
长期增温对荒漠草原短花针茅群落组成和土壤养分及酶活性特征的影响
2020-07-21 02:17:44马丹丹王瑾瑜
甘肃农业大学学报 2020年3期
马丹丹,王瑾瑜
(1.河南林业职业学院,河南 洛阳 471002;2.河南城建学院建筑与城市规划学院,河南 平顶山 450007)
全球气候变暖已经成为不容置疑的事实[1-3],到本世纪末(2100年),全球平均气温将升高1.8~4.0 ℃,以全球气候变暖为主要特征的全球气候变化已引起人们的广泛关注[4-5].气候变化决定着地球上物种的分布以及植被类型,而温度是气候条件中最重要的因素,温度的升高对植物群落组成及物种多样性产生重要的影响.增温直接改变植物的光合能力,影响植物在光合作用中各种酶的活性,进而影响植物的生长速率,改变植物的物候和生长期[6-7].同时,温度的增加会改变土壤含水量和土壤呼吸,导致植被的生长、生物量的生产和分配、植物群落演替方向和速度都随之发生改变.
土壤酶系统在陆地生态系统地下生态过程中扮演着十分重要的角色[8].它是土壤生态系统中的重要组成部分及土壤生物化学过程的积极参与者,也是土壤生态系统物质和能量流动过程中最为活跃的生物活性物质,保持了土壤生物化学的相对稳衡状态[9].温度是控制土壤酶活性的一个重要环境因子,气候变化可能对陆地生态系统土壤酶活性产生深刻影响.有研究表明,土壤温度升高能增加土壤酶活性[10].温度升高可能通过影响土壤微生物群落的组成结构、微生物多样性、微生物生物量、微生物呼吸作用、土壤有机物质的矿化率、土壤的水热条件、有机物质分解等因素间接影响土壤酶的活性[11-12].不同生态系统及不同种类的土壤酶对温度的敏感性存在差异.如土壤蛋白酶更多受土壤水分影响而非温度,而脲酶和酸性磷酸酶对温度很敏感.因此,深入研究温度条件变化对土壤酶活性的影响以及土壤酶的反馈机制,有利于我们更加深入地了解全球变化背景下营养元素的循环格局及其反馈机理等[13].
荒漠草原以短花针茅群落为优势物种,其群落多样性和稳定性能够指示荒漠生态系统的稳定性.荒漠草原年均降雨量少,季节温差和日温差十分显著,植物种类相对贫乏,土壤酶活性较低,严酷的生态环境使得荒漠草原的生态系统十分脆弱.具有发生退化的潜质,一旦遭到破坏,非常难以恢复[14].前人关于短花针茅群落物种丰富度、多样性、稳定性土壤养分、酶活性等进行了大量的研究[15-16],然而短花针茅群落结构和土壤性质对气候变化(增温)响应的研究尚不多见.因此,研究模拟增温下荒漠草原物种产生的反应或适应性变化有助于理解这些物种响应温度升高等环境变化的内部机制,预测其对生态系统群落功能、生物多样性等的影响,以应对未来环境变化,从而为荒漠草原的利用和生态系统的可持续管理提供依据.
1 材料与方法
1.1 研究区概况
试验区位于内蒙古自治区乌兰察布市四子王旗王府一队的内蒙古农牧科学院实验基地内(N 41°47′17″,E 111°53′46″),海拔高度1 456 m,气候属于中温带大陆性气候,多年平均气温3.4 ℃,月平均温度最高为6~8个月,年均气温分别为21.5、24.0、23.5 ℃,≥10 ℃的年积温为2 200~2 500 ℃.多年的平均降雨量248 mm,年均蒸发量2947 mm,降水量主要集中在6~9月,占全年降水总量的70%以上.
实验区主要由旱生型的禾草组成,属于短花针茅建群的荒漠草原地带性植被,草地类型为短花针茅(Stipabreviflora)+冷蒿(Artemisia frigida)+无芒隐子草(Cleistogenessongorica),植被高度低矮,平均高度为8 cm,且植被较稀疏,盖度为17%~20%,种类组成贫乏,植物群落主要由20 多种植物物种组成.建群种为短花针茅,优势种为无芒隐子草和冷蒿.主要伴生种有阿氏旋花、栉叶蒿(Neopallasiapectinata)、木地肤(Kochiaprostrata)、细叶葱(Alliumtenuissimum)、蒙古葱(Alliummongolicum)、狭叶锦鸡儿(Caraganastenophylla)、猪毛菜(Salsolacollina)和小叶锦鸡儿(Caraganamicrophylla)等.
