
基于RAD-seq简化基因组测序的河南斗鸡遗传进化研究
2020-06-10 09:28:16殷建玫朱云芬李国辉张会永苏一军
家畜生态学报 2020年5期
殷建玫,朱云芬,薛 倩,李国辉,张会永,苏一军,韩 威
(江苏省家禽科学研究所 国家级地方鸡种基因库,江苏 扬州 225125)
河南斗鸡,原产地及中心产区为河南省开封市,1989年被《中国家禽品种志》收录,2000年被列入国家级畜禽品种资源保护名录[1]。河南斗鸡饲养历史悠久,在长期的选择和驯化过程中形成了与其他地方鸡品种截然不同的的外貌特征,具有较强的斗性和抗击打能力,是我国家禽库极其宝贵的遗传资源。
微卫星标记及线粒体DNA序列变异已被广泛应用于地方鸡种的遗传多样性、遗传结构及起源分析[2-3],然而这些DNA标记在整个鸡基因组中的覆盖范围是极其微小的,如鸡微卫星标记仅有约30个,鸡线粒体DNA的D-loop区域全长也仅约1 200 bp,其所代表的基因组遗传变异信息量非常有限。新一代测序技术的发展实现了SNP分子标记的高通量检测[4],为从基因组水平上审视鸡的遗传进化提供了高效准确的研究方法。
本研究利用简化基因组测序(Restriction-site associated DNA sequencing,RAD-seq)鉴定河南斗鸡基因组SNP标记,通过与其他地方鸡品种及引入鸡品种比较,揭示河南斗鸡的遗传多样性和遗传结构,鉴定受选择基因,为开展河南斗鸡种质特性评价、品种资源保护及开发利用提供科学依据。
1 材料与方法
1.1 试验材料
河南斗鸡及对照组18个地方鸡品种(狼山鸡、安义瓦灰鸡、固始鸡、仙居鸡、白耳黄鸡、东乡绿壳蛋鸡、金湖乌凤鸡、文昌鸡、大围山微型鸡、藏鸡、茶花鸡、瓢鸡、惠阳胡须鸡、大骨鸡、萧山鸡、鹿苑鸡、边鸡、北京油鸡)、2个引入鸡品种(隐性白羽肉鸡、安卡红鸡)来自于国家级地方鸡种基因库(江苏)。每个品种按照家系选择30个个体(10公、20母),个体间无亲缘关系。翅静脉采血1~1.5 mL,柠檬酸钠(ACD)抗凝,-20 ℃保存备用。
1.2 试验方法
1.2.1 RAD-seq 简化基因组测序 常规酚-氯仿法提取基因组DNA,构建pair-end文库(300~500 bp)进行RAD-seq双酶切简化基因组测序。
1.2.2 SNP 质控 SNP的鉴定采用GATK和samtools程序。质控条件包括:Q20>95%,ddRAD depth>60%,SNP Call rate >90%,MAF>0.05。
