
一株球孢白僵菌的分离鉴定及其对草地贪夜蛾的致病性
2020-07-07 01:35雷妍圆章玉苹薛志洪王裕华黄少华吕利华
环境昆虫学报 2020年3期
雷妍圆,章玉苹*,薛志洪,王裕华,黄少华,吕利华
(1. 广东省农业科学院植物保护研究所/广东省植物保护新技术重点实验室,广州 510640;2. 广东省农业科学院,广州 510640;3. 华南农业大学农学院,广州 510642)
草地贪夜蛾Spodopterafrugiperda是联合国粮食与农业组织(FAO)预警的跨国界迁飞性害虫,已在美洲、非洲及亚洲等近100个国家迅速扩散,对当地玉米、水稻、甘蔗、高粱、棉花、花生、大豆等作物造成了巨大损失(Casmuzeetal., 2010; Earlyetal., 2018; 姜玉英等,2019;张磊等,2019)。自2019年1月以来,该虫从境外迁入我国西南、华南地区,短短数月迅速向北蔓延,至目前发生区域已涉及全国近20多个省份,对我国农业生产构成严重威胁(廖永林等,2019;王磊等,2019;杨普云等,2019)。
目前草地贪夜蛾防控主要依赖化学农药,但该虫对农药具有较强抗性(Yuetal., 2003; Ríos-Díez & Saldamando-Benjumea, 2011; Carvalhoetal., 2013; 李永平等,2019),化学防治易产生农药残留,对人、畜和非靶标生物造成伤害。植保工作者在其物理、生物防治方面做了很多工作,到目前为止,仍未有一种生防病原真菌能对草地贪夜蛾起到完全的控制效果。然而,昆虫病原真菌以其寄主范围广、扩散力强、易于商品化生产和可持续性控制虫害等优势在对草地贪夜蛾的防治中具有重要的应用前景(Shah & Pell, 2003; 张礼生和陈红印,2014;Rivero-Borjaetal., 2018; 陈万斌等,2019;张维等,2019)。由于草地贪夜蛾入侵我国时间较短,对其微生物防治研究起步较晚,筛选对草地贪夜蛾高特异性和高效率的病原真菌,大力推动微生物杀虫剂的研究与利用,是我国草地贪夜蛾可持续防控工作的重要任务。
在昆虫病原真菌防控草地贪夜蛾方面,国内外报道球孢白僵菌Beauveriabassiana、金龟子绿僵菌Metarhiziumanisopliae和莱氏绿僵菌Metarhiziumrileyi对草地贪夜蛾卵和低龄幼虫有一定的防治效果(Lezama Gutiérrezetal., 1996, 2001; Carneiroetal., 2008; Garcíaetal., 2011; Thomazonietal., 2014; Akutseetal., 2019; 郑亚强等,2019)。玫烟色虫草Cordycepsfumosorosea[原玫烟色棒束孢Isariafumosorosea(Kepleretal., 2017)]可致病幼虫各龄期,对低龄防效最佳(Lezama Gutiérrezetal., 2001; 雷妍圆等,2020)。从已报道的菌株来看,大部分仅对草地贪夜蛾卵和低龄幼虫防效较好,对3龄以上高龄幼虫及其他虫态的防效普遍较差。白僵菌Beauveriaspp.是100多属昆虫病原真菌中最常见和最重要的属之一,在我国现已发现的所有白僵菌种之中,球孢白僵菌作为一种广谱性微生物杀虫剂,在农业害虫防治中具有重要地位(Hajek & St. Leger, 1994)。我国利用球孢白僵菌在马尾松毛虫和玉米螟防治中的应用也成为了世界上最大规模的真菌杀虫剂应用项目(李增智,2015)。本研究室从广州地区野外采集的感菌稻黑蝽上分离纯化获得1株病原真菌,对其进行了形态学鉴定和ITS-rDNA序列分析,通过致病性测定,确定其最佳杀虫活性的孢子悬浮液浓度,以期为寻求对草地贪夜蛾更加持续且高效的生物防治方法提供研究材料。
1 材料与方法
1.1 供试虫源和菌株分离培养
草地贪夜蛾幼虫采集于广东省广州市白云区钟落潭镇广东省农业科学院白云基地“粤甜28”甜玉米ZeamaysL.植株上,在实验室用玉米叶和果穗饲养至化蛹,待其羽化产卵后建立实验室种群作为供试虫源。室内饲养条件为26±1℃,相对湿度60%~90%,光照条件14 L ∶10 D。
