花青素分布广泛的二聚体B2为研究对象,探究原花青素B2干预高脂膳食模型小鼠血脂代谢紊乱及对肠道菌群结构的影响。
1 材料与方法
1.1 动物、材料与试剂
6 周龄SPF级(无特定病原体)雄性C57BL/6小鼠,体质量(16±1)g,购于上海灵畅生物科技公司,生产许可证号:SCXK(沪)2015-0014。
原花青素B2(纯度≥80%) 上海同田生物技术股份有限公司;肝素钠、无水硫酸钠、无水乙醚、氯化钠、浓硫酸(均为分析纯) 国药集团上海化学试剂有限公司;总胆固醇(total cholesterol,T-CHO)、甘油三酯(triglyceride,TG)、高密度脂蛋白胆固醇(highdensity lipoprotein cholesterol,HDL-C)、低密度脂蛋白胆固醇(low-density lipoprotein cholesterol,LDL-C)、脂蛋白脂酶、肝脂酶、总抗氧化能力(total antioxidant capacity,T-AOC)、总超氧化物歧化酶(total superoxide dismutase,T-SOD)、过氧化氢酶(catalase,CAT)、丙二醛(malondialdehyde,MDA)测定试剂盒 南京建成生物工程研究所;Quant-iT PicoGreen dsDNA试剂盒美国Invitrogen公司;E.Z.N.A.®soil 试剂盒 美国Bio-Tek公司;DNA提取、纯化和回收试剂盒 天根生化科技有限公司。
1.2 仪器与设备
AL204电子天平 梅特勒-托利多仪器(上海)有限公司;XW-80型混合器 海门其林贝尔仪器公司;K280R冷冻离心机 英国森特恩公司;TQ8040气相色谱-质谱仪 日本岛津公司;MiSeq PE300高通量测序平台 美国Illumina公司。
1.3 方法
1.3.1 实验分组与造模
小鼠以基础饲料饲喂适应1 周后,根据体质量随机分为对照组(A组:饲喂低脂低糖饲料,含有11%(质量分数,下同)脂肪、20%蛋白质和69%碳水化合物)、高脂组(B组:饲喂Paigen饲料,含有35%脂肪、20%蛋白质和45%碳水化合物)和原花青素B2组(C组:饲喂添加0.2%原花青素B2的Paigen饲料),每组7 只。小鼠自由饮食和饮水,相对湿度控制在(50±10)%,室温(22±2)℃,12 h明暗交替,饲养时间为12 周。
1.3.2 样品采集
在第12周,将待取样的小鼠置于干净的铺有无菌滤纸的笼子里,小鼠排便后立即收集粪便样本置于5 mL无菌EP管,-80 ℃保存,粪便采集完后,将小鼠禁食不禁水12 h,次日早晨,先将小鼠称质量,后用无水乙醚麻醉,进行眼球取血,将血放入预先准备好后的0.1%肝素钠生理盐水预处理过的无菌离心管中,然后放在冰浴中静止10 min左右,再以4 ℃、3 500 r/min离心15 min,取上清液置于离心管中,-80 ℃的冰箱保存待测。血取完颈椎脱臼处死后解剖,将小鼠的肝脏、腹脂、结肠、脑和心脏这5 个脏器称质量,其中肝和结肠于各小鼠固定位置称取0.1 g,置于含9 倍组织剪切质量的生理盐水匀浆器中冰浴条件下匀浆,制成质量分数10%的组织匀浆液,4 ℃、3 500 r/min离心10 min,取上清液,-80 ℃保存待测。