1.2.3 统计分析 连锁不平衡(LD)分析采用Haploview软件,观察杂合度(Ho)、近交系数(Fis)和群体分化指数(Fst)计算采用PopGen软件,群体聚类分析应用MEGA软件UPGMA聚类法。选择信号检测采用PLINK软件,以100 kb为窗口、10 kb为step进行滑动的θπ比率和Fst分布计算。
2 结 果
2.1 基因组SNP鉴定
经过两步数据质控,在河南斗鸡中鉴定出SNP 259,412个(表1)。总体上,SNP平均密度与染色体长度呈正相关,大染色体上SNP分布较多(其中1~7号染色体上SNP数量均超过10,000个),小染色体上SNP分布较少。

表1 SNP在河南斗鸡不同染色体上分布Table 1 SNP distribution on Henan game chicken chromosomes
2.2 群体遗传多样性分析
群体遗传多样性统计量结果(表2)表明,河南斗鸡的观察杂合度为0.1560,核苷酸多样度为0.1752,近交系数为0.1099。与其他地方鸡种相比,河南斗鸡的遗传多样性相对较低,与LD连锁不平衡分析结果(图1)相一致。
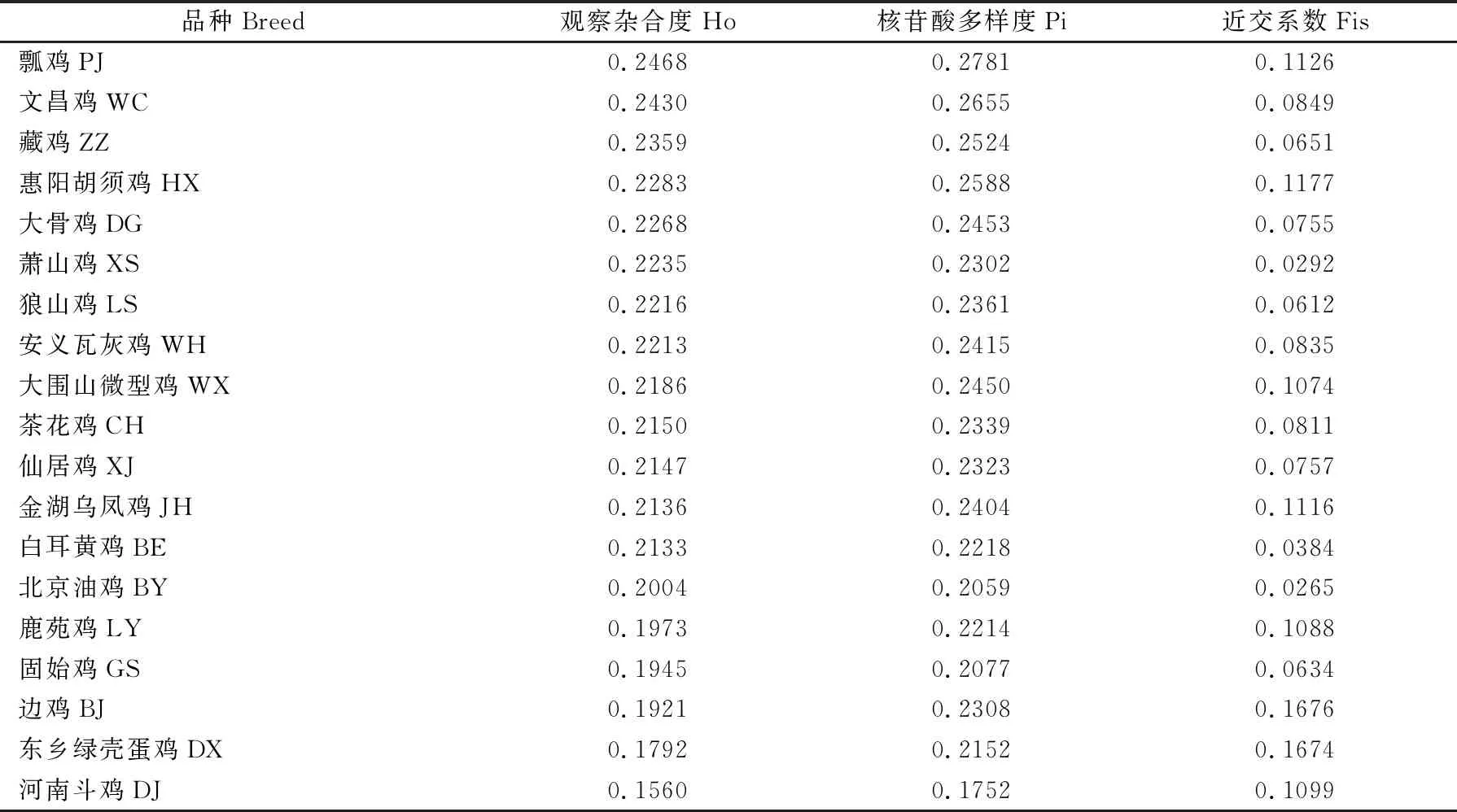
表2 观察杂合度、核苷酸多样度和近交系数Table 2 Statistics of HO, Pi and Fis
2.3 群体遗传结构分析
河南斗鸡与瓢鸡之间的遗传分化系数Fst最低(0.1765),其次为文昌鸡(0.1932)、惠阳胡须鸡(0.1980),相对应的基因流Nm分别为1.1665、1.0443和1.0125(表3)。河南斗鸡与其它地方鸡种间的平均遗传分化系数Fst大于0.20,基因流Nm均小于1.00。聚类结果(见图2)表明,河南斗鸡形成了一个独立的分支。
2.4 选择信号分析
结合核苷酸多样度θπ比率和遗传分化系数Fst的选择信号分析发现24个染色体区域受到选择,129个受选择基因。功能分析结果表明(图3),受选择基因主要富集在代谢过程、神经系统发育、微管细胞骨架、运动行为、适应性免疫和刺激反应等生物学通路。

表3 斗鸡与其他品种间遗传分化系数和基因流Table 3 Fst and Nm between DJ and other chicken breeds
图2河南斗鸡及其他鸡种聚类图
Fig.2 Phylogenetic tree of DJ and other chicken breeds
3 讨 论
3.1 基因组SNP鉴定和遗传多样性分析
简化基因组RAD-seq测序技术能鉴定出合适数量、广泛覆盖基因组范围的变异位点,被广泛应用于资源群体SNP发掘和遗传多样性研究[5-7]。本文利用RAD-seq测定了19个地方鸡种的基因组序列,通过严格的质控,在河南斗鸡群体中鉴定出SNP标记259,412个,这为在基因组水平上分析河南斗鸡遗传进化提供了基础。