昆虫病原真菌采自广东省广州市华南农业大学湿地狐尾藻上罹病稻黑蝽Scotinopharalurida若虫僵虫(图2 A),将带菌虫体置于灭菌培养皿(d=9 cm),在超净工作台中用接种针轻轻挑起少量孢子,采用划线法接种于萨氏培养基SDAY(葡萄糖40 g,蛋白胨10 g,酵母膏10 g,琼脂15~20 g,蒸馏水1 L),于28±1℃,全黑暗,相对湿度70%的恒温培养箱中培养。培养6 d后,挑取少量孢子至新的培养基纯化培养15 d,平板上长出的菌落形态特征一致,将该菌编号为GZSL-1。将纯化的菌株分生孢子置于20%甘油,并保存于-80℃冰箱中。
1.2 菌株培养形态观察
分离纯化的菌株GZSL-1接种于PDA培养基(马铃薯200 g,葡萄糖20 g,琼脂15~20 g,蒸馏水1 L),于28±1℃,全黑暗,相对湿度70%的恒温培养箱中培养10 d。采用光学显微镜(Axio Scope A1,Zeiss)和扫描电镜(S-3400N,Hitachi)观察菌落形态特征、菌株产孢结构和分生孢子形态等显微结构。
1.3 菌株分子生物学鉴定
以菌株GZSL-1基因组DNA为模板,采用真菌通用引物ITS1(5′-TCCGTAGGTGAACCTGCGG-3′)/ITS4(5′-TCCTCCGCTTATTGATATGC-3′)进行菌株rDNA-ITS序列PCR扩增。PCR反应体系为25 μL,其中上下游引物各1.5 μL,2×Taq PCR Master Mix 10 μL,模板DNA 2 μL,ddH2O 10 μL。PCR反应条件:94℃预变性5 min;94℃变性45 s,55℃退火30 s;72℃延伸1 min,30个循环;72℃延伸5 min。PCR产物经1%琼脂糖凝胶电泳检测后送北京擎科新业生物技术有限公司测序。
1.4 系统发育分析
对菌株rDNA-ITS测序结果进行编辑,去除序列两端质量不好的碱基,将优化好的ITS序列提交到NCBI网站(https://blast.ncbi.nlm.nih.gov/Blast.cgi),通过BLAST程序进行比对,得到与目的菌株具有同源性的多个菌株相应序列,从中下载同源性较高的序列,使用MEGA 7.0软件,通过邻接法(Neighbor-Joining method,NJ),运行1 000 次 bootstrap验证,构建系统发育树(Kumaretal., 2004)。
1.5 菌株对草地贪夜蛾幼虫侵染症状观察
从SDAY平板上收集菌株GZSL-1分生孢子,以灭菌0.05%吐温-80溶液配制1×108孢子/mL浓度孢子悬浮液,分别接种草地贪夜蛾1~6龄幼虫,接种后每天观察幼虫存活情况及感染病虫的外部特征,将死亡幼虫保湿培养,虫体表面长出白色菌丝或分生孢子的视为感菌致死。
1.6 菌株对草地贪夜蛾幼虫的致病力测定
采用浸虫法处理草地贪夜蛾幼虫,以灭菌0.05%吐温-80溶液配制1×104、1×105、1×106、1×107、和1×108孢子/mL共5个浓度梯度的孢子悬浮液,每个浓度为一个处理,以灭菌0.05%吐温-80溶液处理为对照。选取个体大小一致的1~6龄幼虫,放入供试孢子浓度的悬浮液中,浸渍10 s后挑出,置于滤纸上吸去多余水分,移至皿底垫有湿润滤纸片的培养皿(d=7.5 cm)中。为避免高龄幼虫自相残杀,1~2龄幼虫高密度集体饲养(10头/皿),3龄幼虫低密度集体饲养(5头/皿),4~6龄幼虫单头饲养(1头/皿),皿内放入充足的新鲜玉米叶供其取食(王道通等,2019)。处理后的幼虫置于人工气候箱中饲养(26±1℃,RH 80%±5%,14 L ∶10 D)。每个处理20~30头幼虫,重复3次,持续观察6 d,每天记录幼虫死亡数,对死亡幼虫进行保湿观察,确定是否为感菌致死。
1.7 数据处理
累计死亡率(%)=(处理死亡总虫数/处理总虫数)×100
累计校正死亡率(%)=(处理累计死亡率-对照累计死亡率)/(1-对照累计死亡率)×100
实验数据经Excel整理后,使用SPSS 20.0软件进行处理分析,统计各处理幼虫的累计死亡率和累计校正死亡率。采用Probit方法计算致死中时(LT50),求回归方程及计算致死中浓度(LC50)。
