
云南松三种同域共存切梢小蠹梢转干期的空间分布格局
2020-07-17 10:25武承旭臧丽鹏张苏芳孔祥波徐芳玲黄桂英
生态学报 2020年11期
武承旭,臧丽鹏,张苏芳,孔祥波,刘 福,张 真,*,李 猷,徐芳玲,黄桂英
1 中国林业科学研究院森林生态环境与保护研究所,国家林业与草原局森林保护学重点实验室,北京 100091 2 贵州大学林学院,贵阳 550025 3 佛罗里达大学森林资源与保护学院,佛罗里达州 32611 4 云南省玉溪市红塔山自然保护区管理局,玉溪 650000
我国西南地区的3种重要切梢小蠹:横坑切梢小蠹Tomicusminor(Hartig)、云南切梢小蠹T.yunnanensis(Kirkendall and Faccoli)和短毛切梢小蠹T.brevipilosus(Eggers)均属切梢小蠹属,主要危害云南松Pinusyunnanensis等松属植物。3种切梢小蠹往往两两或者三者同域发生,协同危害寄主,加速寄主植物的死亡[1- 2]。切梢小蠹危害云南松主要包括两个阶段:蛀梢危害和蛀干危害[3]。首先,每年4—5月新羽化的成虫通过蛀食枝梢髓部补充营养直到性成熟[4- 5],在蛀梢后期(梢转干期)切梢小蠹有聚梢危害习性,造成云南松树梢枯黄易折,树势减弱,导致寄主植物抗性减弱[2,6];其次,小蠹在蛀干期繁殖形成的母坑道和子坑道的过程是导致寄主植物死亡的直接原因[4,7- 8]。
昆虫种群内个体间相互作用和对环境的适应,使其形成不同的空间分布格局[9]。研究昆虫空间格局对种群扩散、预测预报管理及持续防控具有重要意义[10- 11]。运用地统计学研究昆虫种群的空间分布格局已在国内外被广泛尝试[12- 17]。昆虫种群空间分布型常受种类生物学特性[18],发育阶段[19- 21],天敌[22- 23],寄主林分[18,24]和寄主生理生化[25],以及气候等环境因素的影响[10]。目前,有关切梢小蠹种群的空间分布格局研究主要集中在利用传统的聚集度指标法和回归模型法分析单个种群[3,26- 28],结果均表明不同危害程度云南松树冠或者树冠不同方向中切梢小蠹均呈聚集分布,聚集的原因与切梢小蠹本身的生活习性有关。但传统生物统计方法受限于空间位置[17]。尽管俞琳锋等[29]年采用地统计学将蛀梢期两种切梢小蠹的危害梢作为整体研究了其空间分布格局,但没有细化2种切梢小蠹在云南松林中的空间分布类型和差异。因此,3种切梢小蠹在同域共存下各自的空间分布型未曾研究。
本文将传统空间格局和地统计学的方法有机结合深入研究了发生在云南省大理祥云的切梢小蠹种群在同域共存情况下梢转干期的空间格局,为种群预测预报和综合调控提供技术支撑。
1 材料与方法
1.1 试验地基本概括