1.2 试验设计
采用国际冻原计划(ITEX)模拟增温的方法,建造增温室,试验地周围用网围栏封育,禁止放牧[17].每个增温室高2 m,基面积为4 m2、顶面积为4 m2,通过不封顶和封顶实现不同的增温梯度控制,同时每个增温室旁边设置相同面积的对照,每个处理3次重复.为了满足与自然控制和开顶式增温室相似的自然条件,封顶式增温室通过定期的开门来保持与外界通风,并不间断的洒水来保持与外界一致的降水条件.增温区内各安装一个远红外线辐射器进行加热,该装置是通过悬挂在样地中央上方、可以散发远红外线辐射的灯管来实现模拟气候变暖,是目前国际上公认的增温措施.远红外线辐射器由美国伯利恒Kalglo电子公司生产,型号MSR-2420,规格165 cm×15 cm,辐射器距地面2.25 m,安放于3 m×4 m小区的中间部位,灯的最大功率是2 000 W,使用时调至8档.同时,在每个处理小区内随机安装一组测定土壤温度的探头,同时采用Omega 公司生产的型号为HH-25TCSensors 手持温度计与之连接测定10、20、30、40 cm不同土层的土壤温度.同时采用ONSET 电脑公司生产的其型号为S-SMA-M003的土壤湿度探头对0~40 cm土层中每10 cm土壤湿度进行测定.测定数据采用HOBO软件进行记录数据,每隔30 min采集1次.
1.3 群落结构调查
2013年7月,完成增温室的安装,开始对空气和土壤增温(增温2 ℃).在模拟增温1 a后,于2014年8月中旬,分别在对照样地和每个OTC内随机选取样方9 个(25 cm×25 cm),对各群落中各物种的高度、密度、盖度和频度进行统计调查,然后借助样方框(由25个面积为5 cm×5 cm 的方格网组成),目测群落的总盖度和分盖度;连续6 a观测调查.
同年9月份齐地面剪草,收割地上生物量,将活体、立枯和凋落物分开保存,地下生物量用直径5 cm的土钻分层取样,每个样地重复5 次,并立即用网孔为1 mm的滤网洗尽土壤.将地上和地下样品带回实验室,在65℃下烘干至恒质量并称质量.
Shannon-Wiener多样性指数(H)、McIntosh均匀度指数(JP)、Margalef丰富度指数(S)、Simpson优势度指数(D)计算[13]:
H=-∑(PilnPi)
JP=H/lnS
S=(N-1)/lnN
D=1-∑(Pi)2
重要值Pi=100×(相对盖度+相对高度+相对密度)/3[18]
式中,N为植物总数;Pi为第i种的植物个体数量(ni)在总个体数量(N)中的比例.
1.4 土壤酶活性的测定
有机碳采用重铬酸钾外加热法,全氮和有效氮采用半微量凯氏定氮法,全磷和有效磷采用钼锑抗比色法,全钾和有效钾采用火焰分光光度计法[19].
过氧化氢酶活性的测定采用高锰酸钾滴定法,活性单位以1 g土样消耗的0.1 mol/L KMnO4溶液的毫升数表示;蔗糖酶活性的测定采用3,5-二硝基水杨酸比色法,活性单位以1 g土样在37 ℃条件下,经过24 h反应后水解生成的葡萄糖的毫克数表示;蛋白酶用偶氮酪素比色法,以24 h后1 g土壤中氨基氮的毫克数表示;脲酶活性的测定采用比色法,活性单位以1 g土样在37 ℃ 条件下,经过24 h反应后水解生成的氨基氮的毫克数表示;碱性磷酸酶活性测定采用磷酸苯二钠比色法测定,以24 h后100 g土壤中P2O5的毫克数表示.
1.5 数据分析
采用Excel 2003.0和SPSS 18.0数据统计和单因素方差分析(One-way ANOVA),所有值采用2014~2019年的平均数据,LSD多重比较法检验各处理间差异显著.