2 结果与分析
2.1 菌株培养形态特征及分子生物学鉴定
对分离纯化培养的菌株形态特征进行鉴定,观察菌株平板正面菌落培养初期呈乳白色,后略变为淡沙黄色,孢子层厚且均匀,菌落边缘孢子较少,菌落生长速度快,在PDA培养基上28℃培养7 d时,菌落直径达到24.5 mm,菌落中心至边缘1/2处凸起同心环(图2 B)。菌落背面黄色,无深褐色环(图2 C)。营养菌丝无色,分生孢子梗着生于营养菌丝,分生孢子在菌丝或泡囊上簇生,着生在产孢细胞延伸而成的“之”字形结构上;分生孢子透明、光滑,球形或近球形,直径2.0~2.3 μm(图2 D)。
使用PCR扩增目的菌株的rDNA-ITS序列片段,测序结果显示扩增片段为498 bp,将该序列在GenBank数据库中进行BLAST比对,发现目的菌株的ITS序列与已报道的多个球孢白僵菌菌株对应序列的相似性均达到99%及以上。选取相关序列,使用MEGA 7.0将其在NCBI数据库中比对得到的序列下载构建系统发育树(图1)。本目的菌株与B.bassianaTNS/DWD/BB05(MK346250.1)处于进化树最小分支,亲缘关系最近,同源性最高。结合形态学鉴定结果,确定该菌为球孢白僵菌,编号GZSL-1菌株。

图1 基于ITS基因序列构建目的菌株与其他球孢白僵菌的系统发育树(NJ法)Fig.1 Construction of phylogenetic tree of the isolated strain and other related Beauveria bassiana strains based on ITS region sequence (Neighbour-Joining method)注:括号内为菌株在GenBank中的登录号;各分枝上的数字表示支持率。Note: The numbers in parentheses represent GenBank accession numbers. The numbers in each branch points denote the percentages supported by bootstrap.
2.2 感染球孢白僵菌GZSL-1的草地贪夜蛾幼虫症状
草地贪夜蛾各龄幼虫皆可被菌株GZSL-1感染(图2 E-J)。在感菌初期,幼虫取食行为、虫体外部形态与健康幼虫无差别。接种2 d后,低龄幼虫(1~3龄)幼虫取食明显减少或停止取食,部分幼虫死亡。接种后第4天,各龄期幼虫皆出现球孢白僵菌感染致死个体。感菌的低龄幼虫体表覆盖菌丝和分生孢子,分生孢子呈白色。高龄幼虫(4~6龄)感菌死亡时间相对低龄较晚,体表被白色菌丝包裹。
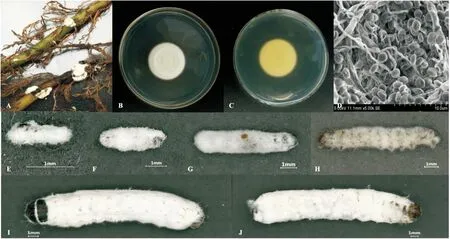
图2 菌株GZSL-1培养特征和草地贪夜蛾幼虫感染症状Fig.2 Morphological characteristics of strain GZSL-1 on PDA plate and external symptoms of larvae of Spodoptera frugiperda after infection treatment注:A,野外被球孢白僵菌侵染的稻黑蝽若虫僵虫;B,菌落正面;C,菌落背面;D,分生孢子和菌丝体;E,1龄幼虫;F,2龄幼虫;G,3龄幼虫;H,4龄幼虫;I,5龄幼虫;J,6龄幼虫。Note: A, symptoms of infected Scotinophara lurida nymphs in field; B, colony on the positive; C, colony on the back; D, conidia and mycelium; E, 1st instar larva; F, 2nd instar larva; G, 3rd instar larva; H, 4th instar larva; I, 5th instar larva; J, 6th instar larva.
