
miRNA在动物毛囊发育中的研究进展
2020-07-06 02:48:34刘公言白莉雅李福昌常莹姜文学杨丽萍孙海涛高淑霞
山东农业科学 2020年6期
刘公言,白莉雅,李福昌,常莹,姜文学,杨丽萍,孙海涛,高淑霞
(1.山东省农业科学院畜牧兽医研究所/山东省畜禽疫病防治与繁育重点实验室,山东 济南 250100;2.山东农业大学动物科技学院,山东 泰安 271018;3.山东省畜牧协会,山东 济南 250100)
microRNA(miRNA)是一种长度约20~25个核苷酸的内源性非编码小分子RNA,通过互补区域与靶基因的mRNA结合导致mRNA降解或转录抑制,影响基因表达,进而影响动物的生长发育过程等[1]。
近年来,越来越多的研究结果表明,miRNA作为有效调节因子参与生物发育和疾病发生过程,包括参与动物被毛生长发育、毛囊周期循环和皮肤角质形成细胞分化等生物学过程[2,3]。Yi等[4]在小鼠胚胎期表皮中分离并克隆了大量的miRNA。Andl等[5]通过微矩阵方法对出生小鼠皮肤miRNA的表达谱进行分析,进一步确定了miRNA在哺乳动物皮肤毛囊发育中发挥重要作用。miR-203是第一个被发现的在动物皮肤及毛囊中丰富表达,且与动物皮肤及毛囊发育密切相关的miRNA,被称为“皮肤特异性miRNA”[6]。刘公言[7]研究证实miR-205在高被毛密度獭兔皮肤毛乳头细胞中低表达,在低被毛密度獭兔皮肤毛乳头细胞中高表达,且在獭兔皮肤组织中高表达,具有组织表达特异性。本研究从哺乳动物miRNA生物合成、miRNA在毛囊发育中的生理功能及其调控毛囊发育作用机制等方面进行详细阐述,旨在为深入了解miRNA调控动物毛囊的发育机制,并为今后指导毛用和皮用动物选育和提高其生产性能提供参考。
1 动物体内miRNA的生物合成
miRNA在动物体内的生物合成过程包括miRNA的形成和成熟过程。首先需要编码miRNA基因在RNA聚合酶Ⅱ(RNA polymeraseⅡ,RNA PolⅡ)的作用下转录生成长度约为300~1 000个碱基的初级转录物(pri-miRNA),再经细胞核中Drosha酶剪切产生具有茎环结构且大小约70个核苷酸的miRNA 前体(pre-miRNA)。随后,pre-miRNA在转运蛋白Exportin-5作用下从细胞核内转运到细胞质,并被细胞质中Dicer酶剪切成长度约22个碱基的miRNA/miRNA*。最终miRNA成熟体被装配到由Argonaute蛋白组成的蛋白复合体中,形成具有生物学功能的沉默复合体(RNA-induced silencing complex,RISC),进而引起靶mRNA降解,抑制翻译过程(图1)[8,9]。
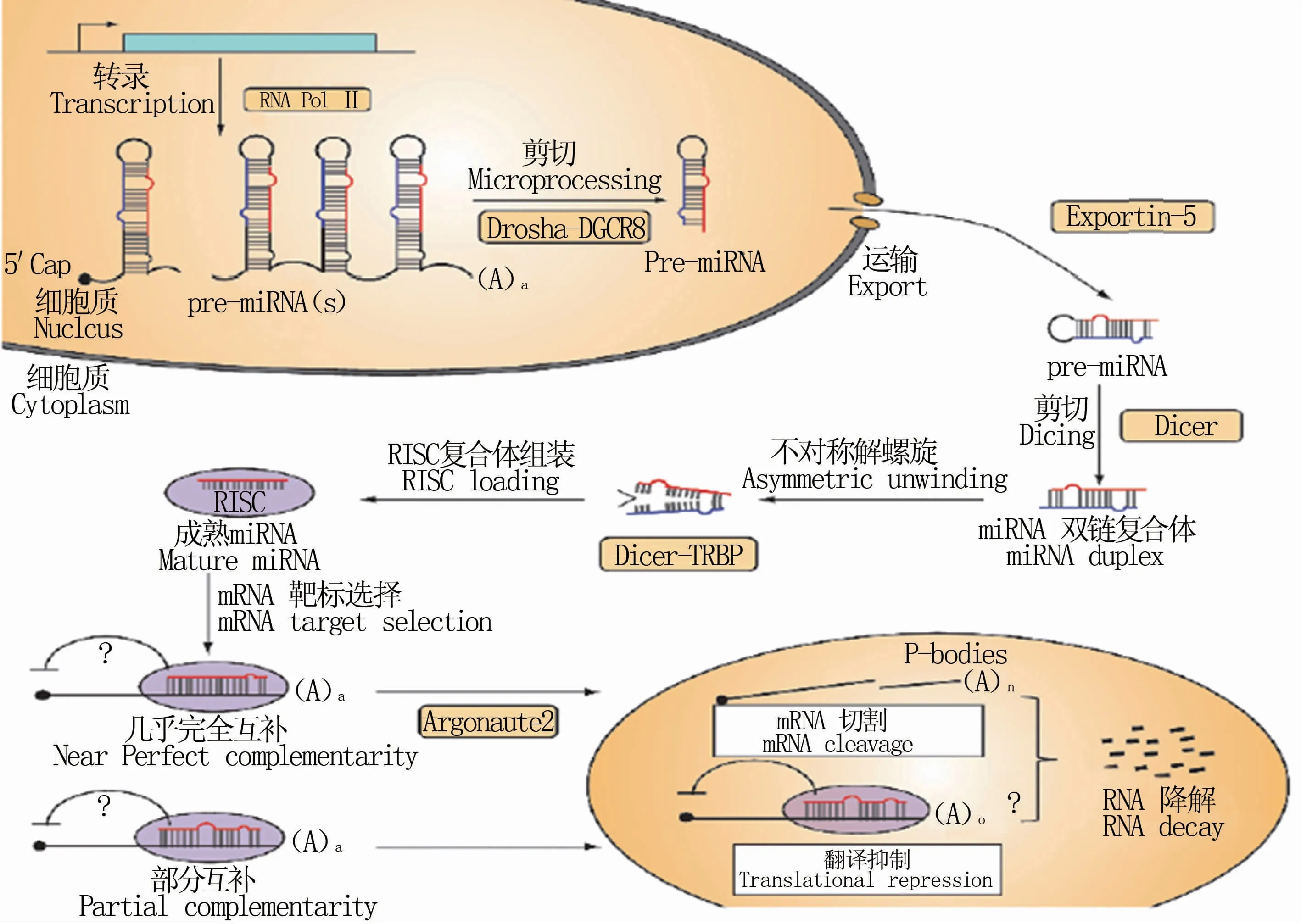
图1 动物体内miRNA的生物合成过程
2 miRNA在动物毛囊发育中的功能
2.1 调节毛囊发育的时序性
唐晓惠[10]利用高通量测序技术分别检测了藏绵羊处于生长期、衰退期和休止期三个时期的毛囊miRNA表达谱,其中毛囊生长期特异表达miRNA的数量为10个,衰退期为27个,休止期为7个,推断这些差异表达miRNA对毛囊生长周期具有重要调控作用[2]。Mardaryev等[2]研究表明,miRNA-31在小鼠毛囊生长期高表达、衰退期低表达,参与毛囊周期性生长过程。miR-205在小鼠毛囊生长期表达量低,但在毛囊衰退期表达量最高,对毛囊由生长期向衰退期的转换具有驱动作用[11]。小鼠皮肤过表达miR-22可抑制毛囊从静止期向下一个生长期转变,促进毛囊从生长期向退行期转变,从而导致小鼠脱毛,但在敲除表达miR-22的基因后,小鼠毛囊从静止期向下一个生长期的转变加快[12]。
