
田头菇属的系统分类学研究进展*
2020-06-19 10:37刘祈猛刘郁林陈明辉翟志军周健平胡殿明1
中国食用菌 2020年5期
刘祈猛 ,刘郁林,陈明辉,翟志军,周健平,胡殿明1,,3**
(1.江西农业大学食药用菌工程技术研究中心,江西 南昌 330045;2.江西环境工程职业学院,江西 赣州 341002;3.江西农业大学生物科学与工程学院,江西 南昌 330045)
田头菇属(Agrocybe)真菌通常生长于林中地上、草地或粪上,如田头菇Agrocybe praecox(Pers.)Fayod、平田头菇 A.pediades(Fr.)Fayod[1];少数生于腐木上,如茶树菇A.aegerita(V.Brig.)Singer生长于腐烂或死亡的杨树、柳树、茶树等树木上[2];极少数寄生于植物上,如寄生田头菇A.parasitica G.Stev.寄生在新西兰阔叶树上并使寄主死亡[3]。
田头菇属常见可食用种类包括柱状田头菇A.cylindracea(DC.)Maire、杨树菇A.aegerita(V.Brig.)Singer、茶薪菇A.chaxingu N.L.Huang、杨柳田头菇A.salicaceicola Zhu L.Yang,M.Zang & X.X.Liu、田头菇A.praecox(Pers.)Fayod、平田头菇A.pediades(Fr.)Fayod等[4-6]。其中,柱状田头菇与茶薪菇、杨树菇、杨柳田头菇,统称为茶树菇,原为江西广昌境内的高山密林地区茶树蔸部生长的一种野生蕈菌[7]。上世纪80年代,我国开始研究茶树菇的栽培技术,经过30余年的努力,我国茶树菇已经在全国大规模推广栽培,并且形成了巨大的产业[8]。2017年中国茶树菇年产量约90万吨,比2016年产量增加了8.95%,占中国食用菌总产量的2.42%[9]。
茶树菇不但具有高蛋白、低脂肪、低糖等营养特性,而且具有独特的风味物质,鲜食、烘干皆宜[10-11]。此外,茶树菇还有一定的药理作用,有研究表明,其多糖和凝集素具有抗氧化、抗衰老、抗肿瘤和凝集红细胞等功效[12-15]。
多种田头菇属真菌是美味可口的食(药)用菌,但只有少数被成功驯化并人工栽培,其种质资源亟待开发。系统分类学研究是生物资源开发利用的基础。通过对国内外田头菇属的分类研究历史和现状进行综述和展望,可为我国田头菇属的系统分类研究、种质资源开发与利用及其遗传育种研究提供理论参考。
1 田头菇属的形态学分类研究
田头菇属(Agrocybe)属于担子菌门(Basidiomycota)伞菌纲 (Agaricomycetes)伞菌目 (Agaricales)球盖菇科(Strophariaceae)。田头菇属由Fayod于1889年建立,该属的模式种为A.praecox(Pers.)Fayod[16]。田头菇属主要形态特征为菌盖外皮层由球形、梨形或泡囊状细胞组成;菌褶直生至弯生,褶幅较宽,具囊状体,或仅有褶缘囊状体,菌髓平行型至近平行型;菌幕存在或消失,通常具有菌环,少数无;菌柄基部常具白色较细根状菌索;担孢子卵圆形、椭圆形或纺锤形,蜜黄色至浅褐色,光滑,芽孔有或不明显或无孔[1,17]。
