
基于SSR标记的朝天椒种质遗传多样性分析
2020-07-22 12:32任佳佳安朝龙吴培云谢龙安
河南农业科学 2020年7期
张 曼,任佳佳,王 钦,安朝龙,吴培云,谢龙安,李 伟
(1.贵州大学 农学院,贵州 贵阳 550025; 2.毕节职业技术学院,贵州 毕节 551700)
辣椒(CapsicumannuumL.)别名番椒、海椒、秦椒,起源于墨西哥、中南美洲的热带地区[1]。辣椒不仅是人们喜爱的蔬菜,还可作为新型绿色药品加以利用,栽培规模日益扩大,是我国仅次于大白菜的第二大蔬菜作物,辣椒营养丰富,含有大量的辣椒素、辣椒红素、胡萝卜素、碳水化合物、矿物质等,尤其是维生素C(Vc)含量更是高居各类蔬菜的榜首[2-3]。朝天椒(CapsicumfrutescensL.)是辣椒的一个变种,为茄科(Solanaceae)辣椒属(CapsicumL.)植物,椒果小、簇生且朝天,辣度高、易干制,既可鲜食、泡制,又可作为干辣椒利用,与羊角椒、线椒构成我国三大干椒品种系列。朝天椒的栽培面积逐年上升,已跃居干椒栽培面积首位[4-5]。
鉴定朝天椒试验材料的遗传多样性,全面了解这些品种的遗传基础和亲缘关系,对发掘利用优异的育种材料,拓宽遗传基础,加快朝天椒育种改良进程具有重要意义。对于种质材料遗传多样性的研究,表型性状极易受环境因素影响而变化,仅依靠表型性状难以准确反映不同材料间的遗传差异和亲缘关系,基于分子水平的亲缘关系较基于系谱的亲缘关系具有更高的可靠性[6]。随着分子生物学的发展,各种分子标记已经广泛应用于辣椒遗传多样性研究中[7-13]。SSR(简单串联重复序列)具有高分辨率、高稳定性、重复性好、共显性遗传和操作简单等特点[14],已在辣椒的遗传多样性分析中得到了广泛应用[15-20]。但是,采用SSR分子标记进行朝天椒遗传多样性的分析较少,蒋向辉等[21]和贺洁等[22]均分析了朝天椒的遗传多样性,但分析材料较少;曾绍贵等[23]采用形态学标记和SRAP(序列相关扩增多态性)分子标记相结合的方法对100份朝天椒品种进行遗传多样性分析;詹永发等[24]采用形态学标记分析了156份朝天椒遗传多样性。目前,尚未见关于采用SSR分子标记技术分析大批量的朝天椒遗传多样性研究。因此,以西南地区朝天椒地方种质为研究材料,从分子水平进行遗传多样性分析,明确它们之间的遗传关系,为朝天椒后续育种亲本选择、种质资源有效利用及种质创新提供依据。
1 材料和方法
1.1 试验材料
试验材料主要为西南地区的朝天椒地方种质,共计112份(表1),主要来源于贵州、四川、云南、重庆。其中,贵州省内收集的朝天椒地方种95份,其他包括四川省4份,云南省5份,重庆市8份。