1.3.3 脏器指数、血脂指标、肝组织酶活力指标和抗氧化指标的测定
用试剂盒测定1.3.2节所取样本的T-CHO、TG、HDL-C、LDL-C、脂蛋白酶、肝脂酶、T-AOC、T-SOD、CAT、MDA水平。分别按式(1)、(2)计算脏器指数和动脉粥样硬化指数(atherosclerosis index,AI)。

1.3.4 粪便中DNA的提取与菌群多样性测序
小鼠粪便中DNA的提取按照试剂盒提供的说明书进行具体相关操作。按照V3~V4区域设计引物聚合酶链式反应扩增序列(引物名称为:338F、806R),然后再用Illumina MiSeq PE300平台进行测序(委托上海美吉生物医药科技有限公司进行测序)。
1.3.5 短链脂肪酸含量的测定
从收集的粪便中取0.1 g置于2 mL离心管中,加500 μL的饱和氯化钠溶液,在冰浴条件下用打碎机打碎均匀,再加20 μL 10% H2SO4溶液,混合器振荡混匀,到通风橱中加500 μL无水乙醚,混合器振荡混匀,14 000 r/min离心15 min,取上清液置于装有0.25 g无水硫酸钠的2 mL离心管中,再以同样的条件离心,取上清液置于气相瓶中待测。
气相色谱-质谱条件:采用Rtx®-WAX色谱柱(30 m×0.25 mm,0.25 μm);进样口温度:250 ℃,柱温:100 ℃,离子源温度:220 ℃,接口温度:250 ℃;升温程序:100 ℃到140 ℃升温速率为7.5 ℃/min,保持4 min,再以60 ℃/min升温至200 ℃;样品进样量:1 μL;载气:He;分流比:10∶1;流速:2 mL/min,溶剂延迟1.5 min;Q3 Scan方式采集1.5~9.5 min,质量范围m/z20.0~300.0。
1.4 数据统计与分析
实验获得的数据利用Excel软件对其进行预处理,结合SPSS Statistics 20软件的Duncan’s单因素方差分析对预处理的数据进行显著性分析(P<0.05)。对肠道菌群原始数据进行处理,得到优化的序列。按照操作分类单元(operational taxonomic unit,OTU)进行单元划分(97%相似度划分在一个单元),在OTU分类基础上进行物种丰度和多样性分析。
2 结果与分析
2.1 原花青素对高脂小鼠脏器指数的影响
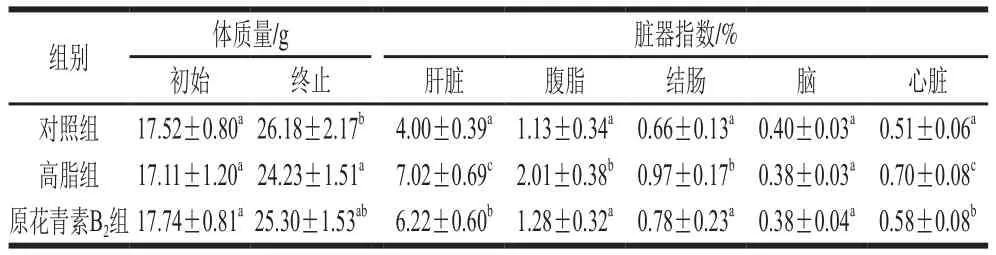
表1 原花青素B2对高脂小鼠脏器指数的影响Table 1 Effect of proanthocyanidin B2 on visceral organ indexes of HFD-fed mice