试验样地位于云南省大理祥云普淜镇(N25°20′25.96″, E100°54′31.43″),海拔约2000 m,林分为人工营造云南松纯林,树龄10年,树高3—8 m,胸径为4—13 cm。林地内的郁闭度为0.5左右,株距为3—7 m,当地气候属于干、湿季节分明的北亚热带高原季风型中温带气候,5—9月为雨季,降水集中;10月至翌年4月为干季,光照充足。常年主要风向为西南风。该地常年受横坑切梢小蠹、云南切梢小蠹和短毛切梢小蠹混合危害,其中短毛切梢小蠹种群数量占前两者约百分之一。
1.2 研究方法
2016年10—11月在祥云县大理祥云普淜镇云南松林中根据林业有害生物发生及成灾标准(轻度受害林枯梢率:10%—20%;中度受害林枯梢率:20%—50%; 重度受害林枯梢率:>50%)选择3块不同受害程度的50 m×50 m的云南松林试验标准地,其中轻度、中度和重度受害样地中3种切梢小蠹的枝梢危害率分别为19.89±1.27,23.02±4.52和51.19±3.06。文中采用每木检尺方法调查各样地切梢小蠹在云南松的空间分布格局。随机抽取每株云南松上中下每层枝梢25梢以上,调查记录各层各梢中切梢小蠹数量,以每100梢虫量为密度指标。
在调查过程中,同时记录采集样地树木的高度(m),胸径(cm)和枝梢在树上的分布位置。同时记录样地内各云南松的行距,将每株云南松的相对空间位置绘置于坐标纸上,用于半方差分析。将采集的切梢小蠹按照样地及分层分别放入带孔离心管(50 mL)同枝梢一起带回实验室,然后鉴定3种切梢小蠹的种类和雌雄,并确定各小蠹在树梢中的位置。鉴定方法同根据Kirkendall等[30],李霞等[31]和王平彦等[32]。由于3种切梢小蠹雌、雄虫在蛀梢期的种群数量高峰期存在差异,所以研究中将雌、雄成虫区别研究。
1.3 分析方法
1.3.1聚集度指标法和回归模型法分析
采用传统生物学统计方法计算每样地各层树冠中每100梢的平均切梢小蠹数量(样本均数)m(头/100梢),并采用平均拥挤度(m*)、扩散系数(C)、丛生分布指数(I)、Cassie指标(CA)、负二项分布指标(k)、Lioyd 聚块性指数(m*/m)[33]等进行计算3种切梢小蠹在蛀梢后期的空间分布格局。
Taylor幂法则:即样本平均数m与方差S2对数值之间的回归关系公式:lgS2=lgα+blgm(α,β为常数)。其中α与样本大小和计算方法有关,受环境异质性影响;b为种群聚集对密度依赖性的特征指数。当b趋向0时为均匀分布;b=1时为随机分布;b>1时为聚集分布。