2.3 球孢白僵菌GZSL-1对草地贪夜蛾幼虫的致病力测定
用5个浓度球孢白僵菌GZSL-1孢子悬浮液处理草地贪夜蛾1~6龄幼虫,随着孢子浓度增加和时间推移,幼虫累计死亡率逐渐升高(图3)。以1×104孢子/mL浓度处理,6 d后1~6龄幼虫的累计校正死亡率分别为76.47%、65.12%、48.24%、36.78%、13.48%和5.75%。随着浓度的升高,菌株对各龄幼虫的致死率逐渐升高。当孢子浓度为1×108孢子/mL时,1至3龄幼虫的累计校正死亡率均达100%,4龄和5龄幼虫也分别达57.47%和55.06%,6龄仅25.28%。
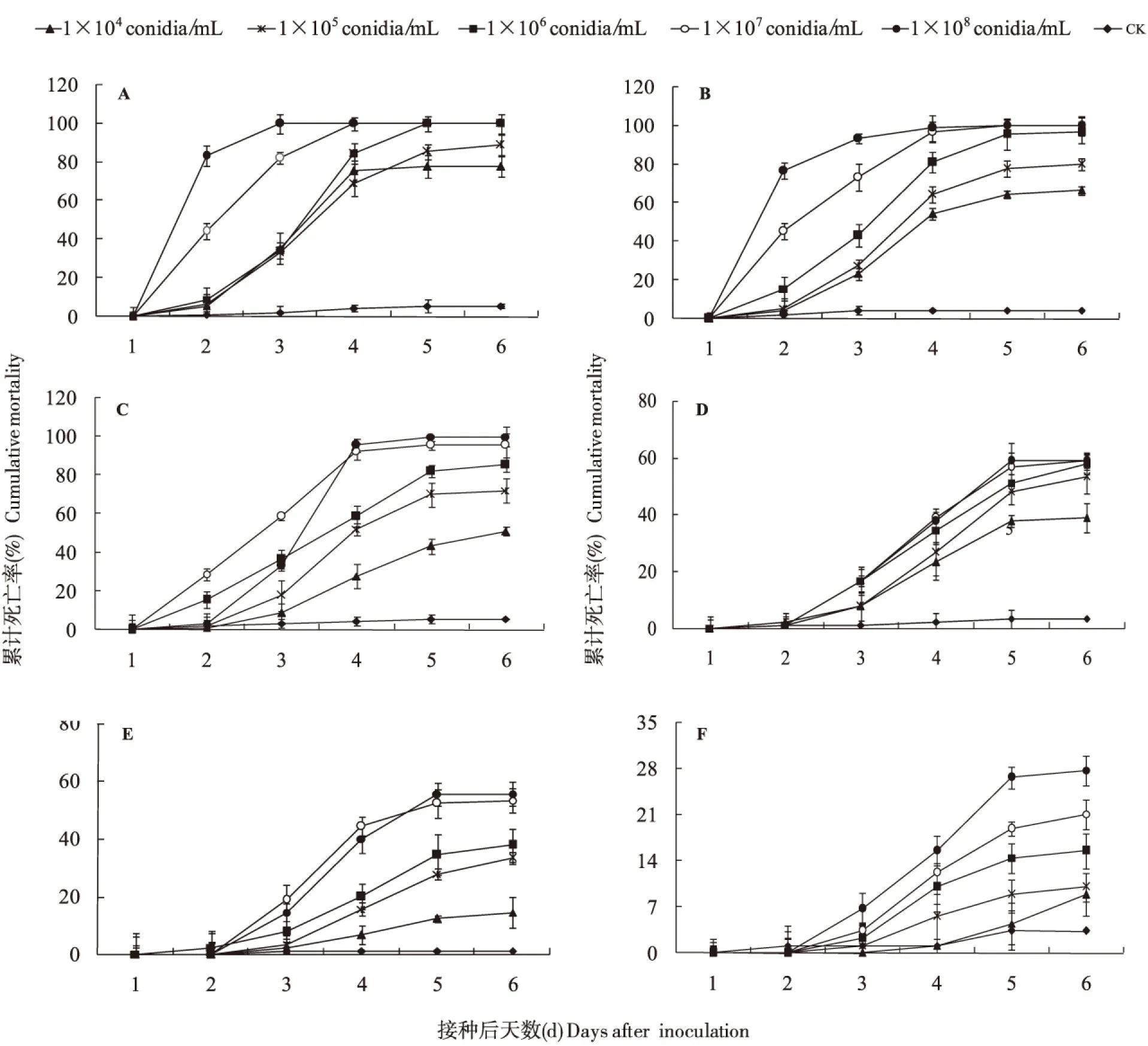
图3 球孢白僵菌GZSL-1对草地贪夜蛾幼虫的致病力Fig.3 The pathogenicity of Beauveria bassiana strain GZSL-1 against Spodoptera frugiperda larvae注:A,1龄;B,2龄;C,3龄;D,4龄;E,5龄;F,6龄。Note: A, 1st instar; B, 2nd instar; C, 3rd instar; D, 4th instar; E, 5th instar; F, 6th instar.