由表1可知,与对照组相比,高脂组小鼠的肝、腹脂、结肠和心脏均存在显著性差异(P<0.05),高脂膳食饲养的小鼠肝、腹脂和心脏指数均大于对照组和原花青素B2,且原花青素B2饮食干预后,高脂小鼠的肝、腹脂、结肠和心脏指数显著性降低(P<0.05)。
2.2 原花青素对小鼠血脂指标和肝组织酶活力指标的影响
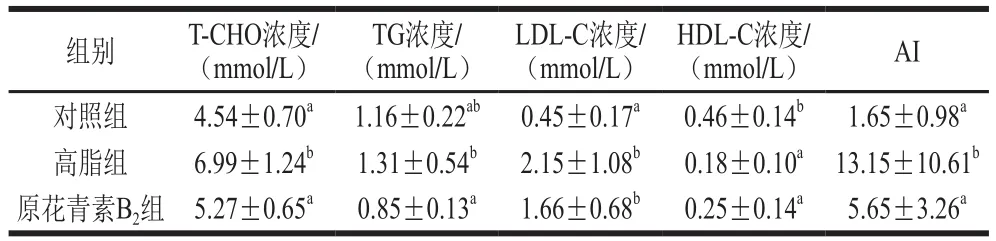
表2 原花青素B2对高脂小鼠血脂指标的影响Table 2 Effect of proanthocyanidin B2 on serum lipase indexes of HFD-fed mice
由表2可知,3 组小鼠中T-CHO浓度最高的是高脂组,且与对照组和原花青素B2组存在显著性差异(P<0.05)。原花青素B2组的TG浓度相对于高脂组显著降低(P<0.05),原花青素B2干预下HDL-C浓度增加但不显著。高脂组和对照组AI指数存在显著性差异(P<0.05),对照组和原花青素B2组没有显著性差异。表明原花青素B2可调节高脂膳食引起的血脂代谢异常。

表3 原花青素B2对高脂小鼠肝组织酶活力的影响Table 3 Effect of proanthocyanidin B2 on liver enzyme activities in HFD-fed mice
由表3可知,3 组总脂酶活力均存在显著性差异(P<0.05),3 组中高脂组总脂酶活力最低,原花青素B2组最高,脂蛋白酶活力中高脂组和原花青素B2组存在显著性差异(P<0.05),肝脂酶活力中对照组和高脂组有显著性差异(P<0.05),原花青素B2组和对照组无显著性区别。表明原花青素B2可增强小鼠肝酶活力,从而达到改善血脂异常的作用。
2.3 原花青素对小鼠抗氧化指标的影响
由表4可知,与对照组相比,高脂组小鼠肝的T-AOC、T-SOD、CAT和MDA色谱均存在显著差异(P<0.05),在原花青素B2干预后T-SOD和CAT活力显著增加,MDA含量显著降低(P<0.05),T-AOC有所增加但不显著。结肠的抗氧化活性中,高脂组的T-AOC和T-SOD活力与对照组相比差异显著(P<0.05),原花青素B2饮食干预后,T-AOC、T-SOD和CAT水平显著增加,MDA水平显著减少(P<0.05),表明原花青素B2具有抑制高脂小鼠氧化损伤的作用。
2.4 原花青素B2对小鼠肠道菌群多样性分析结果

图1 3 组小鼠粪便样品Venn图Fig. 1 Venn diagram of fecal microbiome in three mouse groups
如图1所示,样品在序列相似度97%的OTU数共有831 个,对照组有293 个,高脂组有265 个,原花青素B2组有273 个,对照组、高脂组和原花青素B2组共有201 个,另外对照组和高脂两组共有17 个,对照组和原花青素B2组共有23 个,高脂组共有和原花青素B2组共有37 个。