Iwao回归分析法m*-m:回归模型为m*=α+βm,式中,m*平均拥挤度和m平均密度,α为分布的基本成分按大小分布的平均拥挤度,β向0时为均匀分布;b=1时为随机分布;b>1时为聚集分布。β为基本成分的空间分布型。当α=0时,种群分布均匀,分布的基本成分为单个个体;当α>0时,个体间相互吸引,分布的基本成分为个体群;当α<0时,个体间相互排斥。当β=1时,种群为随机分布;当β>1时,种群为聚集分布;当β<1时,种群为均匀分布。
聚集原因检验:应用Blackith种群聚集均数λ=mγ/2k分析聚集原因,式中,m为平均虫口密度,k为负二项分布的指数,γ是自由度为2k,以0.5概率值对应处的χ2值,应用内插法得出;当λ<2时,说明聚集是由一些环境条件引起;当λ≥2时,说明聚集是由昆虫的自身行为和环境因素综合影响的结果。
1.3.2半方差函数分析
(1)半方差函数(Semivariogram)是指区域化变量Z(xi)和Z(xi+h)区域化变量增量的方差,其既能描述区域化变量的空间结构,又能描述随机性变化。其计算公式如下所示:
式中,γ(h)为半方差函数值,即相隔距离为h的半方差函数值。半方差函数既是距离h函数,又是方向α的函数。半方差函数曲线图是半方差函数γ(h)对距离h的坐标图形。N(h)为被h分隔的数据对的数量(相隔距离为h的所有点的配对数),Z(xi)和Z(xi+h)分别是在点xi和xi+h处样本的测量值,h是两分隔样点的距离。
基台值(Sill)、块金常数(Nugget)和变程(Range)是变异曲线图中3个主要特征参数[16]。基台值表明达到平衡时的变异函数值,其可以反映变量在空间上的总体变异水平;块金常数表示原点处变异函数的不连续性,代表了最小抽样尺度下观测误差等导致的随机变化。变程是当变异函数的值达到平衡时的间隔距离,它反映了区域化变量的相互影响范围的大小,变程之外各数据之间互不相关。
(2)变异函数理论模型的拟合,常用来拟合实际变异曲线图的理论模型有(a)线性模型Linear model:表示种群的空间分布为随机分布或均匀分布;(b)指数模型Exponential model;(c)球状模型Spherical model;(d)高斯模型Gaussian model。后3种模型均表示种群的空间分布为聚集分布。在选择最优拟合模型时分别先后需要考虑决定系数R2,残差 RSS,变程和块金常数的大小,以期确定理论模型的优劣程度[16,29]。
采用本协同处理方案,将产生的浓缩液、污泥、沼气协同入炉焚烧,均匀回喷到3台焚烧炉内。浓缩液高温分解,同时也起到降低出口烟温的作用,二次风量减少至12000m3/h,保证出口氧浓度为6.23%,比设计工况下二次风量降低13 000 m3/h,既保证完全燃烧,满足环保要求,又降低了排烟损失,提高发电量。
(3)本文使用 GS+7.0软件对受害梢的空间分布格局进行地统计学分析。所有种群密度数据采用样地内单位云南松100梢的切梢小蠹虫量为指标。空间分布图在GS+7.0软件中运用Kriging插值法进行插值分析,并生成空间等值线分布图。