表1 朝天椒参试材料Tab.1 Capsicum frutescens varieties

续表1 朝天椒参试材料Tab.1(Continued) Capsicum frutescens varieties
1.2 DNA提取及引物选择
每份材料随机选取20个单株中的幼嫩叶片,混匀后用新型快速植物基因组DNA提取试剂盒(无锡百泰克生物技术有限公司)提取基因组DNA,所提取的DNA使用核酸测定仪检测DNA质量及浓度,加TE缓冲液稀释到50 ng/μL,用于PCR反应。利用中国农业行业标准(NY/T 2475—2013)和贵州省地方标准(DB/T 953—2014)推荐的SSR核心引物,在前人研究[25-28]的基础上挑选出124对引物,随机选用24份朝天椒种质资源进行多态性引物筛选,最终筛选出20对条带清晰、多态性稳定的SSR引物用于后续的遗传多样性分析(表2)。引物由上海生工生物工程股份有限公司合成。

表2 SSR引物序列Tab.2 Sequence of SSR primers
1.3 PCR反应体系与扩增程序
PCR扩增体系为20 μL,包括50 ng/μL模板DNA 1 μL,上下游引物各0.5 μL,ddH2O 8 μL,2×TaqMaster mix 10 μL。PCR扩增反应程序:94 ℃预变性4 min;94 ℃变性45 s,55 ℃(根据引物退火温度改变)退火45 s,72 ℃延伸45 s,35个循环;72 ℃延伸10 min,4 ℃保存待测。
1.4 电泳与银染显色
使用DYY-6C型电泳仪,取2 μL扩增产物进行8%非变性聚丙烯酰胺凝胶电泳检测,电泳液为1×TBE,恒定电压150 V,电泳时间2~4 h。电泳结束后,将胶浸入固定液(10%乙醇和5%冰醋酸)中在摇床上固定10 min,用蒸馏水洗2遍;然后用 AgNO3染色10 min,再用蒸馏水漂洗2次;最后用显色液(200 mL显色液包含3 g NaOH、1 mL甲醛)显色至条带清晰,用蒸馏水洗脱3次停显。最后置于白炽灯箱上照相、记录。
1.5 数据统计分析
将得到的扩增图片以清晰稳定的扩增条带进行SSR标记数据统计,根据SSR标记统计的A、B、C等数据结果,用POPGENE 32软件对不同类群的观测等位基因数(Na)、有效等位基因数(Ne)、观测杂合度(Ho)、期望杂合度(He)和Shannon’s信息指数(I)等进行遗传多样性数据计算和对比分析,根据SMITH等[29]的方法计算标记位点的多态性信息量指数(PIC),根据SSR标记统计的0、1数据结果,利用NTSYS-pc 2.1软件计算遗传相似系数(GS),采用类平均法进行UPGMA聚类分析。
