
小叶杨不同种源一年生扦插苗速生期表型性状变异定量分析
2020-06-10 00:48葛会硕宋跃朋苏雪辉张德强张晓宇
浙江林业科技 2020年1期
葛会硕,宋跃朋,苏雪辉,张德强,张晓宇
(1.北京林业大学 理学院,北京 100083;2.北京林业大学 生物科学与技术学院,北京 100083;3.焦作市农林科学研究院,河南 焦作 454001)
小叶杨Populus simonii具有控制土壤荒漠化、减少土壤侵蚀和稳定沙丘的优势[1]。同时,小叶杨还具有较高的经济和药用价值[2]。已有学者对小叶杨的生长特性,包括节间长度变量之间的关系和表型变异进行分析[3]。节间长度是幼苗生长的重要指标,节间长度变量间的关系对育种遗传具有重要意义。2016 年,Yamamoto 等[4]基于图像分析的机器学习方法探究了番茄Lycopersicon esculentum幼苗节间长度估计,能够检测到3 个番茄幼苗的时间序列图像中358 个节点的72%。2017 年,李慧敏等[5]研究了玉蜀黍Zea mays穗下节间长度之间的关系,发现了两个与产量和抗倒伏性相关的农艺性状。2018 年,王健等[6]探讨了普通小麦Triticum aestivum节间长度与株高的关系,结果表明矮秆小麦是通过缩短各节间长度来降低株高的。
在小叶杨的生长过程中,由于地理位置和生长环境的不同,小叶杨幼苗样本可能具有不同的表型特征。沈德胜等[7]利用K-means 聚类方法提取了林木冠层孔隙度。考虑到K-means 聚类对初始聚类中心敏感和易陷入局部最优解,Wan 等[8]在K-means 聚类算法中引入了多种基于群体智能的优化算法,发现人工蜂群算法(ABC)的搜索能力与粒子群算法相当,前者收敛速度更快。所以采用基于人工蜂群算法的K-means 聚类(KABC)[9]可以有效防止陷入局部最优,进而达到准确分类的效果。