2 结果与分析
2.1 3种切梢小蠹种群危害和分布状况
在云南松林中,严重受害样地中梢转干期的横坑切梢小蠹,云南切梢小蠹种群数量均显著高于中度和轻度受害样地(表1)(横坑切梢小蠹:F(2,897)= 17.452,P< 0.001; 云南切梢小蠹:F(2,897)= 105.945,P< 0.001),但短毛切梢小蠹则差异不显著(F(2,897)= 1.196,P= 0.303)。在重度受害样地中云南切梢小蠹的种群密度显著高于横坑切梢小蠹(F(2,897)= 212.371,P< 0.001),前者种群密度可达15.87头/100梢,然而在轻度受害样地中两者种群数量则相反(F(2,897)= 90.866,P< 0.001)。3种切梢小蠹的种群密度在两性间差异均不显著,可能存在偏差(表1)(P> 0.05)。

表1 3种同域切梢小蠹雌雄虫在不同受害程度云南松纯林样地中的种群密度
2.2 3种切梢小蠹在不同受害云南松林的种群空间格局
2.2.1聚集度指标
横坑切梢小蠹,云南切梢小蠹和短毛切梢小蠹在不同受害程度云南松树冠的聚集度指标如表2所示,结果表明横坑切梢小蠹的平均拥挤度m*介于10.04—20.02之间,云南切梢小蠹的平均拥挤度m*位于9.34—31.91之间,而短毛切梢小蠹的则是3.25—8.10;扩散系数C均大于1,丛生指数I大0,Cassie指数CA都大于0,Lioyd聚块性指数m*/m大于1,负二项分布系数均大于0,均说明三种切梢小蠹在蛀梢期呈聚集性分布。
根据负二项分布系数越小,聚集性越强,可以看出横坑切梢小蠹在重度受害样地的负二项分布系数均小于云南切梢小蠹,说明当云南松被严重危害后,横坑切梢小蠹聚梢能力更强。但在中度和轻度受害样地,两者聚集性则相反。其中样地中短毛切梢小蠹的负二项分布系数均最小,可能是因为总体样本量较小或者聚集性行为等其他原因形成。
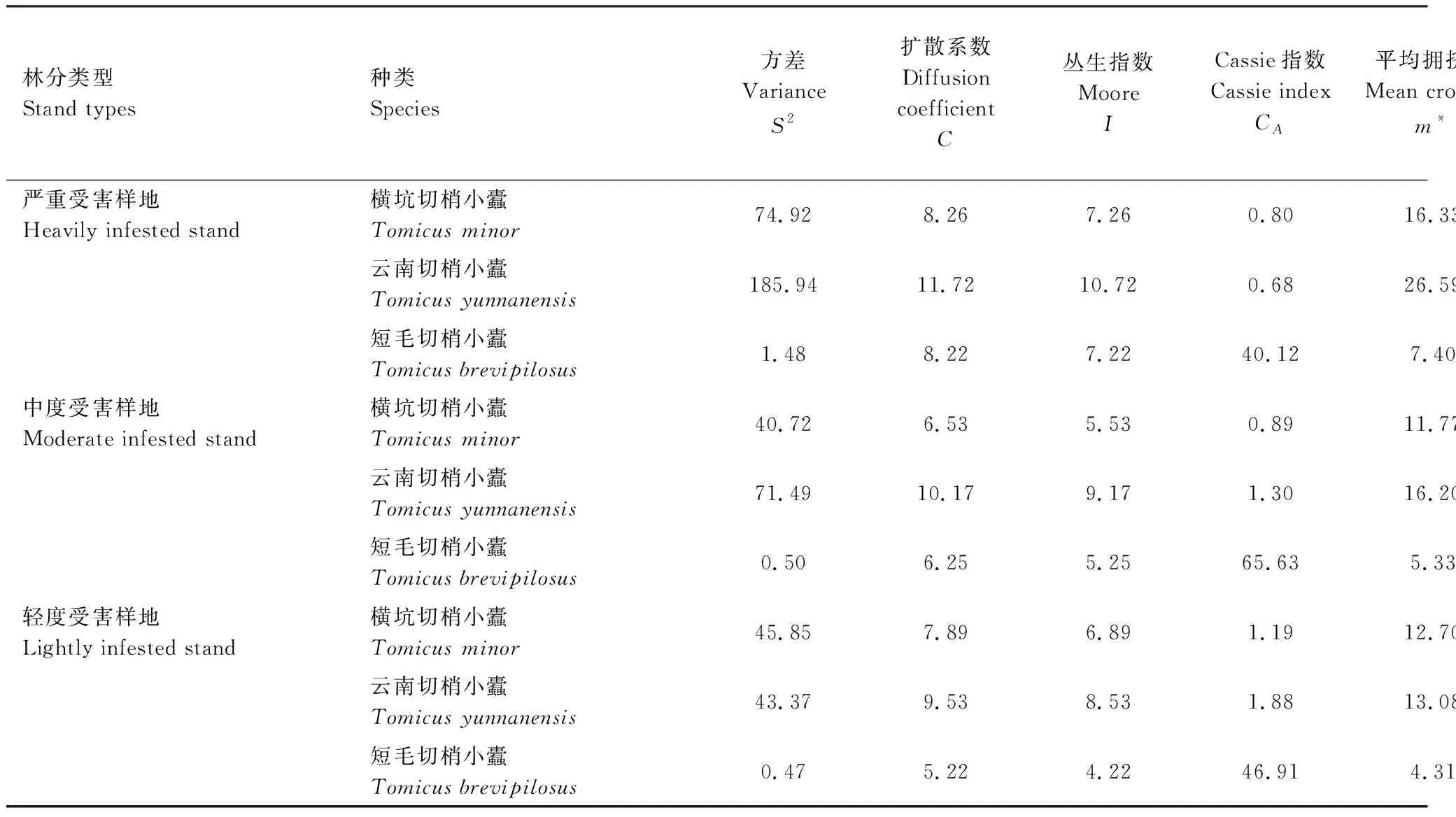
表2 3种切梢小蠹在蛀梢后期的空间分布格局各项参数
2.2.2回归模型分析
Taylor 幂法则:横坑切梢小蠹,云南切梢小蠹和短毛切梢小蠹的S2-m的回归直线方程分别为lg(S2)=0.6443+1.2765lg(m) (R=0.9446);lg(S2)=0.8685+1.1663lg(m) (R=1.0000*)和lg(S2)=1.2406+1.4478lg(m) (R=0.9833),其中lgα均大于1,β均大于1,说明3种切梢小蠹成虫在梢转干期云南松树冠中在任何密度下均呈聚集分布,且具有密度依赖性。m值越高,聚集程度越高。