2 结果与分析
2.1 温度升高对土壤温度和湿度的影响
由表1可知,在整个生长季,增温样地0~10、10~20、20~30、30~40 cm土壤平均温度均高于对照样地,从3月到11月,OTC和CK样地土壤平均
温度呈一致的变化趋势,均呈先增加后降低趋势,在7月达到最大.OTC和CK样地土壤平均温度随着土壤深度的增加而逐渐降低趋势,在表层OTC和CK土壤平均温度相差较大,在30~40 cm,OTC和CK土壤平均温度基本一致,说明荒漠草原增温试验能够改变土壤温度,主要集中在表层土壤.在整个生长季,增温样地0~10、10~20、20~30 cm土壤平均含水量均低于对照样地,从3月到11月,OTC和CK样地土壤平均含水量呈一致的变化趋势,均呈先增加后降低趋势,在7月达到最大.OTC和CK样地土壤平均含水量随着土壤深度的增加而逐渐降低趋势,在表层OTC和CK土壤平均含水量相差较大,在30~40 cm,OTC和CK土壤平均含水量基本一致,说明荒漠草原增温试验能够改变土壤含水量,主要集中在表层土壤.总的来说,荒漠草原增温试验改变了土壤不同层次的温度与土壤含水量.
2.2 温度升高对植物群落组成和特征值的影响
经过增温处理后,增温样地和对照样地群落物种组成基本相同,增温样地和对照样地共有10个物种(表2),说明增温作用没有改变试验样地内植物群落组成.但是增温改变了各物种的重要值,在增温样地内物种重要值均高于对照样地.由表还可以看出,温度升高使群落的高度整体上有所增加,盖度、频度和密度也呈增加趋势.增温的处理改变了物种的密度、高度、盖度和频度,进而改变了物种的重要值.

表1 温度升高对土壤温度和湿度的影响
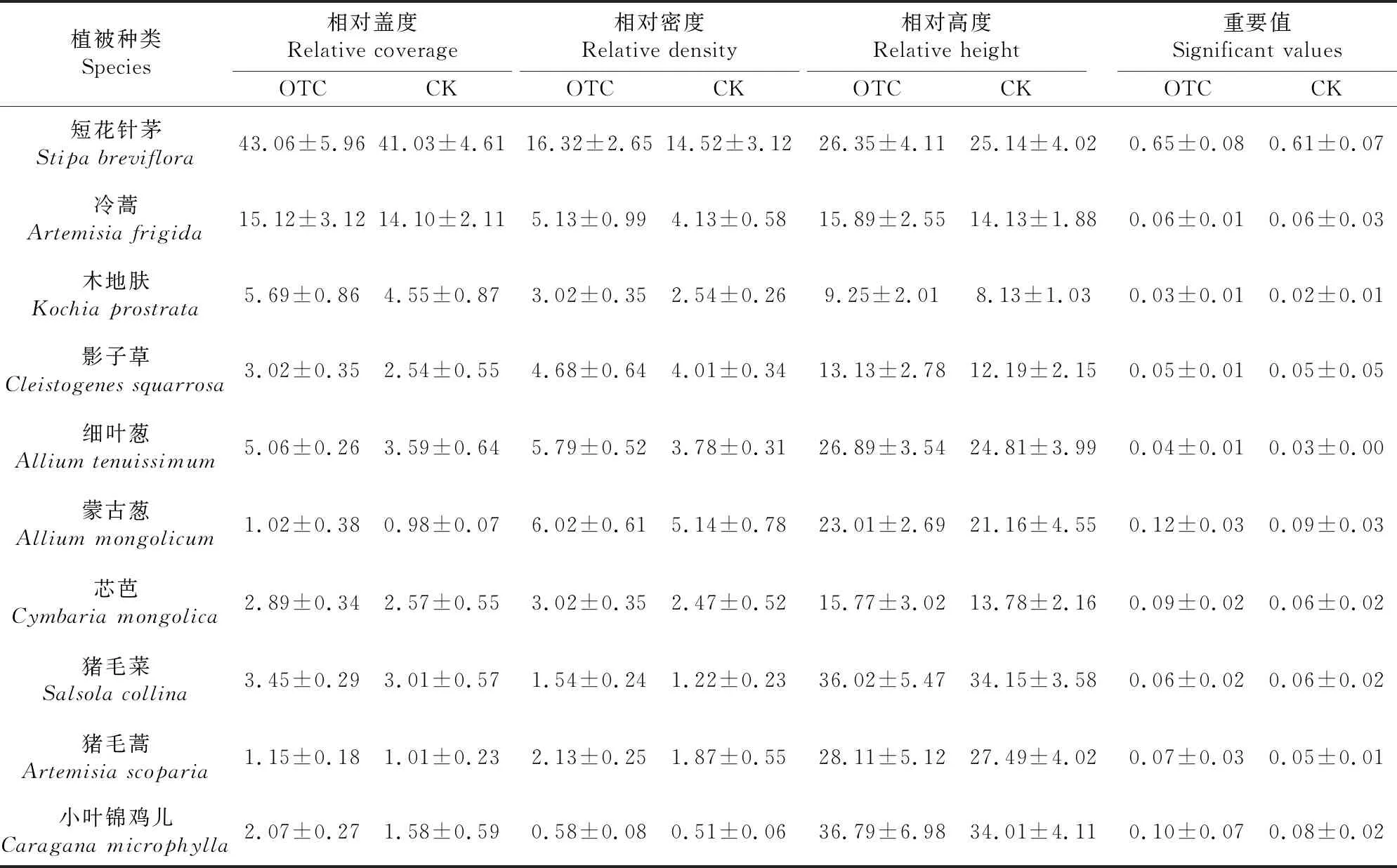
表2 温度升高对植物群落组成和特征值的影响
2.3 温度升高对植物群落生物量分配的影响