应用Probit模型,计算球孢白僵菌GZSL-1处理的草地贪夜蛾1~5龄幼虫LC50值(表1),其分别为1.32×103、3.42×103、1.01×104、1.61×105和1.23×107孢子/mL。结果表明致中死浓度,1龄和2龄幼虫最低,均为103孢子/mL,3~4龄为104和105孢子/mL,对5龄幼虫的LC50最高,107孢子/mL。草地贪夜蛾1~4龄幼虫对该菌株较敏感,其次5龄,6龄幼虫敏感性最低。
2.4 球孢白僵菌GZSL-1对草地贪夜蛾幼虫的致死时间效应
随球孢白僵菌GZSL-1孢子悬浮液浓度的增加,草地贪夜蛾幼虫的致死中时(LT50)缩短(表2)。孢子浓度越高,幼虫的LT50值越小,在1.0×104~1.0×108孢子/mL范围内,1龄幼虫LT50从3.58 d降至1.69 d,对2龄幼虫的LT50从4.30 d降至1.78 d,对3龄幼虫的LT50从5.70 d降至3.12 d;在浓度为1.0×105~1.0×108孢子/mL的范围内,对4龄幼虫的LT50从5.45 d降至4.85 d;在浓度为1.0×107~1.0×108孢子/mL的范围内,对5龄幼虫的LT50从5.04 d降至5.02 d。当孢子浓度为1.0×104孢子/mL时,4龄幼虫最终死亡率为38.89%,低于50%;当浓度为1.0×104和1.0×106孢子/mL时,5龄的最终死亡率从14.45%升至37.71%,6龄的最终死亡率从8.89%升至15.56%,均低于50%;浓度为1.0×107~1.0×108孢子/mL时,6龄幼虫的最终死亡率从21.11%升至27.80%,亦低于50%,因此无法计算LT50值。
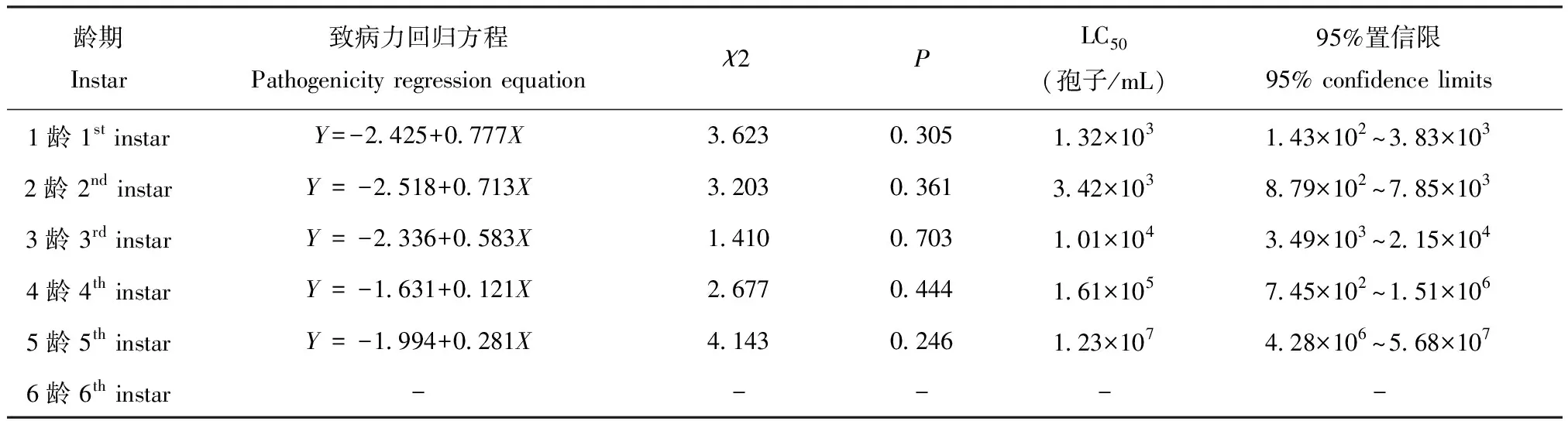
表1 球孢白僵菌GZSL-1对草地贪夜蛾幼虫的致病力回归方程
注:表中数据为平均值±标准差;“-”表示草地贪夜蛾幼虫的最终死亡率低于50%,无法估算LC50。Note: Data in a column are presented as mean±SD. “-” represent that the mortality of infected larvae was less than 50%, LC50values cannot be estimated.