Iwao 的m*-m回归分析:横坑切梢小蠹,云南切梢小蠹和短毛切梢小蠹的m*-m回归关系分别为m*=4.4932+1.2935m(R=0.9502);m*=7.7502+1.1882m(R=0.9999*)和m*=2.6357+26.1068m(R=0.9131),其中α均大于1,β均大于1,说明3种切梢小蠹在不同竞争密度下的云南松树冠中个体间互相吸引,分布型均呈聚集分布。
聚集原因分析:横坑切梢小蠹的λ值介于15.51—22.29之间,云南切梢小蠹的λ值位于12.77—38.36,均大于2,说明两者聚集是由环境因素和昆虫本身的聚集习性引起;而短毛切梢小蠹的λ值是1.67—1.92,小于2,说明是由环境因素引起。
2.3 地统计学分析
采用GS+软件中的地统计学方法对同域危害云南松的3种切梢小蠹的空间分布格局进行了分析,结果表明,除了重度受害云南松样地中短毛切梢小蠹种群为随机分布外,其余各种群均具有空间依赖性,呈现出聚集分布格局(表3,图1)。

图1 同域竞争共存的横坑切梢小蠹,云南切梢小蠹和短毛切梢小蠹种群数量在重度,中度和轻度受害云南松纯林的等值线图Fig.1 Isoline map of the population density for Tomicus minor, Tomicus yunnanensis and Tomicus brevipilosus at heavily infested, moderate infested, and lightly infested stands of pure Pinus yunnanensis forest, respectively
在重度受害样地中横坑切梢小蠹的空间依赖范围最大,为169.30 m,超过了调查空间范围,说明调查范围内各点的横坑切梢小蠹均具有相关性,其余(除去重度受害样地中短毛切梢小蠹)均在4.01—7.45 m(表3),说明在此范围内抽样空间内任意两点种群数量均存在着关联性(图2)。重度受害样地中横坑切梢小蠹和云南切梢小蠹种群的变异函数中C值占基台值的比例(空间变异值)为100%和98%,说明总空间变异由空间自相关现象产生的可达100%,同样现象呈现在轻度受害样地中的3种切梢小蠹(表3)。
在中度和轻度受害云南松林中同域竞争共存的短毛切梢小蠹种群数量最少,占百分之一,其种群密度分布呈若干板块状(图1),但在重度受害样地呈现出较弱的聚集性,接近随机分布,种群个体间相互独立。然而,横坑切梢小蠹和云南切梢小蠹种群在3个样地中基本呈板块状分布,存在若干个聚集点(图1)。其中重度受害样地中的云南切梢小蠹和中度受害样地中的横坑切梢小蠹的决定系数分别为0.87和0.88,说明此时2种切梢小蠹各自的空间分布具有较强的空间依赖性。