2 结果与分析
2.1 SSR多态性引物的扩增结果
利用随机选择的24份朝天椒材料对124对SSR引物进行引物扩增多态性筛选,筛选出20对扩增效果好、条带差异明显的SSR引物,用于112份朝天椒材料的遗传多样性分析,图1为引物19的部分扩增结果。利用筛选出的20对引物共检测出65个等位变异(表3),每对引物的等位变异为2~4个,平均每对引物3.25个等位变异, 20对引物的有效等位基因数为1.311 9~2.834 5,平均为1.829 4。引物2、5、8、10、11、12、10扩增出的等位变异最多,均为4个,其有效等位基因数分别为1.736 1、2.004 0、2.834 5、1.743 3、1.724 1、1.850 8、1.621 8。各引物的PIC值为0.324 1~0.696 0,平均为0.544 0,20对引物中有16对引物PIC值均大于0.5,其中8号引物PIC值最大,为0.696 0。
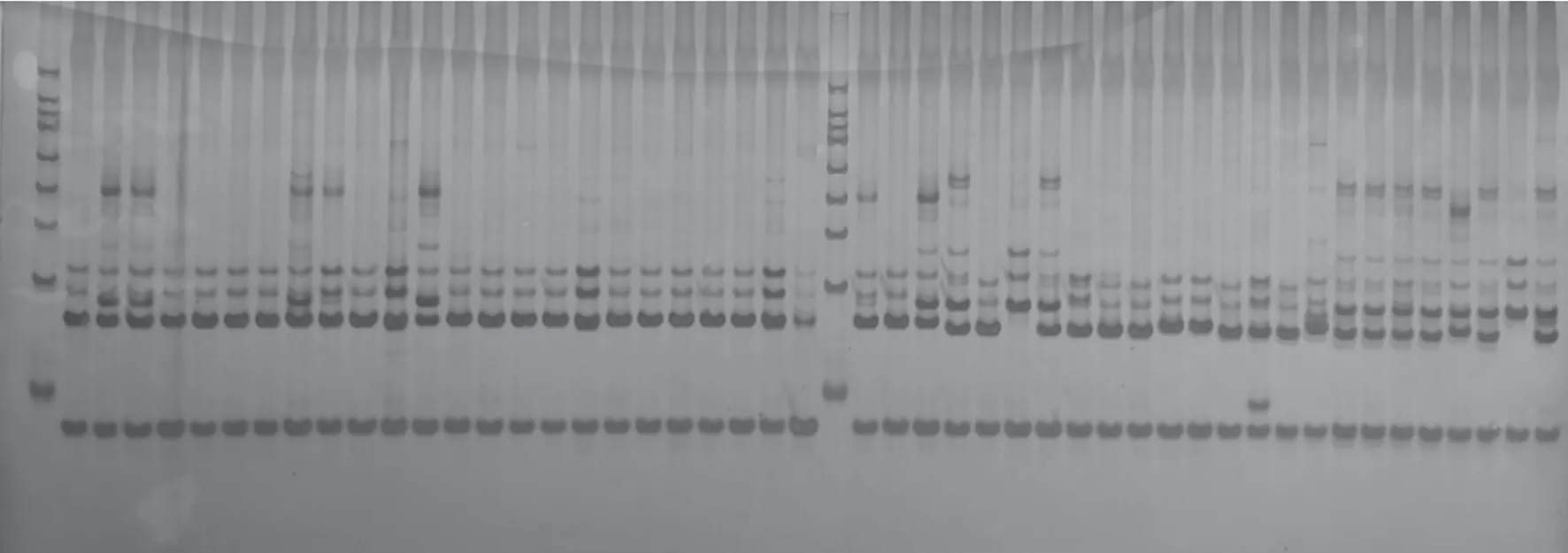
图1 19号引物对部分朝天椒材料SSR扩增结果Fig.1 SSR amplification results of some Capsicum frutescens materials by primer 19
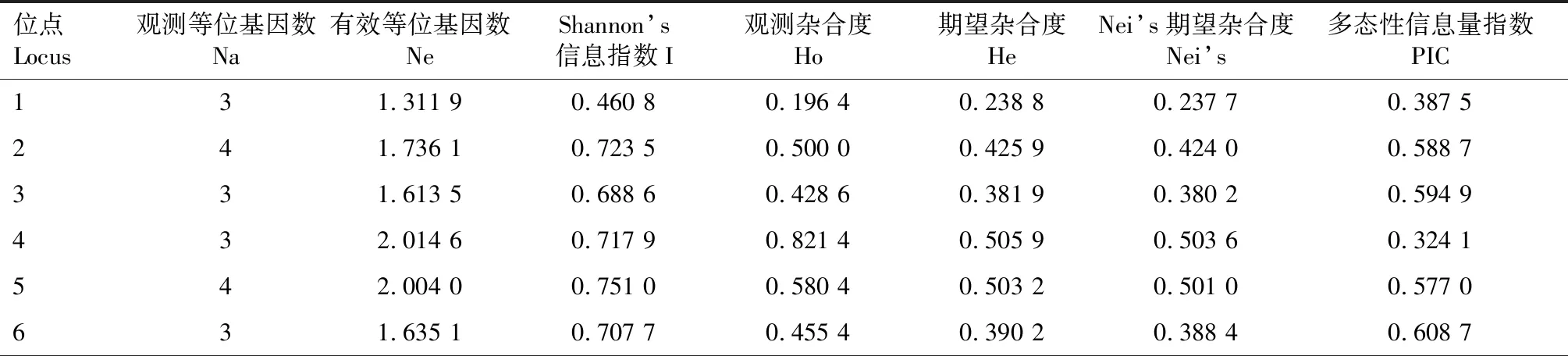
表3 20对SSR引物的遗传参数Tab.3 Genetic parameters from the 20 polymorphic SSR markers
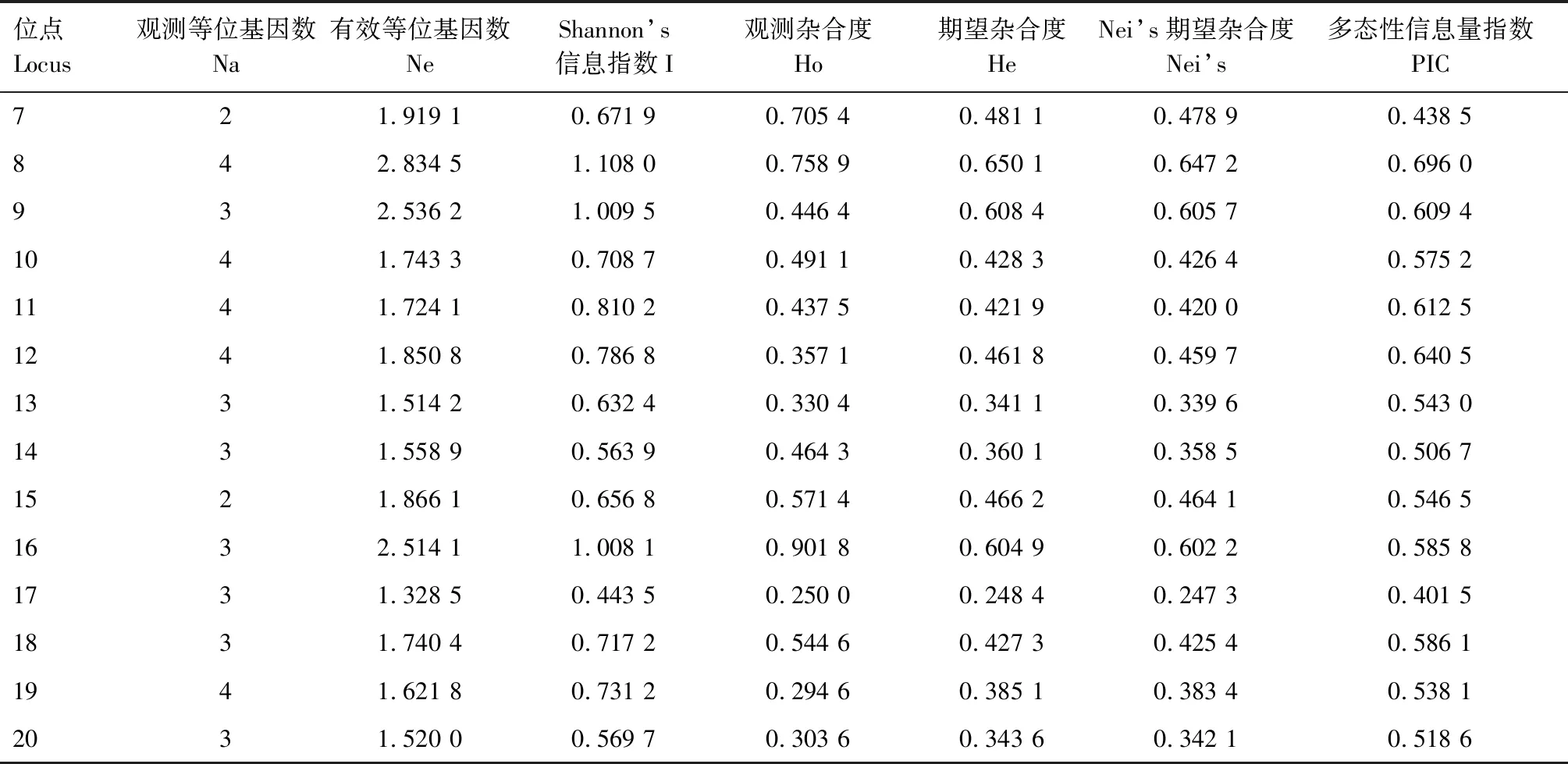
续表3 20对SSR引物测定的遗传参数Tab.3(Continued) Genetic parameters from the 20 polymorphic SSR markers
2.2 SSR引物的遗传多样性分析
基于SSR数据居群间的Shannon’s信息指数(I)为0.443 5~1.108 0,平均为0.723 4,Shannon’s信息指数最大的位点是引物8,最小的为引物17;观测杂合度(Ho)为0.196 4(引物1)~0.901 8(引物16),平均值为0.492 0;期望杂合度(He)为0.238 8(引物1)~0.650 1(引物8),平均值为0.433 7;Nei’s期望杂合度为0.237 7(引物1)~0.647 2(引物8)。以上结果表明,所筛选出的20对SSR引物多态性较高,能较好地反映朝天椒不同材料间的遗传多样性信息。