如图2A所示,横坐标是随机抽取的测序数据量,纵坐标是实际观测到的多样性指数(Shannon指数)。每条线代表一个样本,随着样本量的增加,线条在逐渐趋于平缓,可见Shannon曲线平台期。这表明测序的深度已经满足了检测样品中的所有物种。如图2B所示,运用统计学T检验的方法,Sobs指数中对照组和高脂组存在极显著性差异,表明原花青素B2饮食干预后,使得高脂模型小鼠的肠道菌群丰度增加。
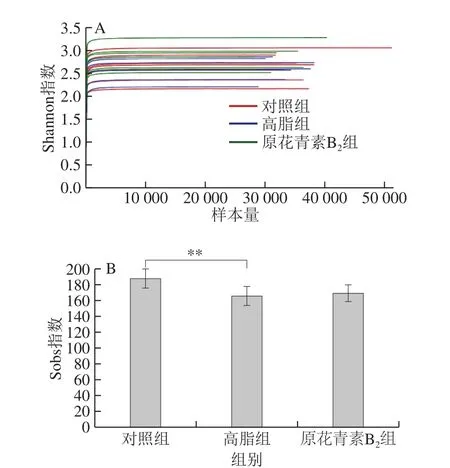
图2 3 组小鼠粪便样品稀释曲线和检验图Fig. 2 Rarefaction curves and test plots of fecal microbiome in three mouse groups
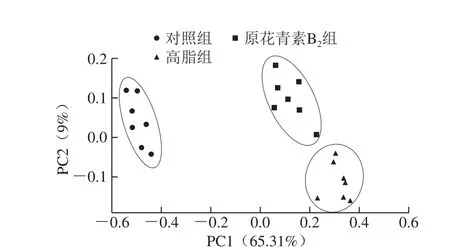
图3 3 组小鼠粪便样品PCoA分析图Fig. 3 Principal co-ordinates analysis of fecal microbiome in three mouse groups
主坐标分析(principal co-ordinates analysis,PCoA)结果如图3所示,横纵主坐标成分的贡献率分别是65.31%和9%,对照组、高脂组和原花青素B2组3 组样本点区别明显,对照组与原花青素B2组的样本点距离小于对照组和高脂组间的距离,说明原花青素B2组的菌群结构有向对照组的菌群结构模式调节的趋势。
2.5 原花青素对小鼠肠道菌群物种组成和差异性分析结果
基于门分类水平上原花青素对小鼠肠道菌群物种组成和差异性分析如图4所示,对照组中厚壁菌门(Firmicutes)、拟杆菌门(Bacteroidetes)、放线菌门(Actinobacteria)和变形菌门(Proteobacteria)在肠道菌群中相对丰度分别为51.39%、37.87%、9.42%和0.56%,与对照组比较,高脂组中厚壁菌门(Firmicutes)和变形菌门(Proteobacteria)是优势菌,并且相对丰度分别为58.21%和37.15%,放线菌门(Actinobacteria,0.79%)和拟杆菌门(Bacteroidetes,3.70%)的数量减少,原花青素B2组与高脂组相比较,其中厚壁菌门(Firmicutes,63.97%)和拟杆菌门(Bacteroidetes,14.69%)的数量增加,变形菌门(Proteobacteria,20.90%)和放线菌门(Actinobacteria,0.27%)的数量减少。拟杆菌门/厚壁菌门比值(B/F值)发生显著变化(P<0.05),其中高脂组B/F值显著降低,而原花青素组较高脂组B/F值显著升高(图5)。
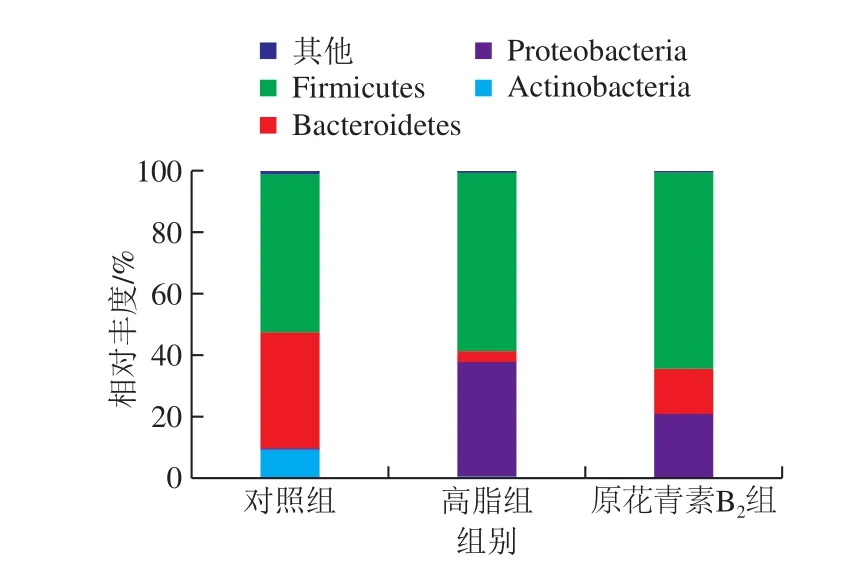
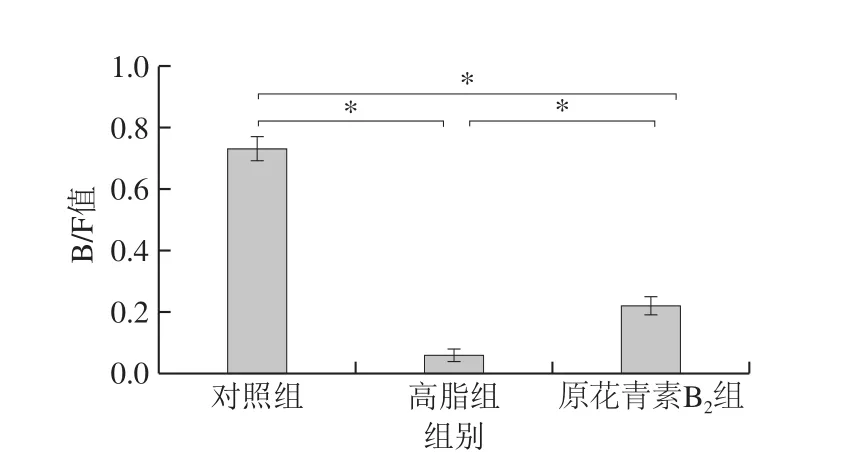
图5 拟杆菌门/厚壁菌门比值的变化Fig. 5 Changes in Bacteroidetes/Firmicutes ratio of fecal samples from three mouse groups at the genus level