增温后,增温样地内的地上生物量和凋落物均显著高于对照样地(P<0.05),与对照样地相比,地上生物量显著增加了8.94%,凋落物显著增加了7.03%(图1).从0~40 cm地下生物量分析,增温样地地下生物量大于对照样地,增温试验使样地内地下生物量增加了89 g/m2.从0~10、10~20、20~30、30~40 cm土层的地下生物量分布来看,增温样地和对照样地的地下生物量主要分布在0~10 cm土层中.但从增温和对照样地内的分配比例上分析,各对应土层的分配比例存在差异.增温样地0~10 cm土层地下生物量为606 g/m2,高于对照样地的563 g/m2;20~30 cm和30~40 cm增温样地生物量与对照没有显著差异(P>0.05).

*和**代表增温和对的差异显著(P<0.05)和极显著(P<0.01).* and ** represent significant difference (P<0.05) and extremely significant difference (P<0.01) of warming and pair,the same as below.图1 温度升高对植物群落生物量分配的影响Figure 1 Effects of enhanced temperature on biomass distribution of plant communities
2.4 温度升高对植物群落物种多样性的影响
由表3可知,增温样地内Margalef 丰富度指数、Shnnon-Winener指数、Sinpson指数均高于对照样地,而Pielou指数差异并不显著(P>0.05).物种丰富度指数与特定群落的物种总数呈正比,说明增温使物种的数量明显增加.在试验进行前,通过对样地的调查显示,样地的本底相同.本底相同的样地在经过增温处理后,相似性指数较低,物种替代和消失的现象比较明显,对温度优势的物种逐渐变得优势更大,而对温度劣势的物种逐渐被淘汰,物种趋向于单一化.
2.5 温度升高对植物群落土壤养分的影响
由表3可知,增温样地内0~10、10~20、20~30、30~40 cm土壤养分含量均高于对照样地,OTC和CK样地土壤养分含量随着土壤深度的增加而逐渐降低趋势,在表层OTC和CK土壤平均温度相差较大,在30~40 cm,OTC和CK土壤养分含量基本一致,说明荒漠草原增温试验能够显著改变土壤养分含量,主要集中在表层土壤.