2.3 朝天椒种质间的遗传相似性分析
根据20对SSR引物扩增产生的65个等位变异,计算了112个朝天椒种质居群间的GS值。结果表明,供试材料居群间GS值为0.569 2~0.953 8,平均值为0.773 3,说明朝天椒种质间的遗传相似性相对较高,材料间的亲缘关系非常接近。7号材料和8号材料、22号材料和23号材料GS值最大(7号和8号材料均来自贵州息烽县,22号和23号材料均来自红花岗区),表明这几个朝天椒居群间的遗传距离最近,遗传相似度最大,说明亲缘关系越近,遗传相似性越高;4号材料和86号材料GS值最小(4号材料来自息烽县,86号材料来自台江县),表明这2个材料间遗传相似性很小,它们之间的亲缘关系较远。
2.4 朝天椒种质的SSR聚类分析
以SSR分子标记在112个材料间多态性数据计算GS值,采用类平均法(UPGMA)聚类分析(图2),结果表明,在阈值0.767处将供试材料分为4类,第1类包含55份材料,55份材料中贵州省内的共47份,分别来自开阳县(2份)、息烽县(7份)、桐梓县(2份)、遵义县(4份)、红花岗区(1份)、绥阳县(1份)、道真县(1份)、务川县(1份)、平坝县(1份)、罗甸县(3份)、六枝特区(1份)、玉屏县(4份)、沿河县(1份)、思南县(1份)、大方县(4份)、黎平县(2份)、台江县(9份)、开阳县(1份)、福泉市(1份),四川省2份,重庆市6份,其中来自玉屏县(57号和58号)、福泉市(95号)和四川省(98号)的材料与其他51份材料的遗传距离较远;第2类只包含1份材料,来自兴义市,说明该材料与其他材料间遗传差异较大,亲缘关系相对较远;第3类包含14份材料,来自贵州省的有11份,分别来自遵义县(1份)、沿河县(5份)、思南县(2份)、台江县(3份),四川省1份,云南省1份,重庆市1份;第4类包含42份,来自贵州省的有36份,分别来自桐梓县(4份)、遵义县(1份)、红花岗区(2份)、绥阳县(2份)、道真县(1份)、务川县(2份)、三都县(4份)、贵定县(3份)、盘县(1份)、六枝特区(5份)、兴仁县(4份)、沿河县(2份)、大方县(4份)、黎平县(1份),四川省1份,云南省4份,重庆市1份,其中来自六枝特区(4份)和四川省(97号)的材料与其他37份材料的遗传距离较远。112份朝天椒材料中,整体分布与地理分布有一定关联,但也存在区域间和省内外间的互相渗透,每个类别均聚集了不同地理来源的材料。

图2 112份朝天椒种质资源聚类分析
3 结论与讨论