由于表型变异是复杂遗传多样性的外在表现,所以是遗传多样性的重要研究领域之一[10]。陈美熹等[11]以马尾松Pinus massoniana58 年生采种母树林为研究材料发现母树间在球果表型及种子性状均存在极显著差异。2010 年卫尊征等[12]以东北和华北地区5 个种源的小叶杨为材料,分别对17 个表型及生理性状进行了比较分析,发现小叶杨各性状在种源间和种源内均存在广泛的遗传变异。2014 年,Du 等[3]研究了毛白杨Populus tomentosa的自然表型变异,发现各性状在个体间存在着较大范围的表型变异。
国内外学者对节间长度的研究主要在农作物方面,而林业上幼苗期的树木节间长度直接反映了幼苗的生长状况。此外,扩大种源范围对研究小叶杨种源间和种源内遗传变异具有重要意义。本文以小叶杨幼苗为研究材料,在2017 年4 月29 日至6 月17 日,对来自15 个种源的与生长、叶片和节间长度有关的性状进行了相关性分析和遗传变异分析,为幼苗的生长研究提供参考。
1 材料和方法
1.1 试验材料
2017 年,在北京林业大学科技温室(40°0' N,116°20' E)共测量并记录了77 株产自我国15 个种源地的小叶杨一年生扦插苗个体。种源地自然概况见表1。测定时间为小叶杨全生长季(从2017 年4 月29 日至6 月17日),使用同样生理年龄的插穗进行无性系扩繁,每个系号4 次重复。即每隔15~ 20 d 利用直尺测量、记录一次样本生长的表型性状,包括株高、叶片数、节数和节间长度,共测量和记录4 次,其中节间长度是从地径部分开始从下往上用直尺测量节与节之间长度。
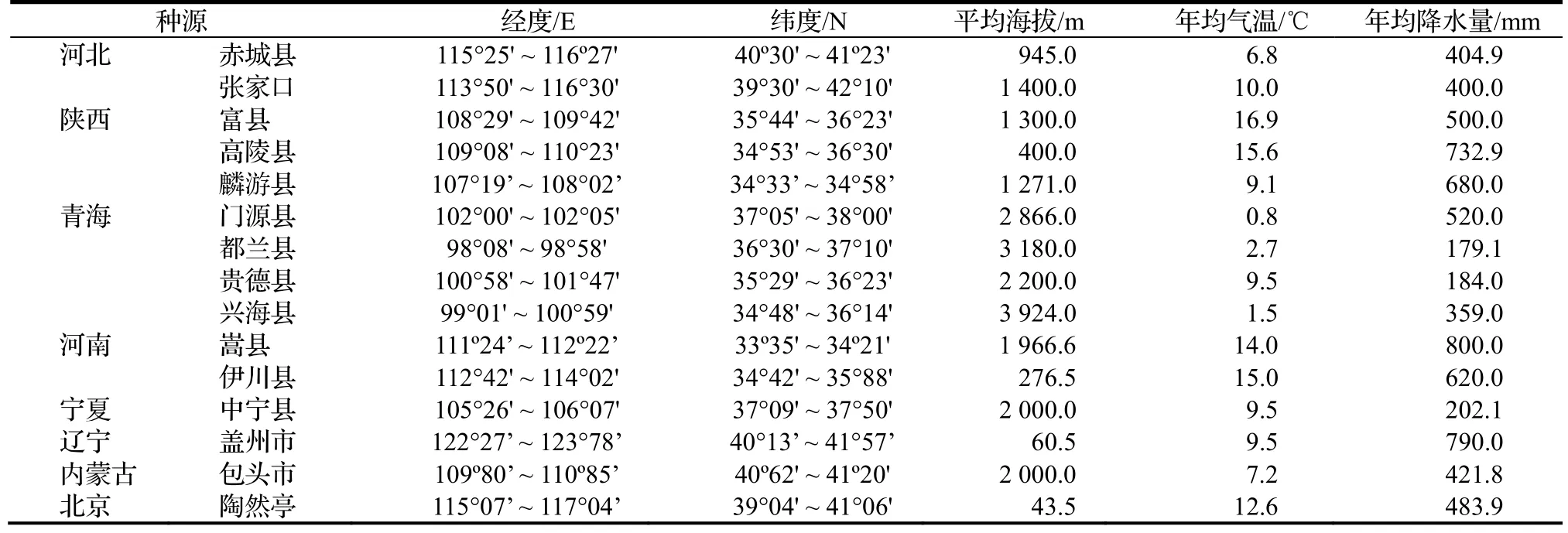
表1 小叶杨种源地的地理位置与气候条件Table 1 Location and climatic conditions of different provenances of P.simonii
1.2 数据处理与统计分析