由图6可知,基于属水平上,3 组小鼠肠道中埃希氏菌-志贺氏菌(Escherichia-Shigella)、未分类拟杆菌(Bacteroidales_S24-7_group)、肠杆菌属(Faecalibaculum)、Romboutsia、乳梭菌属(Lachnoclostridium)、乳杆菌属(Lactobacillus)、瘤胃球菌(Ruminococcaceae)、双歧杆菌属(Bifidobacterium)、拟杆菌(Parabacteroides)等13 种菌群有显著性差异。3 组相比较,对照组中未分类拟杆菌(Bacteroidales_S24-7_group)、肠杆菌属(Faecalibaculum)、乳杆菌属(Lactobacillus)和双歧杆菌属(Bifidobacterium)等菌属为优势菌属;高脂组中埃希氏菌-志贺氏菌(Escherichia-Shigella)、乳梭菌属(Lachnoclostridium)、瘤胃球菌(Ruminococcaceae)和无害芽孢梭菌(Clostridium_innocuum_group)等菌属占优势。与高脂组相比,原花青素B2组占优势的菌属有未分类拟杆菌(Bacteroidales_S24-7_group)、Romboutsia、裂梭菌群(Clostridium_fissicatena_group)、毛螺旋菌属(Lachnospiraceae)和拟杆菌(Parabacteroides)等菌属。
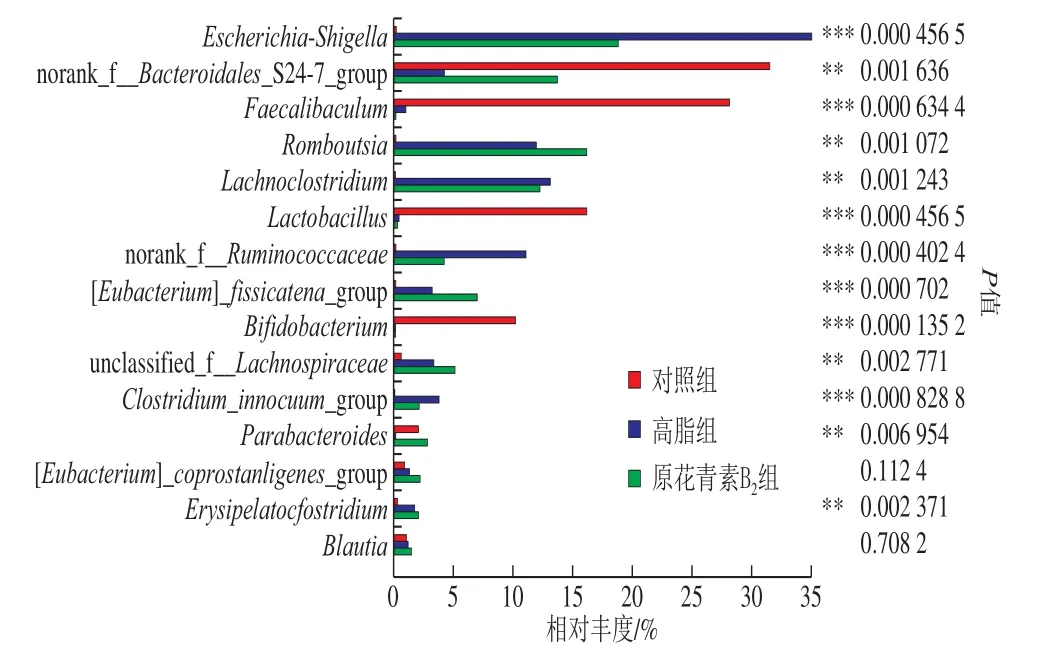
图6 基于属水平的3 组小鼠粪便样品物种差异分析Fig. 6 Analysis of fecal microbial species that differed among three mouse groups at the genus level
2.6 原花青素B2对小鼠粪便中短链脂肪酸的影响

表5 小鼠粪便中短链脂肪酸的含量Table 5 Contents of fecal short chain fatty acids in three mouse groups
原花青素B2对小鼠粪便中短链脂肪酸的影响结果见表5,高脂组较对照组在乙酸、丙酸和总酸含量显著增加(P<0.05),原花青素B2干预可显著增加丁酸的含量(P<0.05),这表明原花青素B2可调节产丁酸菌群的作用。
3 讨 论
本研究发现原花青素B2干预可显著降低高脂膳食小鼠模型的T-CHO、TG和AI指数,显著增加肝组织脂蛋白酶活力、肝脂酶活力以及机体抗氧化能力。长期高脂膳食会引起机体脂质代谢紊乱,从而导致血脂异常,如T-CHO、TG和LDL-C水平升高,HDL-C水平降低[10]。脂蛋白酶和肝脂酶是血浆脂蛋白代谢中的两个关键酶,由于具有高相似度的氨基酸序列,所以具有共同的作用机制[11],肝脂酶参与LDL、HDL和极低密度脂蛋白的产生[12]。Shi Fang等[13]通过高脂血症小鼠模型研究发现,LachnumYM226的降血脂作用部分是由于肝脏中脂蛋白酶和肝脂肪酶的调节实现的。Wang Bin等[14]研究壳聚糖对高脂饲料喂养的小鼠肝脏基因表达的影响,发现壳聚糖可降低高脂模型小鼠血脂指标和增加肝脂蛋白脂肪酶和肝脂肪酶活性,与本研究结果相似。