由3种受害样地的3种切梢小蠹种群数量的等值线图可以看出重度受害样地中横坑切梢小蠹种群存在从西北向东南扩散趋势(图1),而在中度危害样地中由南北两侧向中心延伸(图1)。云南切梢小蠹种群在中度受害云南松样地中则由两侧向中间扩散(图1),在轻度受害样地北部聚集密度大,向南存在2—3个聚集点后,其聚集度逐渐降低(图1),然而其在重度受害样地整块区域均有聚集点,聚集程度大,聚集面积广,危害最为严重(图1)。
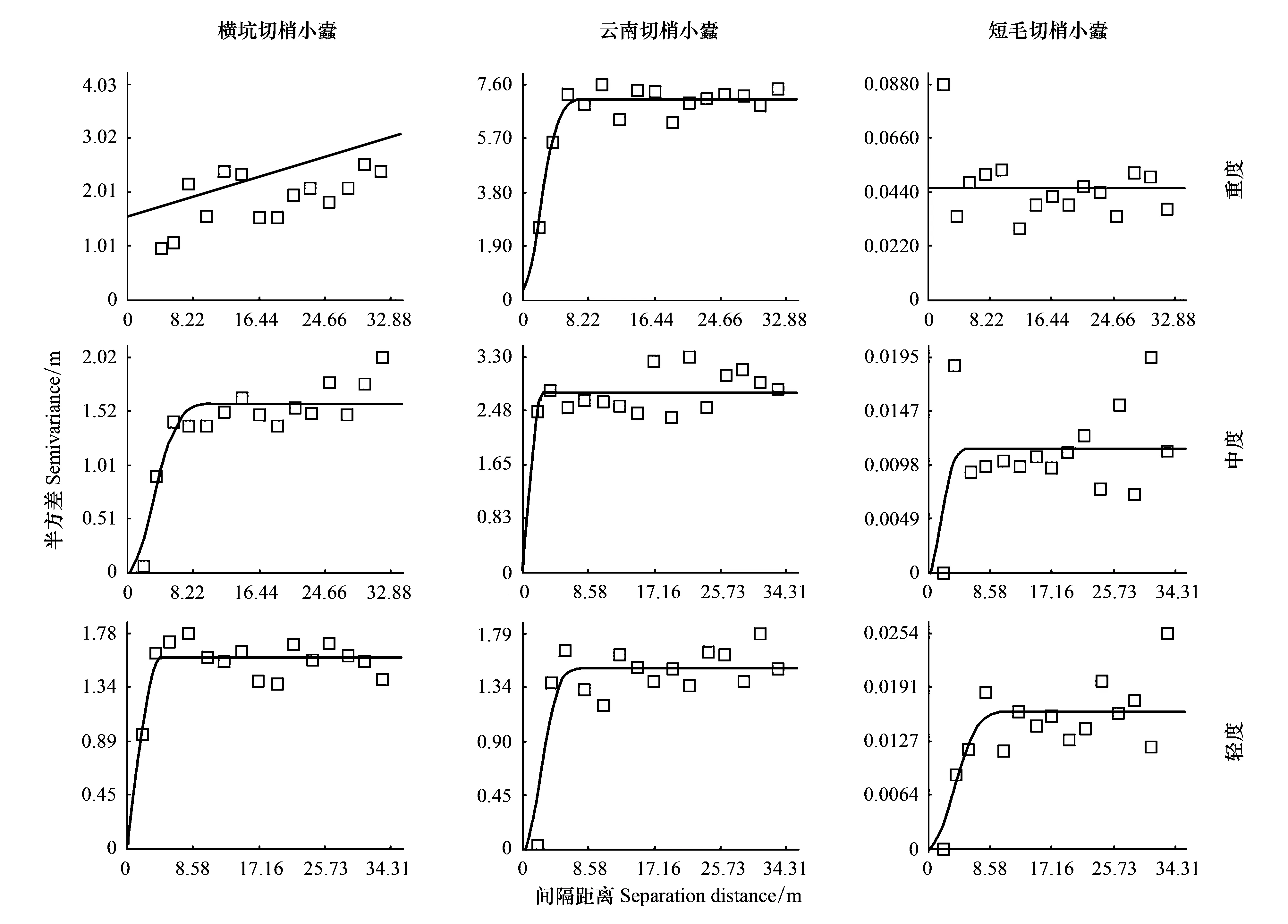
图2 同域竞争共存的横坑切梢小蠹,云南切梢小蠹和短毛切梢小蠹种群数量在重度,中度和轻度受害云南松纯林的变异函数图Fig.2 Variogram of the population density for Tomicus minor, Tomicus yunnanensis and Tomicus brevipilosus at heavily infested, moderate infested, and lightly infested stands of pure Pinus yunnanensis forest, respectively
3 结论与讨论
空间分布型是切梢小蠹种群的重要属性之一,能够揭示昆虫聚集强度和空间结构异质性程度[34]。传统的空间格局研究方法如聚集指标法仅强调样本间的数量变化,忽略了样本的空间位置和程度,无法解释昆虫种群的空间相关性,依赖性和连续性;而地统计学揭示了昆虫种群在不同方向上的差异,且受样方大小、取样位置和样本容量大小的影响较小[35],但缺陷是仅考虑时刻变化的昆虫种群[34]。两者相互补充,一方面有助于全面准确地了解切梢小蠹的空间分布规律,还可为害虫预测预报和综合防治提供理论基础。
本文结果进一步证实同域共存的横坑切梢小蠹和云南切梢小蠹雌、雄虫在蛀梢后期云南松树冠中均呈现出聚集分布,这与单个切梢小蠹种群存在时的空间生态格局一致[3,26- 28],说明同域共存不影响其分布类型,但影响其空间变异。昆虫种群内和种群间的空间相关一方面受空间因素影响,另一方面受种群密度制约[34],切梢小蠹的聚集程度随着种群密度的增加而增强,个体间相互吸引,种群高密度时甚至出现一梢多虫,一母坑道多雄虫现象,但各蛀孔间互不相通[36];在重度受害样地,短毛切梢小蠹种群密度小,仅占其他2种切梢小蠹种群数量的百分之一,采用传统生物统计学方法分析结果显示其为聚集分布,但地统计学分析结果发现其空间分布型为随机分布,说明其具有不稳定性。