本研究从124对引物中筛选出20对SSR标记,对西南地区的朝天椒地方种质,共计112份进行遗传多样性分析,平均每个标记扩增出3.25个等位变异。陈文超等[30]利用33个EST-SSR标记分析了31份辣椒种质的遗传多样性,平均每个位点扩增出2.76个等位基因。李艳等[31]利用17对SSR引物对169份辣椒材料进行遗传多样性分析,平均每对引物扩增出2.71个位点。贾豪等[19]利用41对SSR引物分析了24份辣椒种质资源的遗传多样性,平均每对引物扩增出5.15个位点。傅鸿妃等[20]利用31对SSR引物对52份辣椒种质材料的遗传多样性进行分析,平均检测到6.613个等位位点。本研究中SSR位点检测到的平均等位基因数量高于陈文超等[30]和李艳等[31]的结果,却低于贾豪等[19]和傅鸿妃等[20]的结果。表明本研究所筛选出的SSR引物可以反映出种质之间的遗传多样性,只是引物多态性信息不够丰富,其原因可能是由于朝天椒为辣椒变种,为辣椒种下的一个亚种,而所选引物均是研究辣椒遗传多样性的,还有原因可能是所选112份材料绝大多数来自于贵州本地,其亲缘关系较近。按照BOTSTEIN等[32]提出的衡量指标,当PIC值>0.5时,该位点为高度多态位点,本试验中的平均PIC值为0.544 0,20对引物中有16对引物PIC值均>0.5,表明这些位点较好地反映了朝天椒材料的遗传多样性,适合应用于朝天椒材料的遗传多样性检测。
SSR聚类分析表明,供试材料中的平均遗传相似系数是0.773 3,说明朝天椒品种间的遗传相似性相对较高,材料间的亲缘关系比较接近,说明大部分材料亲缘关系较近,需要努力拓宽品种选育的遗传基础,112份朝天椒材料类群划分结果与品种的地理来源有一定关联,但群体遗传分化不明显,群体间的遗传关系较近,且不同群体间材料存在着互相渗透,可能原因是所选择的群体范围比较窄,或者选择群体不够大。本研究利用20对SSR标记分析了112份朝天椒的遗传多样性,将112份材料分成了4类,为朝天椒地方种资源间亲缘关系的揭示、资源保存和新品种选育提供了参考依据。
猜你喜欢
作物学报(2022年2期)2022-11-06
社会科学战线(2022年7期)2022-08-26
世界科学技术-中医药现代化(2022年3期)2022-08-22
昆明医科大学学报(2022年3期)2022-04-19
昆明医科大学学报(2022年2期)2022-03-29
农技服务(2021年11期)2021-12-17
小作家报·教研博览(2021年33期)2021-10-16
昆明医科大学学报(2021年2期)2021-03-29
北方文学(2019年21期)2019-08-18
小说月刊(2014年1期)2014-04-23