为了分析小叶杨幼苗节间长度之间的相关性和减少计算复杂度,借助R 语言软件进行因子分析,提取代表大部分特征的公因子(旋转后的因子分析其累积方差贡献率达到65%以上即可)。在进行因子分析之前,首先通过KMO(Kaiser-Meyer-Olkin)检验[13]是否适合做因子分析。KMO 检验值在[0,1]区间。定义如下:

KMO 检验值可以通过R 软件中的psych 包(1.8.4 版)[14]进行计算。
其次,基于Matlab 2016b 软件,将3 个表型特征(株高、叶片数和节数)和因子分析得到的公因子作为输入值代入KABC 进行聚类分析。参数设定:根据R 软件MClust 包中的14 个指标,确定最佳聚类数 。小叶杨的样本集X=(X1,X2,……XN),N(样本量)分别等于80,78,79 和77(4 个时期)。Xi(i=1,2,……N)表示1*7 的向量(株高、叶片数、节数和4 个公共节间长因子)。种源大小(SN)为20,最大迭代次数为100,同一食物源(聚类中心)重复使用次数为10[9]。经过人工蜂群算法的一次聚类中心的优化结合K-means 聚类算法的一次聚类,交互进行,直到算法收敛或达到最大迭代次数即输出聚类中心(K×7)和聚类结果。通过聚类结果和聚类图可以将不同地区的样本进行明确分类,以分析样本和地区间的关系。
最后利用SPSS 21.0 软件对小叶杨幼苗进行统计检验以分析其变异性,试验测定其表型性状的平均值、标准差、变异系数CV和表型分化系数(Vst),以方差的百分比表示群体间的成分,并解释了群体间和群体内表型数据的分化程度。CV和Vst计算如下:
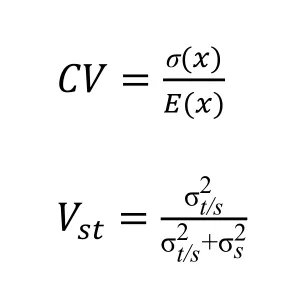
式中,σ(x)代表种源间样本标准差,E(x)代表种源间样本均值,代表种源间的方差分量,代表种源内的方差分量[15]。
2 结果与分析
2.1 小叶杨幼苗节间长度性状定量分析
经计算,4 个时期(4 月29 日、5 月20 日、6 月4 日、6 月17 日)的KMO 值均属于[0.7,0.8]区间,说明样本数据适合做因子分析。分析计算公因子个数,结果如图1。由图1 表明,主成分分析(PCA)建议提取四个公因子;因子分析(EFA)建议提取三个公因子。从EFA 的结果来看,前四个特征值(三角形)在Scree 测试中高于拐点,因此,为了最大限度地保留原始数据信息,决定提取四个公因子(表2)。