2.2 调节毛囊干细胞的增殖分化和凋亡
Liu等[13]认为miRNA在大鼠、小鼠、山羊和绵羊等多种哺乳动物皮肤中均有表达,并在毛囊形态发生、毛囊干细胞增殖分化和凋亡中发挥重要的调节作用。Yi等[14]利用小鼠皮肤组织进行免疫组化发现miR-203在分化的细胞中高表达。Zhang等[15]研究认为miR-125b在小鼠皮肤干细胞中大量表达且随皮肤干细胞分化程度增强而降低,过表达miR-125b可使小鼠皮脂腺变大、表皮变厚不能形成被毛,因此,miR-125b能够抑制小鼠干细胞分化。miR-22可以通过促进小鼠毛囊干细胞凋亡,抑制毛囊干细胞增殖和分化等来促进毛囊退化。Yuan等[12]釆用短期诱导miR-22表达的方法证实miR-22可以直接促进小鼠毛囊干细胞分化过程,而miR-31在小鼠皮肤中干扰表达会导致毛囊干细胞异常增殖和毛囊形态异常[2]。
2.3 调节皮肤被毛颜色的形成
miRNA参与动物皮肤及被毛颜色形成。Yan等[16]研究发现miR-429参与调节鱼类皮肤色素沉着。Wang等[17]研究表明,miR-204/211在小鼠黑色素细胞中通过小眼畸形相关转录因子(microphthalmia-associated transcription factor,MITF)靶向调节色素沉着,并在MITF敲除的黑色素细胞中差异表达。Zhu等[18]认为miR-25在动物皮肤及毛发生长发育过程中发挥重要调控作用,其作用机理是显著下调羊驼黑色素细胞中MITF基因的表达,同时抑制肌球蛋白5a(myosin 5a,MYO5a)和酪氨酸酶(tyrosinase,TYR)的表达。Dong等[3]研究证实miR-137影响小鼠皮肤和被毛颜色,而此前已有研究证明miR-137参与小鼠皮肤和被毛颜色的形成[5],但目前尚无相关数据表明miR-137在人毛囊或皮肤组织中表达。此外,皮肤中的miRNA还参与了动物皮肤对紫外线照射的应答[19]。
2.4 调控多种毛囊疾病的发生
miRNA参与人类多种疾病的发生发展过程[20-23]。Polytarchou等[22]研究发现miR-214表达与结肠癌病程发展密切相关。miR-17-92在结肠癌的发生发展过程中起重要作用,可靶向调控多个诱导肿瘤血管发生相关基因的表达[23]。Xiang等[24]应用miRNA扩增分析了男性秃顶与非秃顶部头皮所分离的毛乳头细胞miRNA的差异表达,发现miR-106a、miR-410、miR-221、miR-125b的表达存在显著差异,它们与人类脱发疾病发生密切相关。
3 miRNA在动物毛囊发育中的作用机制
3.1 通过Wnt信号通路参与毛囊发育调控
Wnt家族是一类分泌型脂质修饰糖蛋白,参与调控动物毛囊发育及周期性生长过程[24,25]。Wnt蛋白配体通过与跨膜蛋白(frizzled,FZD)受体及脂蛋白受体相关蛋白5或6(lipoprotein receptor related protein 5 or 6,LRP5/6)等共受体结合来激活该途径[26]。Wnt与FZD/LRP复合物可以诱导β-连环蛋白(β-catenin)磷酸化,从而使β-catenin和T细胞因子(T-cell factor,TCF)/淋巴增强因子(lymphoid enhancing factor,LEF)/β-catenin介导的基因表达,调节毛囊细胞增殖和分化[27,28]。