高脂膳食也可引起机体内自由基失衡,产生大量的自由基,导致氧化应激反应[15],在一些病理生理条件下,自由基产生和抗氧化能力之间的平衡被打破,导致氧化应激反应和组织损伤的增加[16],这些变化使机体产生动脉粥样硬化、心脑血管疾病和糖尿病等慢性疾病风险的几率增加[17]。原花青素B2具有较强的清除自由基,抗氧化和抑制脂质过氧化产物堆积的作用[18]。本研究发现高脂组小鼠血浆的T-CHO、TG、LDL-C水平显著上升,HDL-C水平下降明显,小鼠体内T-AOC和T-SOD活性均显著下降,而MDA水平上升,表明高脂组有着明显的脂质过氧化损伤的现象,这与Sour等[19]的研究结果一致。而高脂血症导致的血脂异常与结直肠息肉的发病息息相关[20],自由基及其产生的链式反应会导致组织损伤,引发组织炎症反应,甚至导致DNA损伤引发癌症[21],本研究发现高脂组小鼠结肠脏器指数上升,可能与结肠组织损失引发炎症反应有关,经过原花青素B2干预后结肠的脏器指数下降,表明原花青素B2具有抗氧化干预高脂膳食导致消化道氧化损伤的作用。
人体肠道菌群微生态失衡会引起许多人体疾病,例如代谢性疾病、消化性疾病和免疫性疾病[22]。有相关报道称肠道相关免疫系统对细菌成分的识别可能引起低水平炎症,该炎症是导致动脉粥样硬化的重要因素之一[23]。高脂膳食会致使小鼠体内B/F值降低[24-25],本研究的高脂膳食小鼠肠道B/F值显著降低,经过原花青素B2干预后比值增加,小鼠肠道菌群的结构得到了调节。厚壁菌门和拟杆菌门与机体的脂质代谢有着密切关系,它们会形成一种相互促进的共生关系,共同促进宿主对于多糖的代谢,促进能量的吸收和储存,而糖酵解所产生的磷酸二羟丙同酮还原后形成甘油,与丙酮酸氧化脱羧形成的乙酰辅酶A共同作用合成脂肪。一些致病菌如革兰氏阴性杆菌的增会导致肠道内产生大量的脂多糖(lipopolysaccharide,LPS),一方面,LPS会破坏肠道上皮细胞之间的紧密连接,导致肠道通透性的增加,随后LPS通过上皮细胞间的间隙进入体内血液循环,使机体产生炎症反应,从而对肝脏的功能产生显著影响,导致肝脏的脂代谢能力下降,引起脂肪积累在体内[26];另一方面,LPS和革兰氏阴性菌的鞭毛会使肠道上皮细胞对脂肪的吸收增加,血液内的游离脂肪酸含量增加,机体内的脂肪不断积累,最终导致肥胖等脂代谢紊乱[27]。