由表2 可知,4 个时期的累计贡献率为69.07%~76.69%,所以对应的4 个公因子代表了原始数据的大部分信息。4 个时期因子分析载荷值明显分为4 个区域,D 和C 部分的方差贡献率在4 个时期均比B 和A部分高,说明D 和C 部分的幼苗生长速度较快。且每一个公因子都由相邻节长变量组成(载荷较大),表明相邻节长度具有较强的相似性。
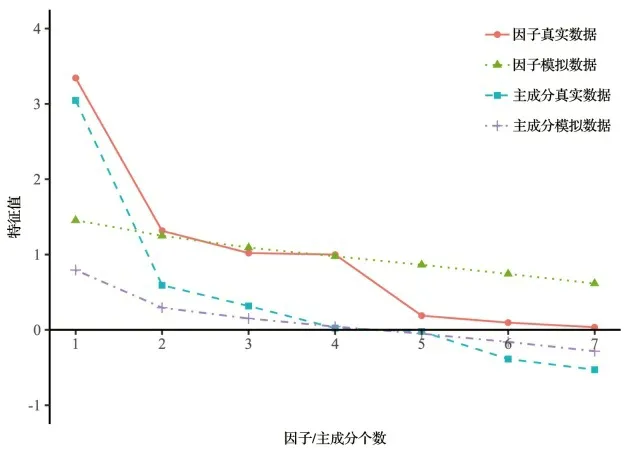
图1 评估因素数量Figure 1 Number of assessment factors
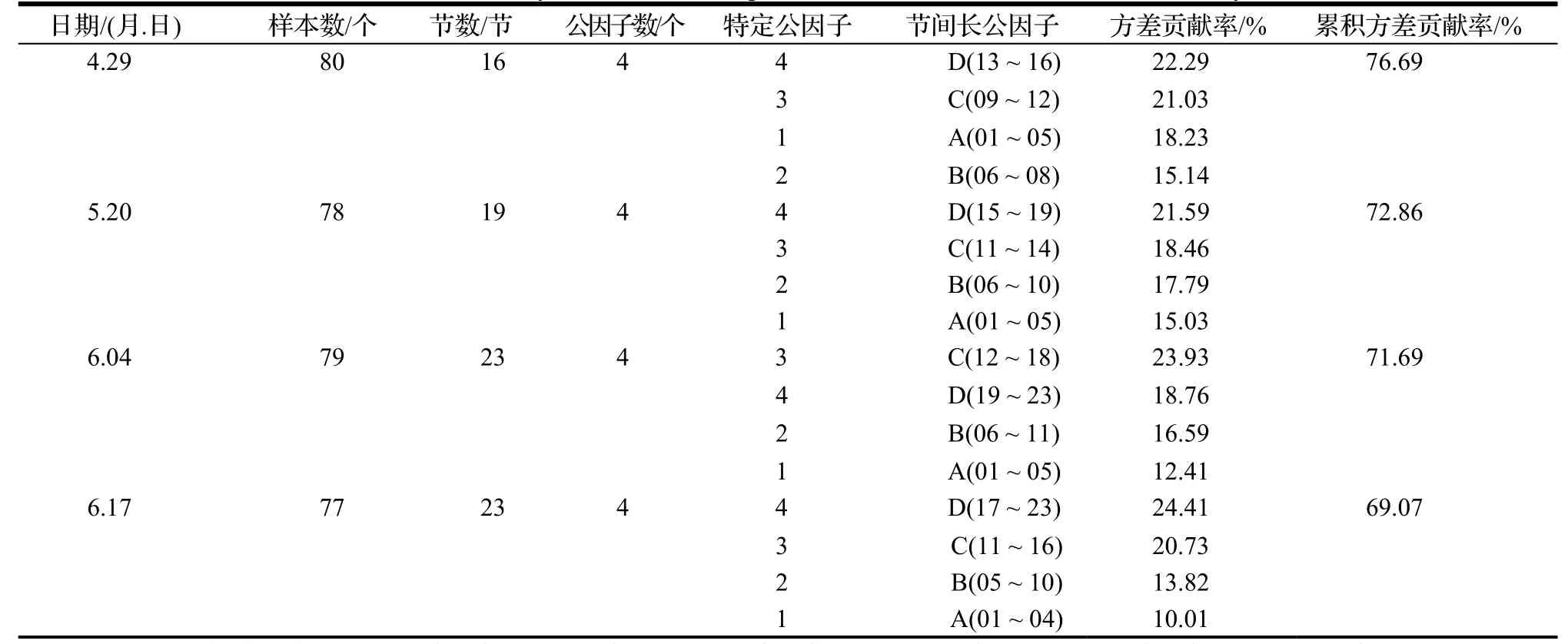
表2 小叶杨4 个时期节间长度变量的因子分析Table 2 Factor analysis of internode length variables of P.simonii at 4 measurement days
2.2 小叶杨幼苗表型性状聚类分析
首先利用R 软件中mclust 包的14 个索引获得推荐聚类结果,结果显示BIC 值在4 个时期中值均为2。因此,确定最佳聚类数为2,如图2。然后利用KABC 算法进行迭代计算,最终得到了4 个时期内小叶杨幼苗样本的聚类结果,如图3。
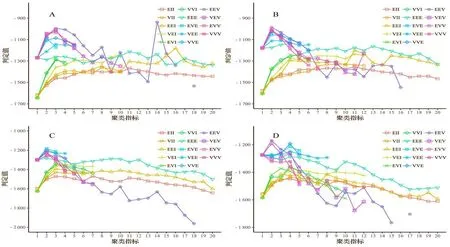
图2 4 个日期的小叶杨幼苗推荐聚类数Figure 2 Recommended cluster number of P.simonii seedlings at 4 measurement date
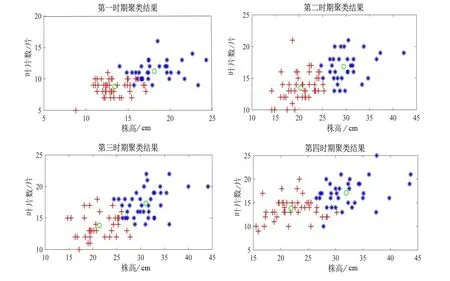