大肠腺瘤性息肉病(adenomatous polyposis coli,APC)蛋白与β-catenin相互作用,在控制β-catenin蛋白稳定性方面起着关键作用。大量研究表明,miRNA能够通过Wnt信号通路相关基因调控动物皮肤毛囊发育。miR-24通过其靶基因TCF-3参与调控小鼠毛囊形态的发生[29]。miR-214通过Wnt/β-catenin信号在小鼠毛囊发育中发挥作用,其在外根鞘和毛母质细胞中表达丰富,可通过β-catenin基因靶向调控皮肤上皮细胞的分化,抑制角质细胞增殖,进而抑制毛囊生长发育[30]。Mardaryev等[31]对小鼠不同生长时期皮肤中miRNA进行研究,发现miR-31可通过Wnt信号通路和BMP信号通路调控小鼠毛囊发育及被毛生长。Dickkopf相关蛋白1(dickkopf relative protein 1,DKK1)是Wnt信号通路的重要拮抗物,胚胎期过表达DKK1的小鼠不能生成毛囊,而小鼠出生后DKK1过表达可抑制毛囊生长[32]。过表达miR-218能够显著降低Dickkopf相关蛋白2(dickkopf relative protein 2,DKK2)和分泌型FZD相关蛋白(secreted FZD related protein 2,SFRP2)水平,此外,miR-218可激活Wnt/β-catenin信号通路,并通过Wnt3a/DKK2/SFRP2增加β-catenin转录活性[33-35]。miR-29的表达受Wnt途径的影响,可通过降低Wnt拮抗剂DKK1和SFRP2的表达来增强Wnt信号传导[36]。转基因小鼠过表达DKK1后,表皮内miR-200b和miR-196a的表达量会显著减少,表明它们可能存在于Wnt信号通路中[5]。在毛囊发育早期,miR-205可负调控PI3K信号通路抑制因子Inppl1、Inpp4b、Frk、Phlda3等,从而调控毛囊干细胞中Akt磷酸化水平,维持毛囊干细胞正常增殖;PI3K/Akt信号通路可通过磷酸化GSK-3β抑制β-catenin磷酸化,激活Wnt信号通路[11]。
3.2 通过BMP信号通路参与毛囊发育调控
骨形态发生蛋白质(bone morphogenetic protein,BMP)信号通路参与哺乳动物皮肤生长发育过程,是毛囊发育的重要信号通路之一。转化生长因子-β(transforming growth factor-β,TGFβ)可以抑制体外培养的角质形成细胞增殖,参与动物皮肤生物学形成过程。BMP信号通路与TGF-β信号通路密切相关,miRNAs在调节皮肤生物学形成过程中受到TGF-β信号转导的影响。TGF-β1在毛囊生长期向退行期转换过程中表达于内根鞘,具有抑制细胞增殖作用,促使毛囊由生长期过渡到退行期。部分let-7b和miR-24的靶基因与毛囊生长发育和毛发品质相关[37]。Shen等[38]研究得出let-7b通过抑制转化生长因子-β 受体1(transforming growth factor-βreceptor 1,TGF-βR1)的转录,促进羊驼毛生长。Noggin对BMP信号通路具有负调控作用,主要表现为生长期毛囊中BMP信号下调,Noggin表达量增加。进一步研究表明,对缺失Noggin型转基因小鼠给予Noggin蛋白刺激,能够使毛囊由休止期进入生长期[39]。miR-21在正常小鼠表皮细胞和毛囊上皮细胞中均有表达,Ahmed等[40]研究发现BMP4对miR-21起负调控作用。miR-31通过抑制成纤维细胞生长因子10(fibroblast growth factor 10,FGF10)等靶基因的表达调控动物被毛生长。另有研究认为,miR-31可以通过负向调控缺氧诱导因子1(hypoxiainducible factor-1,HIF-1)的表达来激活Notch信号通路,进而影响角质形成细胞的分化[41]。