自田头菇属建立以来,该属未有统一的分类标准,致使其分类较混乱。直到Singer[17]确定了田头菇属的分类标准,即盖皮层由梨形、近球形、泡囊状的细胞组成;褶缘和褶侧囊状体有无;孢子印锈褐色至黄褐色,孢子芽孔有或不明显等微观特征,并将田头菇属归入粪锈伞科(Bolbitiaceae),这为后续田头菇属的形态分类研究奠定了基础。为了进一步确定其分类地位,将田头菇属与其他相近属形态特征进行比较[1],具体情况见表1。

表1 田头菇属与其他相近属形态特征比较Tab.1 Comparison of morphological characteristics between Agrocybe and other similar genera
如表1所示,田头菇属与粪锈伞科中的一些属的某些形态特征很相似,因此Hawksworth也支持Singer的观点[18]。然而,田头菇属某些种类与球盖菇科(Strophariaceae)的一些属,如鳞伞属 [Pholiota(Fr.)P.Kumm.]和球盖菇属 [Stropharia(Fr.)Quél.],在形态特征上也很相似。于是在很长一段时间里,田头菇属的科级归属地位存有争议。直到Kirk[19]根据分子特征将田头菇属放入球盖菇科中,其分类系统一直沿用至今。当时田头菇属真菌包含100种,而截至目前,Index Fungorum报道的田头菇属真菌约160种[20]。
早期有关田头菇属的分类研究较少,且形态描述主要包括子实体形态特征,而微观结构特征很少被提及。Singer[17]根据孢子有无芽孔或不明显,将田头菇属分为2个亚属,田头菇亚属(Subgen.Agrocybe)和非显孔亚属(Subgen.Aporus)。其中,田头菇亚属又划分为5个组,田头菇组(Sect.Agrocybe)、平田头菇组(Sect.Pediades)、小孢田头菇组(Sect.Microsporae)、全囊体田头菇组(Sect.Allocystides)和幕出田头菇组(Sect.Evelatae);非显孔亚属则包括非显孔田头菇组(Sect.Aporus)和具幕田头菇组(Sect.Velatae)。Watling[21]对田头菇属进行详细的分类学研究,并对Singers分类系统进行了修订,在2个亚属的基础上,将田头菇亚属下的幕出田头菇组(Sect.Evelatae)放入非显孔亚属中,同时删减了该亚属中的全囊体田头菇组(Sect.Allocystides)。此分类系统也一直被后来学者沿用至今,本郷次雄[22]、金鑫[1]等都采用Watling的分类方法对田头菇属真菌进行了形态学系统分类并鉴定。
在Fayod建立田头菇属之初,该属包括9种,其中田头菇A.praecox(Pers.)Fayod为模式种[20]。但由于该属形态特征较原始定义更为宽泛,后续不断有新的物种被纳入田头菇属。如:Singer增加田头菇属真菌56种,其中新种或变种17种:A.aegerita var.rugosovenosa、A.allocystis、A.apepla、A.coprophila、A.firma var.tucumana、A.insignis、A.karelica、A.lazoi、A.microspora、A.neocoprophila、A.paradoxa、A.procera、A.procera f.procera、A.procera var.procera、A.tuberosa var.heterospora、A.viscosa和A.xerophytica[23-26],极大的丰富了该属的种类,其中一个坚实田头菇新变种 A.firma