图3 KABC 算法对4 个时期小叶杨幼苗的分类结果Figure 3 Classification maps by KABC algorithms for P.simonii seedlings at 4 measurement dates
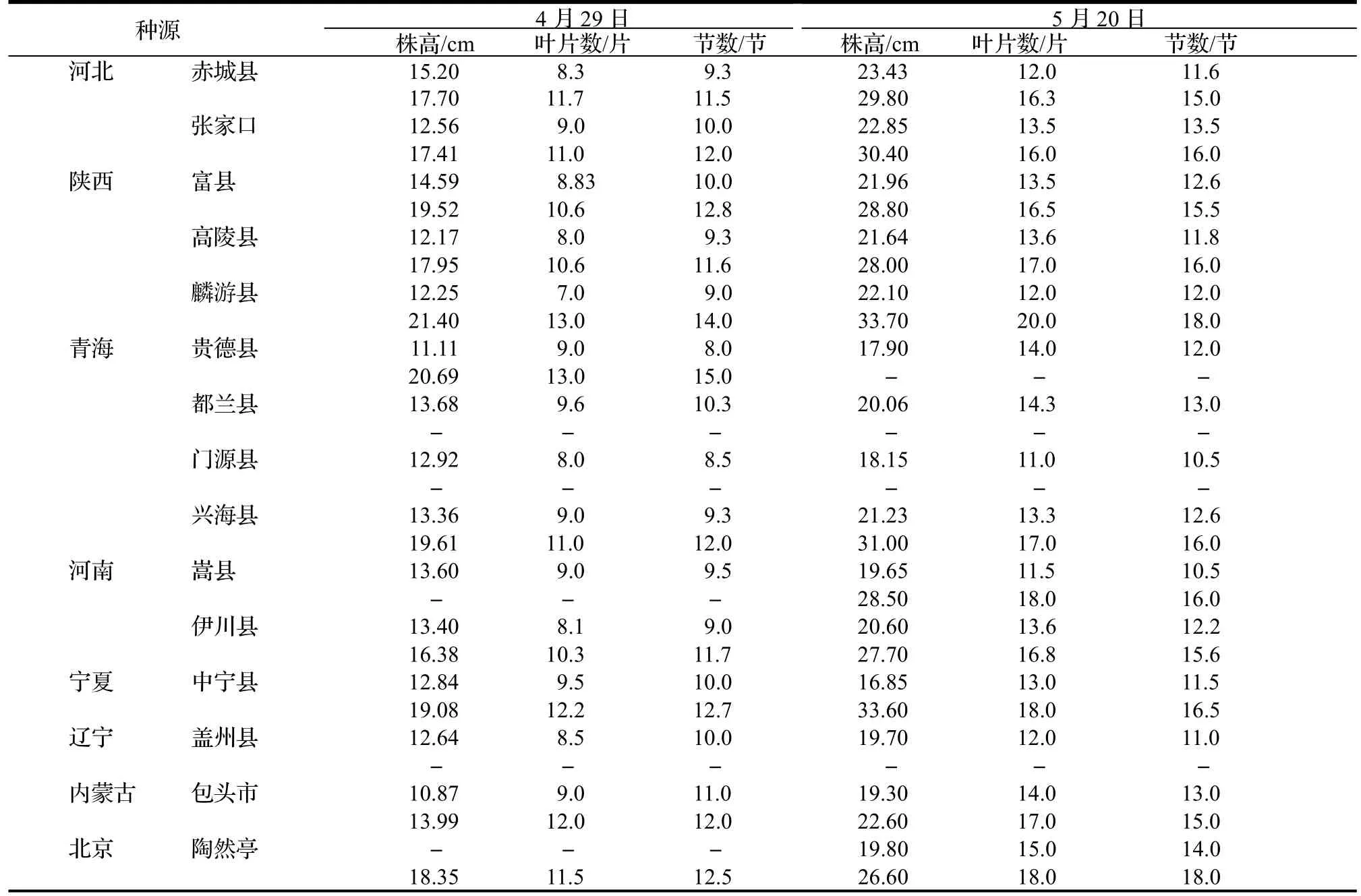
表3 4 月29 日和5 月20 日小叶杨幼苗平均株高、叶片数和节数Table 3 Average height,number of leaf and internode of P.simonii seedlings on April 29th and May 20th
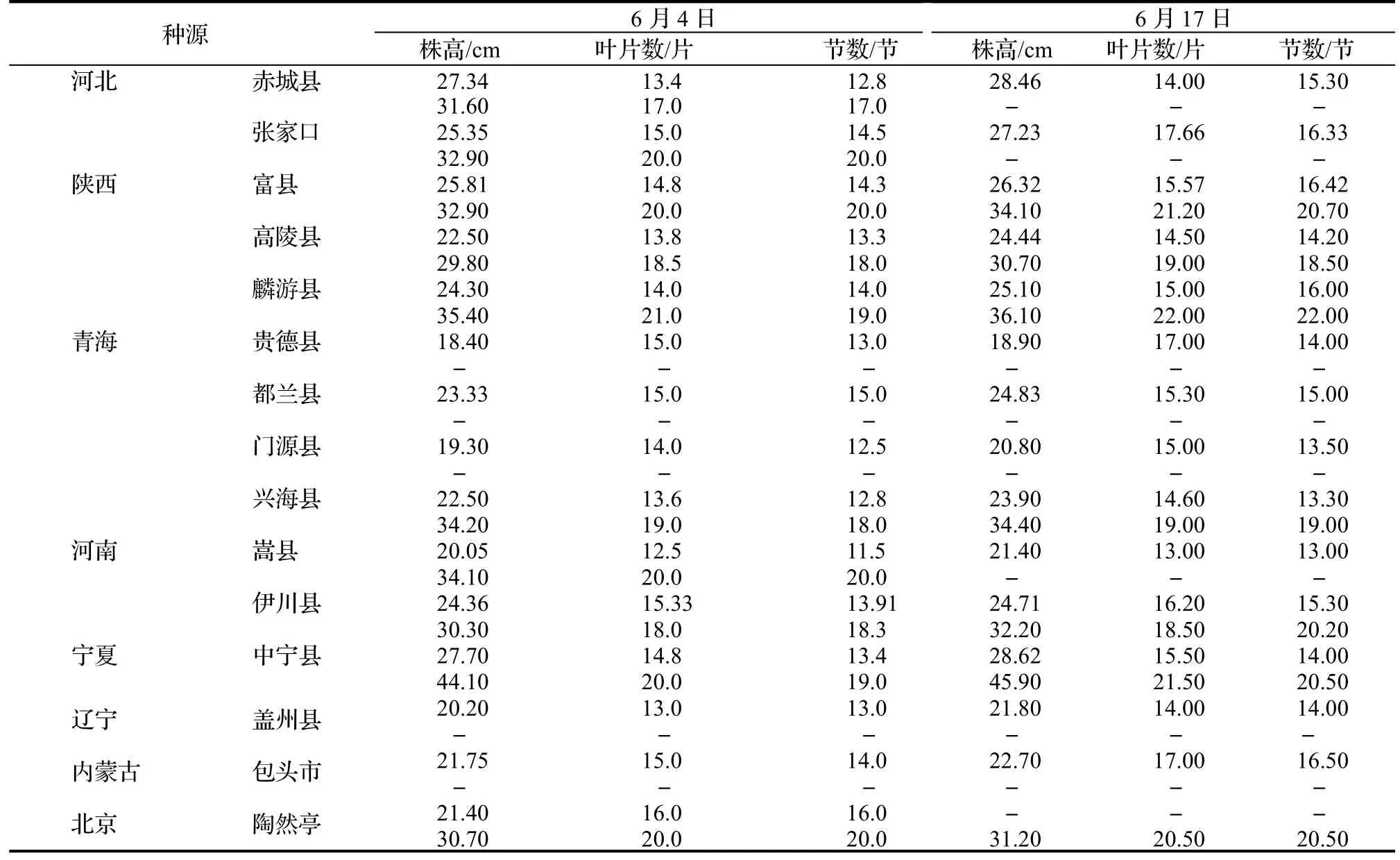
表4 6 月4 日和6 月17 日小叶杨幼苗平均株高、叶片数和节数Table 4 Average height,number of leaf and internode of P.simonii seedlings on June 4th and June 17th
2.3 小叶杨幼苗不同种源间表型变异分析
由表5 可知,株高、叶片数和节数3 个性状种源间差异明显。陕西麟游县种源在株高、叶片数和节数上指标值均最大,平均海拔为1 271 m;陕西富县株高、叶片数和节数在种源间差异次之,平均海拔为1 200 m。对同在西北地区的青海省和宁夏回族自治区而言,宁夏中宁县种源在株高性状上差异均值仅略大于陕西麟游县,平均海拔为2 955 m;贵德县种源均值在株高和节数性状上小于麟游县,平均海拔为2 200 m,都兰县和兴海县种源在株高性状上表现相似且均值均小于贵德县,平均海拔分别为3 180 m 和3 924 m。整体表现为西北地区的这些种源株高较高和节数较多,随着海拔升高,株高和节数值均降低。辽宁盖州种源在株高和叶片数性状上最小,表现为辽宁地区种源株高较矮,叶片数较少。
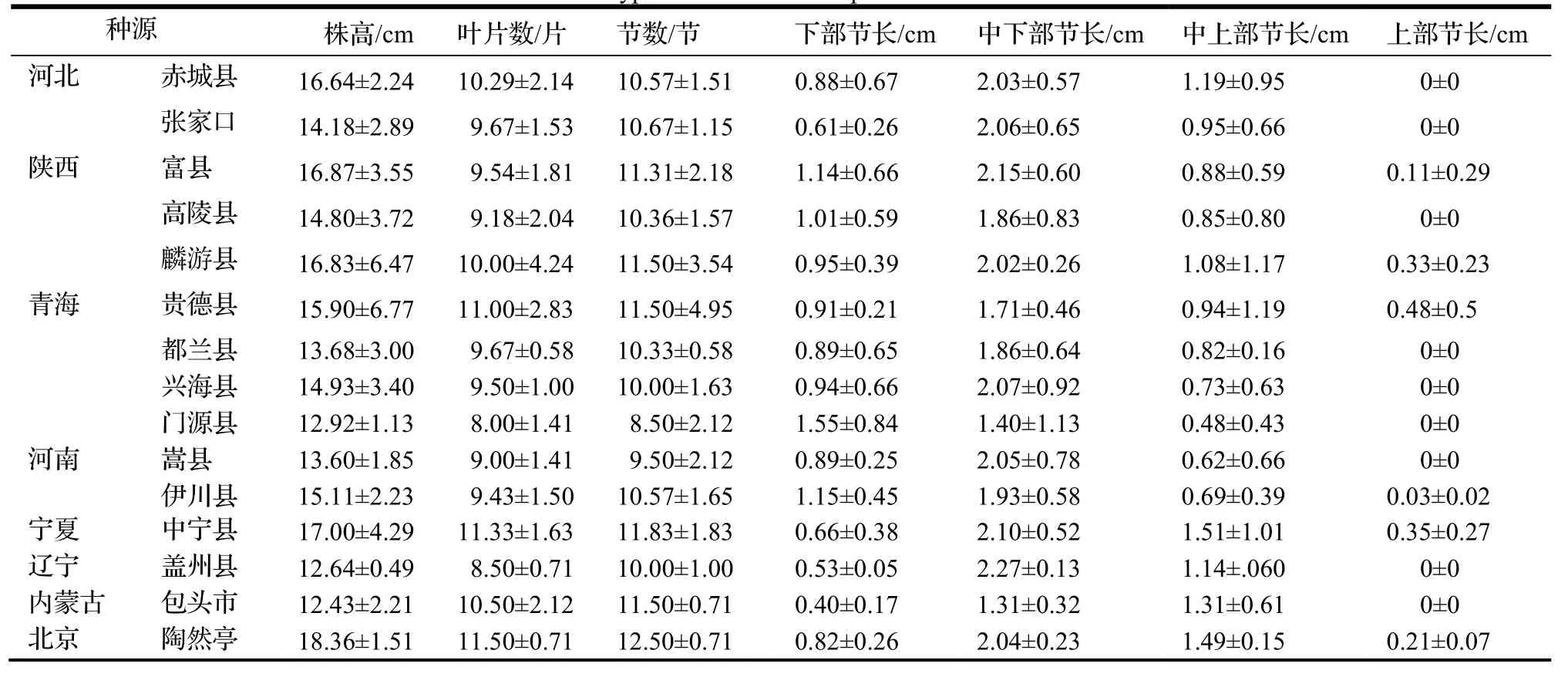