3.3 通过细胞因子参与毛囊发育调控
胰岛素样生长因子家族中的胰岛素样生长因子1(insulin-like growth factors 1,IGF-1)广泛分布于毛囊中,能够刺激黑色素细胞、上皮细胞和毛乳头细胞等各种毛囊细胞的增殖分化,其作用机理是通过与IGF-1受体结合将信号传导来刺激毛囊细胞的增殖[42]。成纤维细胞生长因子5(fibroblast growth factor 5,FGF5)作为一种重要的生长因子,通过诱导毛囊由生长期向退行期转变来调控动物的被毛长度,在毛囊发育中发挥重要作用。FGF5发生突变后,可以增加人睫毛的生长[43]。miR-let7a在绒山羊生长期中的表达较退行期弱,通过TargetScan软件预测,胰岛素样生长因子1受体(insulin-like growth factors 1 receptor,IGF-1R)mRNA 3′非翻译区上存在miRlet7a的结合位点,且IGF-1R在绒山羊生长期毛囊中的表达量高于退行期,与miR-let7a的表达量正好相反,进一步证实了IGF-1R为miRlet7a的靶基因[44]。ocu-miR-205能够促进獭兔毛乳头细胞凋亡,影响毛乳头细胞和獭兔皮肤内PI3K/Akt、Wnt、Notch和BMP等信号通路中基因和蛋白的表达,促进毛囊由生长期向退行期及休止期转变,影响獭兔被毛密度。
4 存在问题与展望
对于毛用和皮用动物来说,被毛和皮张具有重要的经济价值,而经济价值的高低与毛囊发育密切相关,尤其是毛囊生长期的长短决定毛用动物的产毛量。miRNA作为一类内源性非编码小分子RNA,通过作用于靶基因mRNA的非编码区参与基因转录后调控,进而影响动物毛囊发育的时序性、毛囊干细胞增殖分化以及皮肤被毛颜色等。随着高通量测序技术的成熟和普及,越来越多的与毛囊发育相关的miRNA被挖掘。因此,miRNA在毛囊发育调控过程中的功能和调控机制将成为未来研究工作的重点,且miRNA在其靶基因上靶位点的复杂性和功能的多角色性等也有待后续深入研究。同时,如何通过miRNA来提高毛用动物的产毛性能、皮用动物的皮张质量等解决生产问题以及解决人类脱发等毛囊疾病等,仍然需要科研工作者的不懈努力。
猜你喜欢
军事文摘(2023年2期)2023-02-17 09:20:24
感染、炎症、修复(2021年1期)2021-07-28 06:18:08
学苑创造·A版(2020年12期)2020-01-07 14:07:23
中国外汇(2019年15期)2019-10-14 01:00:34
中华家教(2018年7期)2018-08-01 06:32:38
作文教学研究(2016年1期)2016-07-05 12:22:47
保健与生活(2016年1期)2016-04-12 18:29:44
文学少年(原创儿童文学)(2016年16期)2016-02-28 17:50:19
医学研究杂志(2015年8期)2015-06-22 14:00:57
中国当代医药(2015年7期)2015-03-01 02:01:09