var.tucumana,后经Watling修订为A.tucumana(Singer)Watling[27];Pegler[28-30]增添了该属真菌8种,其中4个新种和1个变种:A.manihotis、A.peradenica、A.stercoraria 和 A.manihotis var.manihotis,Pegler[31]把田头菇属中的 A.cubensis(Murrill)Singer和A.underwoodii(Murrill)Singer分别放入滑诱伞属(Hebeloma P.Kumm.)和脆锈伞属[Naucoria(Fr.)P.Kumm.]中,把Pholiotina musae Earle放入田头菇属;随后,Watling又扩充田头菇属真菌31种,其中新种或变种15种[32-33]。Watling[21]列出了英国田头菇属真菌种类17种,并进行了形态学描述,编制了检索表。Watling和Bigelow[34]对该属进行研究,发表了北美东部的2个新种:Agrocybe smithii Watling & H.E.Bigelow和A.occidentalis Watling & H.E.Bigelow。1986年,Watling和Abraham[35]报道了印度田头菇属真菌17种,其中2个新种:A.aporata Watling & S.P.Abraham和A.indica Watling & S.P.Abraham。随后在该地区又有一些学者对该属做过相继报道[36-38],Natarajan和 Purush[39]发表了 1个新种:A.musicola Natarajan & Purush.;Thoma 和 Manim[40]报道了该属4个新种:A.calicutensis K.A.Thomas & Manim.、A.guruvayoorensis K.A.Thomas & Manim.、A.munnarensis K.A.Thomas & Manim.和 A.wayanadensis K.A.Thomas & Manim.以及2个新记录种。Noordeloos[41]对荷兰田头菇属真菌进行了详细的研究,报道了该国该属13种和2个变种。Niveiro[42]对阿根廷粪锈伞科的整理中,列出了田头菇属名录23种。Coimbra[43]整理了中美洲和南美洲球盖菇科真菌,该属曾报道有29种。此外,许多学者对田头菇属做了零星报道,Stevenson[3]报道了唯一1株寄生在新西兰阔叶树上并使寄主致死的田头菇属新种A.parasitica G.Stev.;Nauta[44]报道了荷兰田头菇属1个新种A.rivulosa Nauta。
综上所述,经Fayod、Singer、Watling、Kirk等其他国外真菌学者的研究,不仅丰富了田头菇属的种类,最终还确定田头菇属归属于球盖菇科(Strophariaceae)。该属包括2亚属和5组,即田头菇亚属(Subgen.Agrocybe),以及田头菇组(Sect.Agrocybe)、平田头菇组(Sect.pediades)、小孢田头菇组(Sect.Microsporae)、非显孔亚属(Subgen.A-porus)、非显孔田头菇组(Sect.Aporus)、具幕田头菇组 (Sect.Velatae)。
2 田头菇属的分子系统学研究
2.1 线粒体基因