表5 小叶杨表型性状的描述性统计分析(平均值±标准差)Table 5 Phenotypic traits of different provenances of P.simonii
小叶杨节长性状在种源间也存在很大差异,辽宁盖州种源中下部节长均值在种源间差异最大,但标准差差异最小;而西北地区陕西富县、麟游县和宁夏中宁县种源中下部节长均值次之,标准差差异明显;青海门源县种源中下部节长标准差最大,表现为辽宁地区种源中下部节长种源间占植株比例波动较小,位于西北地区的4个种源中下部节长占比差异较大。北京陶然亭公园种源中上部节长均值较大,平均海拔为43.5 m,河北赤城县和内蒙古包头市种源次之,且赤城县种源均值大于内蒙古包头市种源,两地区平均海拔分别为945 m和2 000 m,表现为华北地区种源中上部节长占植株比例较大,随着海拔的升高,中上部节长比例呈减小的趋势。
2.4 小叶杨幼苗种源内表型变异分析
从表6 可知,性状变异都受地区差异影响,以叶片数变异系数波动较小,中上部节长变异系数最大,中下部节长次之。株高、叶片数和节数变异系数小于4 个节长公因子(下部节长、中下部节长、中上部节长和上部节长)变异系数,表明株高、叶片数和节数性状较稳定,受较高的基因控制,4 个节长公因子稳定性差,有较多变异可能性。
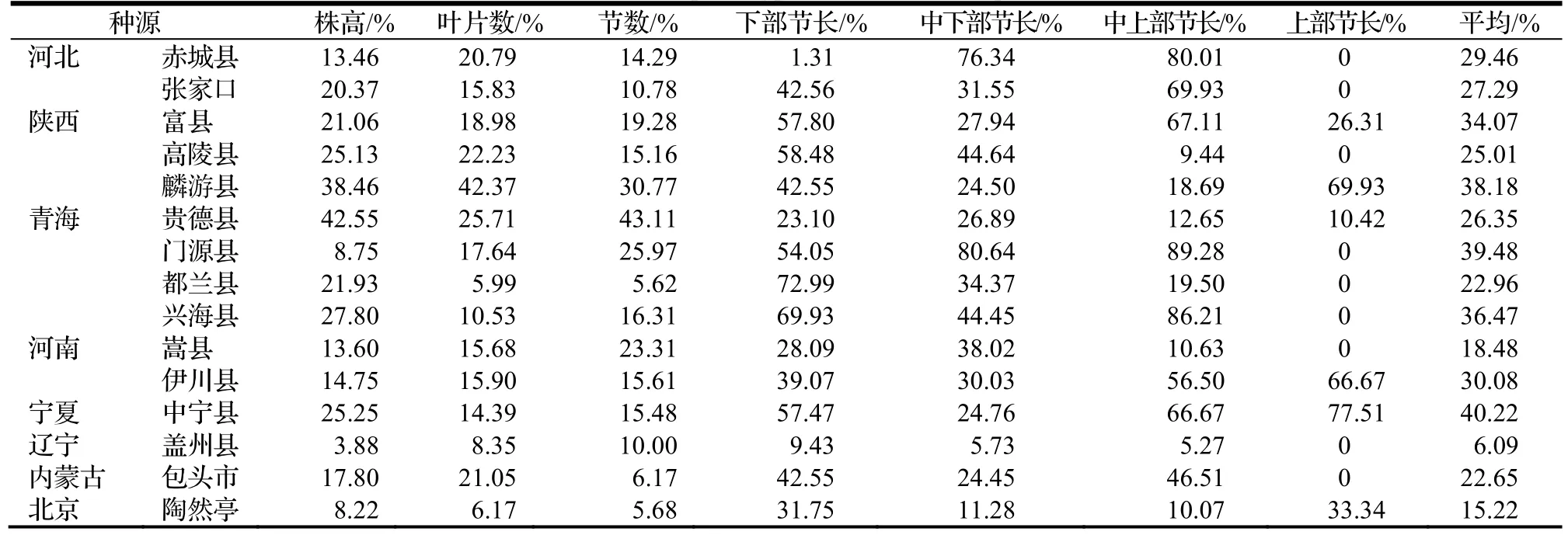
表6 小叶杨表型性状的变异系数Table 6 Coefficient of variation of phenotypic traits of different provenances of P.simonii
对不同种源变异综合分析可知,平均变异系数为6.09%~ 40.22%,变化幅度较大。其中西北地区宁夏中宁县、青海门源县和陕西麟游县种源性状变异系数较大,平均变异系数分别为40.22%,39.48%和38.18%。说明西北地区表型多样性较丰富,辽宁盖州县平均变异系数最小(6.09%),表明东北地区种源多样性较低。河北赤城县、张家口和内蒙古包头市平均表型变异系数在22.65%~ 29.46%,波动不大,三个地区年均降水量在400~421.8 mm,表明这些地区种源多样性相似,与相似的气候条件有关。