*和**代表增温和对的差异显著(P<0.05)和极显著(P<0.01).* and ** represent significant difference (P<0.05) and extremely significant difference (P<0.01) of warming and pair.图2 温度升高对植物群落物种多样性的影响Figure 2 Effects of enhanced temperature on plant community species diversity
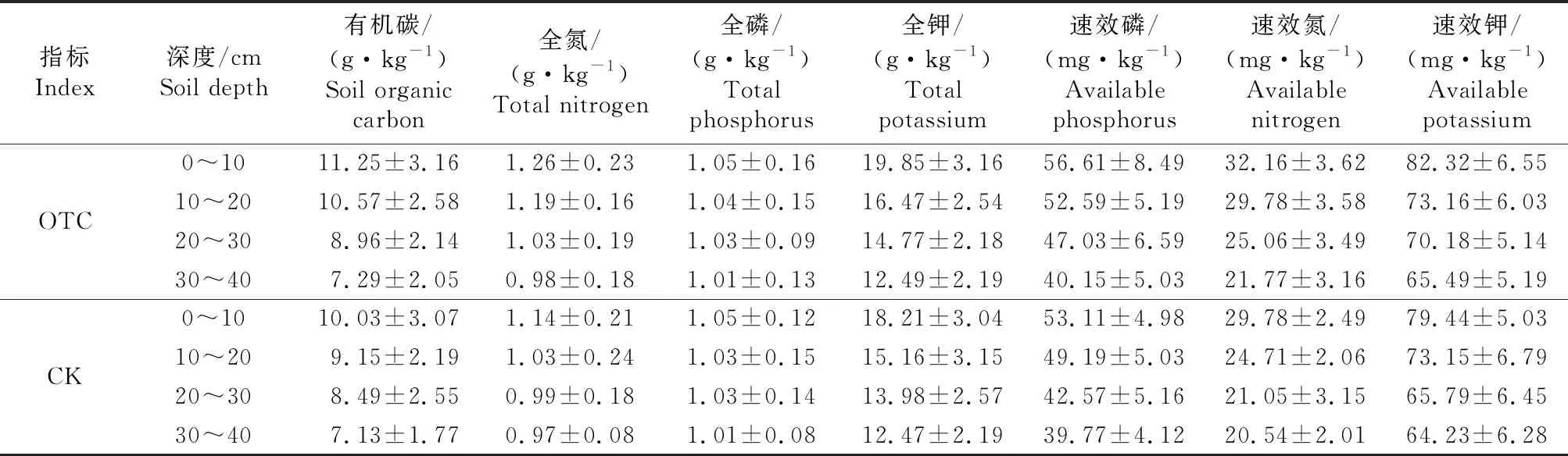
表3 温度升高对植物群落土壤养分的影响
2.6 温度升高对植物群落土壤酶活性的影响
由表4可知,增温样地内0~10、10~20、20~30、30~40 cm土壤酶活性均高于对照样地,OTC和CK样地土壤酶活性随着土壤深度的增加而逐渐降低趋势,在表层OTC和CK土壤平均温度相差较大,在30~40 cm,OTC和CK土壤酶活性基本一致,说明荒漠草原增温试验能够显著改变土壤酶活性,主要集中在表层土壤.
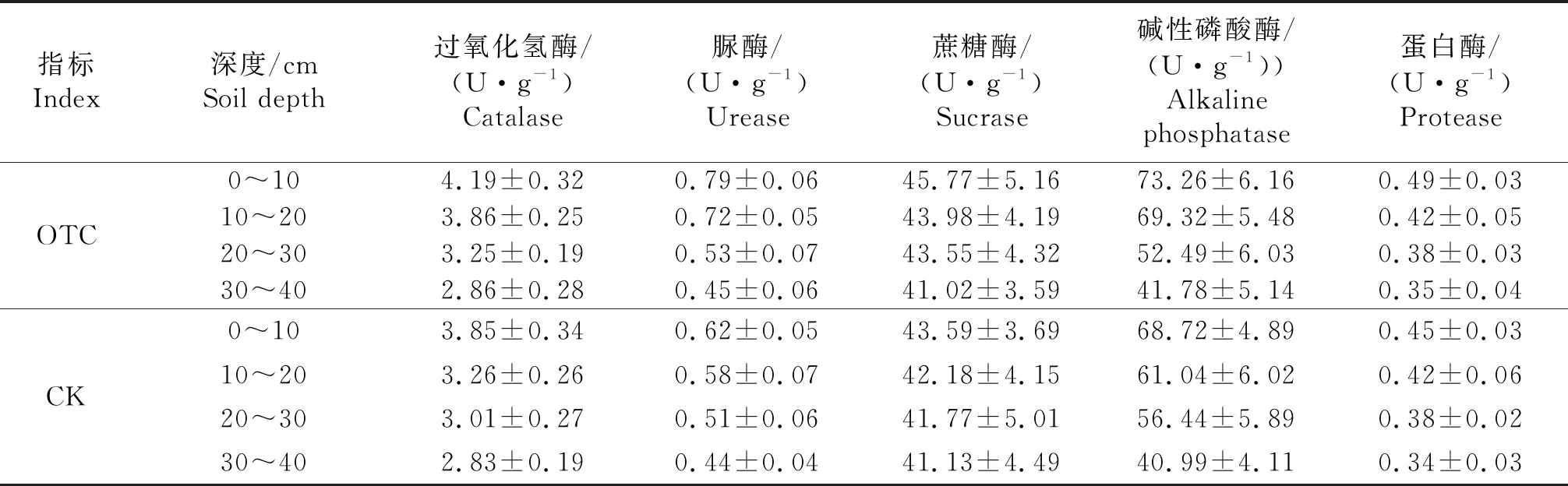
表4 温度升高对植物群落土壤酶活性的影响
2.7 植物群落多样性与土壤养分和酶活性的关系
相关性分析可知(表5),增温处理下地上生物量与有机碳呈显著正相关(P<0.05),枯落物量与有机碳和全氮呈显著正相关(P<0.05);地下生物量与有机碳呈极显著正相关(P<0.01),与全氮、全钾、速效磷和速效钾呈显著正相关(P<0.05);多样性指数(H)与有机碳呈极显著正相关(P<0.01),与全氮、全钾、速效磷、速效氮和速效钾呈显著正相关(P<0.05);丰富度指数(S)与有机碳呈极显著正相关(P<0.01),与全氮、全钾、速效磷、速效氮和速效钾呈显著正相关(P<0.05).