线粒体核糖体小亚基(mitochondrial small-subunit,mtSSU)序列的核苷进化率要比核基因组核糖体小亚基(SSU rDNA)高16倍,但比核糖体内部转录间隔区(internal transcribed spacer,ITS)进化率低,非常适用于蕈菌的发育演化关系研究[45]。Gonzalez[46]首次对茶树菇A.aegerita、硬田头菇A.dura、茶薪菇A.chaxingu、湿粘田头菇A.erebia、坚实田头菇A.firma、田头菇A.praecox、沼生田头菇A.paludosa、平田头菇 A.pediades、维瓦田头菇 A.vervacti、A.alnetorum等10种田头菇属真菌采用mtSSU可变区域V4、V6、V9序列进行分子系统学分类研究,成功将10种田头菇属分成4个进化分支。随后,Uhart[47]采用同样的方法把茶树菇复合群A.aegerita complex分成3个分支:欧洲分支、阿根廷分支和远支亚-美分支,发现在亚-美分支菌株之间存在大量遗传交换现象。Callac[48]结合mtSSU序列和线粒体细胞色素b基因(mitochondrial cytochrome b gene,mtcob)对亚洲茶薪菇A.chaxingu和欧洲柱状田头菇A.cylindracea进行了分析,发现这两者间具有高的差异性和可杂交性,认为前者可能是后者的1个变种;陈卫民[49]基于mtSSU序列对云南茶树菇复合群多样性进行系统发育分析,认为杨柳田头菇可从茶树菇复合群作为1个分类单元独立出来,证实茶树菇复合群具有高度的多样性,可能存在新物种。由此得知,mtSSU序列能够很好地区分田头菇属不同物种之间的亲缘关系。
除mtSSU序列外,线粒体核糖体大亚基(mitochondrial large-subunit,mtLSU)[50]、A.aegerita 线粒体基因组中编码家族B DNA聚合酶的基因Aa-PolB[51]、线粒体中间肽酶基因[52]等线粒体基因也将是田头菇属遗传研究热点。
2.2 核糖体DNA基因(rDNA)
真菌rDNA不同区段的保守性不同,于是不同片段系统学分类水平也不同[53]。其中ITS和LSU广泛应用于真菌分类鉴定,田头菇属系统学研究的rDNA基因片段主要有LSU序列和ITS序列,如Petersen[54]采用LSU基因对湿粘田头菇A.erebia和田头菇A.praecox系统发育分析,结果表明,2种田头菇属真菌均分布于球盖菇科分支。田恩静[55]则采用ITS序列对中国球盖菇科10个属进行分子系统学研究,发现田头菇属茶薪菇A.chaxingu与球盖菇属、鳞伞属等分布在同一支,这与Kirk观点一致。Cheong[56]分析了30株田头菇属菌株ITS序列的差异,将其分成8个组。Malysheva[57]基于ITS序列构建系统发育树并分析了俄罗斯平田头菇复合群34种,发现大多数物种属于平田头菇变种A.pediades var.pediades,少数为新变种A pediades var.bispora。Malysheva[58]基于多基因LSU+ITS序列分析,将Galera besseyi Peck 和 Bolbitius liberatus Kalchbr.分别修订为A.besseyi(Peck)E.F.Malysheva,G.Moreno & M.Villarreal和A.liberate(Kalchbr.)E.F.Malysheva。Vizzini[59]分别采用LSU和ITS序列分析,将田头菇属分成4组,并把非显孔亚属的茶树菇复合群、湿粘田头菇Cyclocybe erebia和意大利新种C.erebioides归并入环伞属Cyclocybe。Callac[48]、陈卫民[49]、Alam[60]等对田头菇属遗传多样性的研究中均用到ITS序列。