在相似小生境中昆虫的空间分布取决于其生物学特征和栖息环境[11]。由于梢转干期是切梢小蠹从蛀梢期到蛀干期的过渡阶段[26],通常切梢小蠹在这一时期大量聚梢取食[3]。本研究调查的3块样地林分类型,立地条件均较为相似,但各样地中3种切梢小蠹的种群密度差异较大,本文研究结果表明重度受害样地中的云南切梢小蠹种群数量高于横坑切梢小蠹,其空间依赖性更大,由于切梢小蠹聚梢危害,这说明各自蛀梢时间跨度和高峰期不同[37]。在中度受害样地,横坑切梢小蠹相关性最高,说明此时期其种群可能处于上升期。3种切梢小蠹在不同受害样地的种群数量和比例有差异,这也符合地统计学结果,即变异函数理论模型有差异,如同种切梢小蠹在不同受害程度样地中的变异函数理论模型在球形模型和高斯模型之间转换,而且空间依赖程度也不同。3种切梢小蠹种群的空间分布等值线图表明切梢小蠹的聚集程度不同,其扩散方向和形式也不同,这可能受环境和种间生物学差异影响,如切梢小蠹聚集能力的不同可能与它们对聚集信息物质的依赖程度和反应强度不同有关,进一步说明切梢小蠹的聚集性和不稳定性。
林分受害程度也影响昆虫种群的空间分布格局[17]。俞琳锋等[29]研究发现切梢小蠹混合种群在轻度受害林分内危害梢的空间依赖范围要大于中度受害和重度受害林分内的空间依赖范围,而我们的研究结果表明云南切梢小蠹的空间依赖范围比较稳定,但横坑切梢小蠹和短毛切梢小蠹的空间依赖范围受林分受害程度影响很大。叶辉和李隆术[3]研究表明在中度受害的云南松林中,蛀梢期松纵坑切梢小蠹在云南松树梢上存在聚集危害的习性,与我们研究得到的在中度受害林中受害梢里面切梢小蠹的空间分布为聚集分布一致。
传统聚集指标法和地统计学方法表明同域共存关系不影响横坑切梢小蠹和云南切梢小蠹在云南松梢转干期空间分布类型,均呈聚集分布。切梢小蠹在这一时期大量聚梢危害能够短时间内克服寄主抗性,削弱树势,为蛀干期繁殖创造条件[6]。这种现象可能与小蠹化学信息物质(聚集信息素)和寄主挥发物的增效作用密切相关[38]。在空间大尺度下,小蠹依靠寄主挥发物长距离定位寄主[39],如很多次生性小蠹和食菌小蠹对树势衰弱的树有明显的趋性,因为这类型树产生的乙醇对其定位起关键作用[40]。有研究表明切梢小蠹对被害云南松树梢挥发物有明显的趋性,且其对自身为害的枝梢萃取物,抽提物均有强烈的趋性[41],充分说明信息化学物质(寄主挥发物)可能是切梢小蠹聚集危害云南松树梢一个重要的识别信号,然而小蠹聚梢危害的详细行为和机制仍需进一步研究。
猜你喜欢
林业资源管理(2022年6期)2023-01-18
今日农业(2022年15期)2022-09-20
现代园艺(2021年23期)2021-12-01
湖南电力(2021年1期)2021-04-13
新农业(2020年18期)2021-01-07
花卉(2020年24期)2021-01-04
种子(2019年5期)2019-07-02
乡村科技(2019年9期)2019-02-22
山西农业科学(2019年10期)2019-02-12
红土地(2018年7期)2018-09-26