Cheng Mei等[28]建立小白鼠高脂血症动物模型,给小鼠服用乳酸制剂和他汀药物(降血脂药),比较发现它们都显著降低T-CHO、TG、HDL-C的含量,达到降血脂的功能,而灌胃乳酸菌的小鼠肠道中乳酸菌的数量显著增加,大肠杆菌和肠球杆菌的数量显著降低,表明调节肠道菌群与降血脂有显著的关联性。本研究发现原花青素B2干预高脂模型小鼠的肠道中埃希氏菌-志贺氏菌(Escherichia-Shigella)显著性降低,未分类拟杆菌(Bacteroidales_S24-7_group)显著性增加。埃希氏菌-志贺氏菌(Escherichia-Shigella)为革兰氏阴性菌,赵梅等[29]从一名重度肥胖患者肠道内分离出19 种不同的ERIC类型的分离物,进行16S rRNA基因分子鉴定,发现19 种代表菌株的16S rRNA基因和埃希氏菌属和志贺氏菌属相似度高达99%。表明高脂膳食可通过增加埃希氏菌-志贺氏菌的丰度从而改变肠道菌群结构。未分类拟杆菌(Bacteroidales_S24-7_group)属于丁酸盐产生菌,有研究显示高脂膳食会引起机体内肠道菌群未分类拟杆菌(Bacteroidales_S24-7_group)减少致使丁酸盐减少,丁酸盐的减少会破坏肠菌群生存环境的pH值,弱酸环境对于双歧杆菌(Bifidobacterium)等益生菌的生长比较有利[30-31],本研究也发现原花青素B2干预高脂膳食小鼠肠道中双歧杆菌相对丰度也有增加的趋势。Hua等[32]研究证明,益生菌能够刺激机体免疫功能,上调抗炎细胞因子,抑制机体炎症反应,其代谢产物还能促进肠上皮细胞的修复和再生,从而有效减少LPS的吸收。
短链脂肪酸是由肠道菌群发酵部分未消化的碳水化合物、蛋白质与脂肪等营养素在结肠中产生的,它们能够促进机体内难消化产物的吸收和结肠上皮细胞增殖与黏膜生长,短链脂肪酸的增加还具有调节改善结肠的功能[33-35]。本研究发现高脂膳食可导致肠道乙酸含量增多,这可能是过多摄入的脂肪等未被消化的营养素在结肠经乙酰辅酶A或Wood-Ljungdahl途径产生,研究发现体内乙酸的增加可导致胰岛素抵抗的作用,从而加剧脂代谢异常[36]。本研究结果发现高脂膳食小鼠经原花青素B2干预,肠道中丁酸含量显著增加。研究表明,丁酸的代谢至少占结肠细胞能量需求的70%,丁酸可通过增加磷脂转运蛋白mRNA的水平及磷脂转运蛋白的分泌,促进胆固醇的转运[37],激活蛋白PGC-1α增强线粒体功能,从而加速脂肪氧化[38],调节脂代谢异常[39]。因此,原花青素B2干预可调节肠道中的拟杆菌、双歧杆菌、消化链球菌和梭菌属等丰度的变化,从而改善小鼠血脂代谢异常。
本研究结果表明原花青素B2可以降低高脂小鼠血脂水平、抗氧化活性和增强肝酶活性,调节肠道菌群丰度和结构,促进短链脂肪酸的产生,从而可能改善血脂代谢紊乱。
猜你喜欢
中老年保健(2022年2期)2022-08-24
中国饲料(2022年5期)2022-04-26
科学(2020年4期)2020-11-26
中华养生保健(2020年5期)2020-11-16
天然产物研究与开发(2018年11期)2018-11-30
中成药(2017年5期)2017-06-13
中国运动医学杂志(2016年3期)2016-07-10
中成药(2016年8期)2016-05-17
中国康复理论与实践(2015年10期)2015-12-24
医学研究杂志(2015年12期)2015-06-10