此外,一些其他基因比如茶树菇子实体分化基因Aa-Pri1、Aa-Pri2、Aa-Pri3[61-63],以及信息素受体基因[64]也有相关田头菇属真菌的报道。
3 田头菇属的其他分类方法研究
传统形态学和分子系统学是田头菇属分类研究的主要方法,除此之外,一些生理生化特征也可以作为分类依据。根据生物学物种的定义,具有交配亲和性或无生殖隔离的类群可视为同一物种,因此,这一特点可作为物种鉴定——交配试验[65]。
Flynn[66]对北美和欧洲田头菇复合群Agrocybe praecox complex进行交配相容性试验,认为A.molesta分类群应放在田头菇复合群A.praecox complex。Uhart[67]通过交配试验在柱状田头菇分类单元发现1新种,即 A.wrightii Uhart & Albertó。Callac[48]通过交配试验证明亚洲茶薪菇和欧洲柱状田头菇可杂交,说明这两个菌株有一定的亲和性,认为茶薪菇是柱状田头菇的1个变种。陈卫民[49]基于ITS和mtSSU序列并结合交配试验,从而证明杨柳田头菇可作为单独的物种从茶树菇复合群独立出来。
4 中国田头菇属真菌资源研究
田头菇属的研究主要集中在欧美和印度地区,我国对田头菇属的研究较少,仅有金鑫[1]对我国田头菇属真菌资源进行过系统研究,其他都是零星报道。
茶薪菇Agrocybe chaxingu N.L.Huang,是我国黄年来于1970年在福建油茶上发现并命名的1种新田头菇属真菌,该命名存在争议[20,68]。1993年,杨祝良[4]发表了无孔组的1个滇产新种:杨柳田头菇A.salicaceicola Zhu L.Yang,M.Zang & X.X.Liu;卢成英[69]报道了湖南吉首和张家界国家森林公园中中国田头菇属1个新记录种无环田头菇 A.fariancea Hongo。金鑫[70]报道了田头菇属3个中国新记录种:隆起田头菇A.elatella(P.Karst.)Vesterh.、褐色田头菇A.brunneola(Fr.)Bon和平田头菇环状变种A.pediades var.cinctula Nauta。同时对中国田头菇属进行系统描述,增加了2个中国新记录种,即平田头菇粪生变种A.pediades var.fimicola(Speg.)Nauta和坚壳田头菇A.putaminum(Maire)Singer及5个省级新记录,共鉴定该属真菌21种[1]。秦改娟[71]报道了采自北京延庆的中国田头菇属1个新纪录种菌核田头菇A.tuberose(Henn.)Singer;王锋尖[72]在湖北省赛武当国家级自然保护区发现中国田头菇属1个新记录种坚实田头菇A.firma(Peck)Singer。