2.5 小叶杨幼苗种源间表型分化分析
由表7 可以看出,株高在种源内方差分量较大,叶片数和节数次之且差异相似,4 个节长公因子分化幅度较小。各性状分化幅度为6.9%~ 12.2%,其中株高的分化系数最大为12.2%,节数的分化系数也达到了11.9%,叶片数的分化系数最小为6.9%。各个性状在种源内的方差明显大于种源间方差约4.56 倍,说明表型特征在种源内变异大于种源间变异且主要来源于种源内变异。7 个性状的平均表型分化系数为17.6%,株高和节数性状占较大比例,表明株高和节数在幼苗种源内变异较大。
3 讨论
目前对小叶杨幼苗节间长度性状的分析尚未见报道,而节间长度也反映了幼苗的生长状况,如王健等[6]探讨了普通小麦节间长度与株高的关系,结果表明矮秆小麦是通过缩短各节间长度来降低株高的。所以通过探究小叶杨节间长度之间的关系可以得到影响幼苗生长的重要部位,由于试验得到4 个时期上部节长(D 部分)和中上部节长(C 部分)的方差贡献率均比中下部节长(B 部分)和下部节长(A 部分)高,可推测这两部分在幼苗生长中占据重要地位,对下一步群体基因组变异研究具有借鉴意义。
对小叶杨幼苗种源进行表型性状的变异分析发现,15 个种源的7 个性状在种源内和种源间均存在很大差异,且种源内变异大于种源间变异,这与卫尊征等[12]对小叶杨5 个群体的研究结果和Du 等[3]对毛白杨表型性状的变异分析结论一致。这表明表型变异是小叶杨种源甚至杨属Populus的普遍遗传特性。这种形态特征的变异有利于小叶杨幼苗适应环境,种源内基因型的丰富性表现为表型性状的多样性,使种源能够适应不同的环境[16],这也许造就了小叶杨生长优良、寿命长、能在复杂气候条件下生存的特点。
4 结论