对照处理下地上生物量与有机碳呈显著正相关(P<0.05);地下生物量与有机碳呈极显著正相关(P<0.01),与全氮、全钾、速效磷和速效钾呈显著正相关(P<0.05);多样性指数(H)与有机碳和全氮呈极显著正相关(P<0.01),与全钾、速效磷、速效氮和速效钾呈显著正相关(P<0.05);丰富度指数(S)与有机碳、全氮、全钾、速效磷、速效氮和速效钾呈显著正相关(P<0.05).
由表6可知,增温处理下地上生物量和枯落物量与土壤酶活性(过氧化氢酶、脲酶、蔗糖酶、碱性磷酸酶和蛋白酶)均没有显著相关性(P>0.05);地下生物量与过氧化氢酶呈极显著正相关(P<0.01),与脲酶、蔗糖酶、碱性磷酸酶和蛋白酶呈显著正相关(P<0.05);多样性指数(H)和丰富度指数(S)与过氧化氢酶、脲酶呈极显著正相关(P<0.01),与蔗糖酶、碱性磷酸酶和蛋白酶呈显著正相关(P<0.05);均匀度指数(JP)和优势度指数(D)与土壤酶活性(过氧化氢酶、脲酶、蔗糖酶、碱性磷酸酶和蛋白酶)均没有显著相关性(P>0.05).

表5 植物群落多样性与土壤养分的相关关系
对照处理下地上生物量和枯落物量与土壤酶活性(过氧化氢酶、脲酶、蔗糖酶、碱性磷酸酶和蛋白酶)均没有显著相关性(P>0.05);地下生物量和多样性指数(H)与过氧化氢酶、脲酶呈极显著正相关(P<0.01),与蔗糖酶、碱性磷酸酶和蛋白酶呈显著正相关(P<0.05);丰富度指数(S)与过氧化氢酶呈极显著正相关(P<0.01),与脲酶、蔗糖酶、碱性磷酸酶和蛋白酶呈显著正相关(P<0.05);均匀度指数(JP)和优势度指数(D)与土壤酶活性(过氧化氢酶、脲酶、蔗糖酶、碱性磷酸酶和蛋白酶)均没有显著相关性(P>0.05).

表6 植物群落多样性与土壤酶活性的相关关系
3 讨论
3.1 增温对土壤温度和湿度的影响
在野外自然条件下采用远红外线加热的方法,能够模拟气候变暖.在本研究中,开顶式生长室(OTCs)起到了增温的作用.开顶式增温棚的设计,允许棚内外空气的自由交换流通,并尽可能减少不利的生态影响,如:低光照条件、温度极值、非自然降水、CO2浓度聚集及湿度的影响等[20].增温棚增温的原理如同温室效应,增温棚降低了风速,减弱了空气湍流的强度,使得温室内热量不容易散失,同时,由于太阳辐射很容易透过玻璃纤维,使得室内温度升高.本研究中,增温样地0~10、10~20、20~30、30~40 cm土壤平均温度均高于对照样地,从3月到11月,OTC和CK样地土壤平均温度呈一致的变化趋势,均呈先增加后降低趋势,在7月达到最大;增温样地土壤平均温度随着土壤深度的增加而逐渐降低趋势,在表层OTC和CK土壤平均温度相差较大,在30~40 cm,OTC和CK土壤平均温度基本一致;而平均含水量与土壤温度变化趋势相反,土壤说明荒漠草原增温试验能够显著改变土壤温度和含水量,主要集中在表层土壤.相对含水量的降低主要是由于增温棚内温度的升高导致.