关于我国田头菇属真菌的遗传多样性有较多报道,谭琦[73]利用随机扩增多态性(random amplified polymorphic DNA,RAPD)技术对不同地理环境下柳松菇菌株遗传差异进行测定,发现源于国外的8个菌株与我国6株野生菌株遗传差异明显。鲍大鹏[74]在谭琦研究的基础上增加了核糖体DNA扩增片段限制性内切酶分析(amplifed ribosomal DNA restriction analysis,ARDRA)技术,对7株柳松菇菌株进行遗传多样性分析,发现2株希腊菌株和5株中国菌株差异明显。何莹莹[75]和姜性坚[76]分别对云南2种茶树菇和湖南地区8株柱状田头菇栽培品种进行了遗传多样性的扩增片段长度多态性(amplified fragment length polymorphisms,AFLP)分析,通过聚类得出,前者将茶树菇和杨柳田头菇聚为2类,得到了很好的区分,后者能清晰地显示出不同品种的多态性差异,将8株分为3支。王筱凡[7]对茶树菇进行表达序列标签简单重复序列(expressed squence tags-simple sequence repeats,EST-SSR) 引物开发及通用性研究,挑选的28对引物多样性好且稳定的有13对,其PCR产物的多态性可成功将17株茶树菇菌株进行分类,从而说明菌株间具有一定的遗传差异性。
我国幅源辽阔,田头菇属的物种资源丰富,且其生长宿主具有多样性。根据上述文献报道,我国已报道田头菇属大约有21个,约占全世界的13%,其中包括同物异名和变种,见表2。
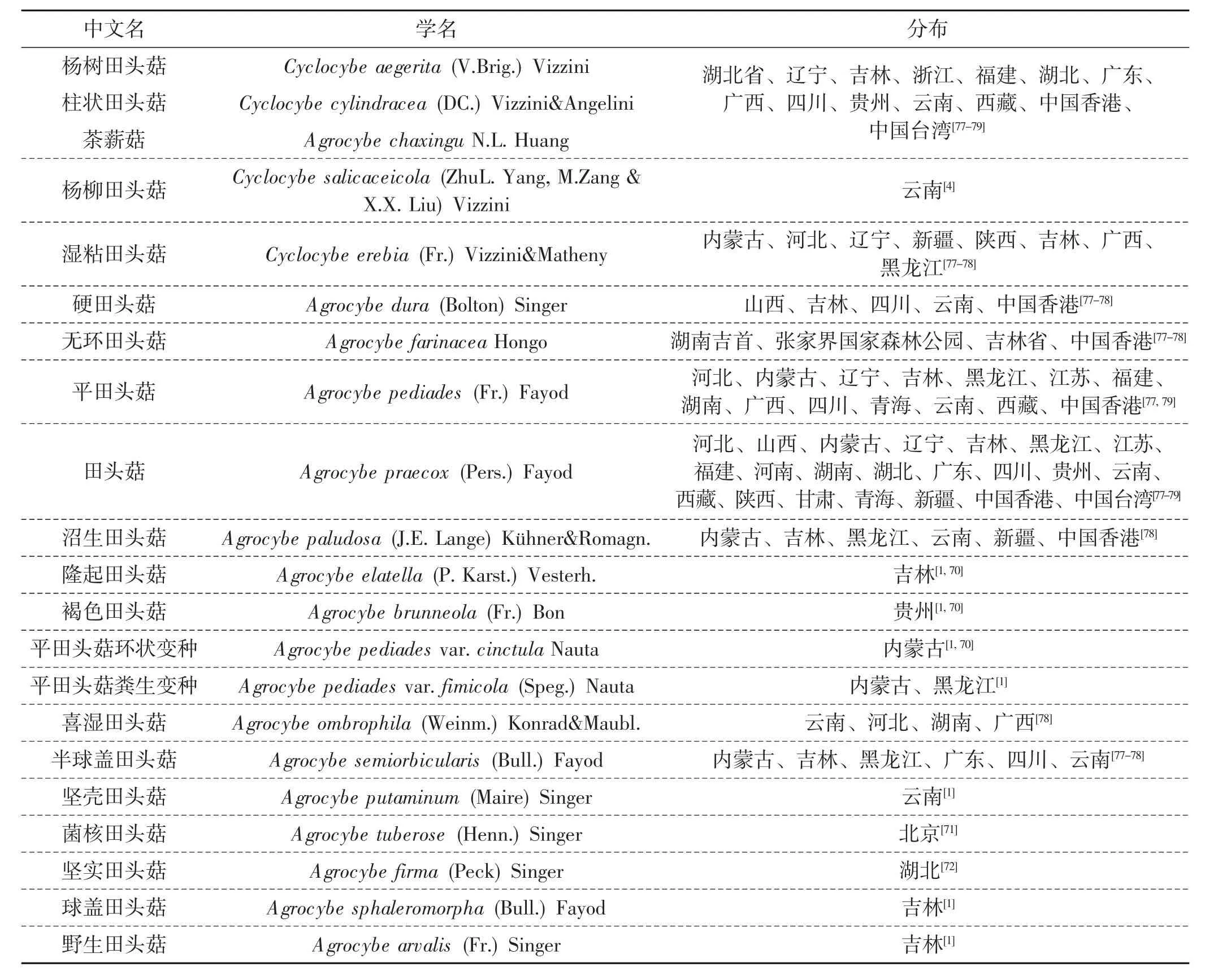
表2 中国田头菇属种类及分布Tab.2 Species and distribution of the genus Agrocybe in china
5 小结与展望
长期以来,田头菇属的分类一直较为混乱,没有统一的分类标准,国内外对该属分类研究主要包括传统形态学、分子系统学、种间隔离-交配试验。
形态学研究主要以有无菌环、有无褶缘和褶侧囊状体、孢子大小以及芽孔有无或不明显等特征作为分类依据,易与相近属鳞伞属、裸盖菇属和球盖菇属等混淆。但仅通过传统形态特征分类,无法准确反映不同物种的亲缘关系和进化路线。
现代分子系统学分类已成为真菌物种分类研究的强有力手段,涌现出多种分子标记技术。现有田头菇属的分子系统学研究主要以单基因或2个基因片段的系统发育研究为主,但并不能全面体现其系统演化关系。真菌多基因片段系统发育树的构建被广大学者所采用,其更有利于疑难种或近似种的正确界定。但是田头菇属的多基因分子系统发育学研究还没有报道,因此,多基因分子系统发育研究是田头菇属分子系统分类的研究热点之一。由于地理因素引起的环境变化,导致田头菇属在形态和生理生化特征上的不稳定性,容易发生变化,此时应通过交配试验来验证它们是否产生生殖隔离。
综上所述,未来在田头菇属分类研究,尤其是由于地理因素造成难以区分的真菌中,应该从形态学、分子系统学和交配试验3个层面进行研究。
猜你喜欢
北京大学学报(自然科学版)(2022年4期)2022-08-18
今日农业(2021年3期)2021-12-05
风流一代·青春(2021年10期)2021-10-27
学苑创造·A版(2021年9期)2021-09-16
科学导报(2020年50期)2020-09-09
当代陕西(2018年24期)2019-01-21
东坡赤壁诗词(2018年1期)2018-03-31
烹调知识(2017年11期)2017-11-06
故事会(2017年20期)2017-10-19
读者·原创版(2014年12期)2014-01-16