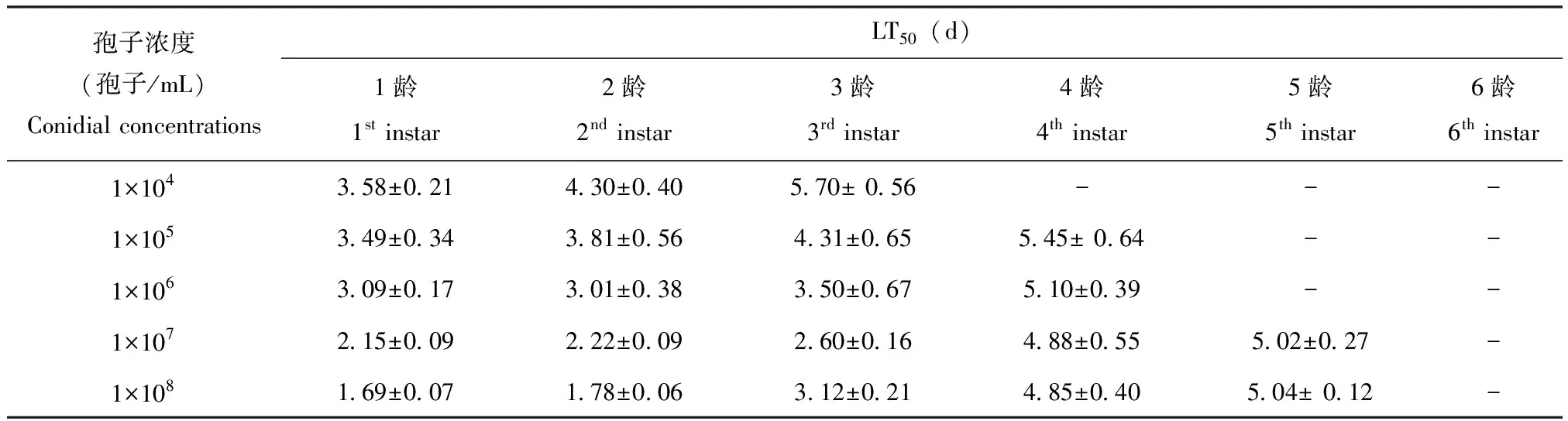
表2 球孢白僵菌GZSL-1对草地贪夜蛾幼虫的致死中时间
注:表中数据为平均值±标准差;“-”表示草地贪夜蛾幼虫的最终死亡率低于50%,无法估算LT50。Note: Data in a column are presented as mean±SD. “-”represent that the mortality of infected larvae was less than 50%, LT50values cannot be estimated.