对15 个种源小叶杨一年生幼苗表型性状分析发现节间长度性状间存在很大联系,将23~ 26 个节长性状提取为4 个公因子,将植株整体分为了4 部分,且每部分之间都存在较强的相关性,即由相邻节长组成(载荷较大),说明幼苗相邻节间长具有较稳定的遗传特征。4 个时期上部节长(D 部分)和中上部节长(C 部分)的方差贡献率均比中下部节长(B 部分)和下部节长(A 部分)高,说明幼苗D 和C 部分形态特征变化较大,这与植株自身生长情况相符。
通过对小叶杨幼苗均值和标准差统计结果得到西北地区种源株高较高和节数较多,随着海拔升高,株高和节数值均降低;辽宁盖州种源在株高和叶片数最小,表现为辽宁地区种源株高较矮,叶片数较少。此结果和文中KABC 聚类算法得到的结果相同,聚类结果中第二类样本主要分布在西北的陕西、青海和宁夏等地,平均株高、平均叶片数和平均节数值在4 个时期明显比第一类样本大。
研究结果也表明,小叶杨幼苗种内也存在极其丰富的种源间和种源内变异,15 个种源幼苗的7 个表型性状平均变异系数在6.09%~ 40.22%,变异幅度较大,其中以中上部节长度变异系数最大,株高、叶片数和节数变异小于4 个节长公因子变异系数,表明株高、叶片数和节数性状受较高的基因控制,4 个节长公因子变异较丰富。其次,7 个表型性状分化幅度为6.9%~ 12.2%,株高的分化系数最大为12.2%,各个性状在种源内的方差分量明显大于种源间方差分量,说明表型性状在种源内变异大于种源间变异,即种源内的多样性程度大于种源间的多样性程度。
猜你喜欢
安徽农业大学学报(2022年3期)2022-10-25
绿色科技(2021年21期)2021-11-26
南方林业科学(2021年2期)2021-05-27
辽宁林业科技(2021年1期)2021-03-16
农家科技中旬版(2020年4期)2020-07-06
农村百事通(2019年17期)2019-10-08
作文周刊(高考版)(2016年16期)2017-06-01
现代农业科技(2017年1期)2017-03-06
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