3.2 增温对群落结构的影响
开顶式生长室(OTC)的增温效果引起植物群落变化.低温是限制荒漠草原植物生长的关键因素之一,OTC改善了植物群落的小气候环境,一定程度上满足了植物对热量的需求,有利于植物的生长和发育,对群落结构产生一定的影响[21-22].与此同时,温度的变化也将影响植物对水分和养分的吸收,间接影响植物的生长和生物量的积累.本研究中,增温样地和对照样地群落物种组成基本相同,说明增温作用没有明显的改变试验样地内植物群落组成,但是增温改变了各物种的重要值,在增温样地内物种重要值均高于对照样地.增温样地内Margalef 丰富度指数、Shnnon-Winener指数、Sinpson指数均高于对照样地,而Pielou指数差异并不显著(P>0.05),说明温度升高使荒漠草原植物群落的均匀度增加,但并没有提高草地植物的物种多样性.增温样地内0~10、10~20、20~30、30~40 cm土壤养分含量均高于对照样地,OTC和CK样地土壤养分含量随着土壤深度的增加而逐渐降低趋势,在表层OTC和CK土壤平均温度相差较大,在30~40 cm,OTC和CK土壤养分含量基本一致.这与增温导致的土壤含水量降低有关,由于增温降低了土壤含水量,使得水分成为限制植物生长的最关键因子,植物为了更好的适应环境,根系向更深的层次延伸寻求水分,地下生物量在向深层土壤转移,与前人的研究结果相一致[23].
3.3 增温对土壤养分和酶活性的影响
土壤酶催化土壤中的一切生物化学反应和物质循环,其活性大小反映生物化学过程的强度和方向,与土壤肥力密切相关.一般情况下,适度的增温会明显地提高土壤酶活性,这在很大程度上依赖于温度升高条件下土壤微生物数量的增加和植物根际分泌速度的提高.本研究中,连续5年的增温结果发现:增温样地内0~10、10~20、20~30、30~40 cm土壤酶活性均高于对照样地,OTC和CK样地土壤酶活性随着土壤深度的增加而逐渐降低趋势,在表层OTC和CK土壤平均温度相差较大,在30~40 cm,OTC和CK土壤酶活性基本一致,说明荒漠草原增温试验能够显著改变土壤酶活性,主要集中在表层土壤,这与前人的研究结果相似[24-26].此外,土壤酶活性对升高温度的响应还可能受土壤理化性质、水热状况、土壤代谢、植物种类及土壤生物区系、数量和生物多样性等因子的综合影响.
4 结论
内蒙古荒漠草原连续6 a的增温作用没有明显的改变植物群落组成,其中温度升高使荒漠草原植物群落的均匀度增加,但并没有提高草地植物的物种多样性.在土壤理化性质中,土壤有机碳和土壤全氮对土壤脲酶、过氧化氢酶活性的影响最强烈,温度升高增加了土壤有机碳,也使得微生物生物量和酶活性增加,说明土壤中的微生物和酶活性对温度的升高有一个适应的过程,也正是这些土壤生物通过调节自身活动带给整个陆地生态系统增强或削弱全球变化所带来的后果的可能性.相关性分析表明,丰富度指数和多样性指数均与土壤养分和酶活性呈显著正相关,而增温处理下的各相关系数高于对照.综合分析表明:长期增温并未改变荒漠草原短花针茅的群落组成,而提高了群落丰富度指数和多样性指数,同时增加了土壤养分和酶活性,而并未改变其垂直变化规律.
猜你喜欢
小哥白尼(趣味科学)(2022年5期)2022-08-15 08:34:46
现代园艺(2021年23期)2021-12-01 07:47:44
中国石化(2021年8期)2021-11-05 07:00:14
林业勘查设计(2020年1期)2021-01-18 02:40:48
新农业(2020年18期)2021-01-07 02:17:08
绿色中国(2019年14期)2019-11-26 07:11:34
现代园艺(2017年21期)2018-01-03 06:41:42
文史春秋(2016年3期)2016-12-01 05:42:20
绿色科技(2016年16期)2016-10-11 06:53:51
亚热带资源与环境学报(2015年1期)2015-01-22 07:04:58