3 结论与讨论
我国微生物资源丰富,发展生物农药具有充分的优势。为进一步丰富草地贪夜蛾生物防治菌种资源,本研究从采集的感菌稻黑蝽若虫分离获得1株对草地贪夜蛾有高致病性的昆虫病原真菌GZSL-1,根据该菌株培养形态特征及ITS序列比对分析,将其鉴定为球孢白僵菌。菌株GZSL-1对草地贪夜蛾1龄至6龄幼虫皆有侵染效果,但龄期间的致病力差异较大。在同一孢子浓度下,对低龄幼虫的致病力大于高龄幼虫,对高龄幼虫的LC50远大于低龄幼虫的LC50,即使以高浓度孢子悬浮液侵染,对6龄幼虫的致死率仅25.28%(1.0×108孢子/mL),这与以往报道的草地贪夜蛾病原真菌相类似,绝大部分仅对草地贪夜蛾低龄幼虫防效较好(Lezama Gutiérrezetal., 2001; Carneiroetal., 2008; Garcíaetal., 2011; Thomazonietal., 2014; Akutseetal., 2019; 郑亚强等,2019;雷妍圆等,2020)。尽管菌株GZSL-1对4龄和5龄幼虫具有一定程度的致病力,但也仅为50%的致死率,因此利用球孢白僵菌GZSL-1防治草地贪夜蛾幼虫的适期是低龄幼虫种群的高峰期。
筛选昆虫病原真菌高效菌株时,致病力是衡量其防效潜力的重要指标之一。目标害虫死亡率越高且LT50越短,意味着菌株具有更好的致病力。据报道,用35株球孢白僵菌B.bassiana菌株和14株金龟子绿僵菌M.anisopliae对草地贪夜蛾进行致病力筛选,仅有1株球孢白僵菌菌株Unioeste 26可导致草地贪夜蛾3龄幼虫44.5%死亡率(Thomazonietal., 2014)。对分离自非洲的6个球孢白僵菌和14个绿僵菌的杀虫效果进行分析,在1.0×108孢子/mL的孢子浓度下,大部分的绿僵菌菌株对卵、初孵幼虫和2龄幼虫有高于80%的致病力,但仅有一株球孢白僵菌ICIPE 676对2龄幼虫造成30%的死亡率(Akutseetal., 2019)。此外,另有报道1株球孢白僵菌菌株Bb42对草地贪夜蛾2龄幼虫的LC50为5.92×103孢子/mL,且1.0×109孢子/mL时,其2龄幼虫死亡率达96.6%,致死中时LT50为3.6 d(Garcíaetal., 2011)。本研究表明,对于龄期相同的2龄幼虫,供试球孢白僵菌GZSL-1的LC50为3.42×103孢子/mL,略低于前者的5.92×103孢子/mL;当处理孢子悬浮液浓度低于前者,仅为1.0×108孢子/mL时,1~3龄幼虫的累计死亡率皆达100%,较Bb42的致病力强;再以2龄幼虫LT50值比较,当孢子浓度为1.0×108孢子/mL时,本研究仅1.78 d,较前者的致死速度快。表明本研究中供试球孢白僵菌GZSL-1对草地贪夜蛾幼虫具有较高的致死率和较短的致死中时,菌株具有较强的致病力,有进一步研究的价值。
球孢白僵菌作为一种重要的昆虫病原真菌,其制剂的应用开发潜力巨大,但球孢白僵菌菌株具异核现象,从虫尸上分离的野生型菌株多为异核体,由于异核体稳定,人工培养基上继代培养过程中,异核内核比例不断变化,更适应于培养基腐生类型的核被选择出来占优势,这意味着分离出来的菌株在腐生条件下存在变异和退化可能。从菌株分离、毒力保持到利用白僵菌菌剂在自然环境中进行大规模害虫防治,仍然面临着诸多问题。虽然本研究中的球孢白僵菌GZSL-1对草地贪夜蛾幼虫具有较强致病力,这说明在室内条件合适时可起如此大的作用,但在田间应用时是否有同样的致病力,以及如何挖掘菌株本身的潜力去适应外界条件,还有许多工作要做。
猜你喜欢
中华活页文选·教师版(2022年19期)2022-12-31
中国生物防治学报(2022年3期)2022-07-09
家长(2021年20期)2021-12-03
云南农业(2021年11期)2021-11-12
湖南林业科技(2021年3期)2021-07-02
企业改革与管理(2021年10期)2021-01-02
今日农业(2020年13期)2020-12-15
就业与保障(2020年23期)2020-11-24
今日农业(2019年13期)2019-08-12
今日农业(2019年10期)2019-06-26