斗鸡是我国地方鸡种中一类特殊的品种资源,由于主要是用于玩赏打斗,加之驯养方法非常保守(主要是靠斗鸡爱好者小规模的家庭饲养),因此斗鸡在品种形成过程中与其它地方鸡种间鲜有血缘交流。本文的遗传结构分析结果很好地验证了这一点,在19个地方鸡种中河南斗鸡的平均遗传分化系数Fst 最高(>0.20)、平均基因流Nm 最低(<0.10),在系统发育树中形成了一个独立的分支。在19个地方鸡种中河南斗鸡的观察杂合度Ho 和核苷酸多样度Pi 最低,这与河南斗鸡资源群体较小的表型变异相一致,已报道的基于微卫星DNA及线粒体DNA研究结果也发现河南斗鸡的遗传多样性相对匮乏[8]。河南斗鸡群体的近交系数Fis(0.1099)处于较高水平,这提示在河南斗鸡的保种过程中,应严格控制近交水平的过快增加,保持遗传多样性的相对稳定。
3.2 基因组选择信号分析
自然选择和人工选择作用会在动物基因组上留下选择信号[9],通过选择信号分析,可以发掘重要的功能基因。目前,研究者通过基因组测序或芯片检测,已经对不同物种开展了选择信号研究[10-13],发掘出大量种质特性相关基因。针对鸡而言,Rubin等[14]在家鸡中鉴定了与代谢调节和生殖光周期调控相关的促甲状腺激素受体基因TSHR,在肉鸡中发现与生长、食欲和代谢调控相关的基因重叠在一起。Elferink等[15]在商业化品系鸡、荷兰及中国地方鸡种中发现396个染色体区域受到选择作用,其中26个区域显示出强烈的选择信号。Wang等[16,17]发现藏鸡群体中受到选择作用的基因在钙离子信号通路中显著富集,大量视觉相关基因在家鸡驯化过程中受到正选择作用,使视觉退化。
本文中,选择信号分析结果表明河南斗鸡基因组中有24个染色体区域受到选择作用,受选择基因显著富集于不同的生物学通路。斗性行为是河南斗鸡在进化过程中形成的最显著的特性,分析发现多个参与下丘脑-垂体-肾上腺/甲状腺轴调控通路及激素介导信号通路的基因CRH、ADRB1、SLC64A、DRD1、SORCS2等受到选择作用。肾上腺素、多巴胺、5-羟色胺及雄激素等已被证实是调控攻击行为的主要神经递质[18],Vps10p蛋白家族成员SORCS2是调节鸡攻击行为的重要候选基因[19]。与成骨细胞分化、骨生长及矿化相关的基因FBN2[20]、DDR2[21]和TWSG1[22]受到了选择。这些基因的作用可能使得河南斗鸡具有较其它鸡种更粗壮致密的骨骼。与头部发育相关的基因MEF2C、Wnt6也受到了选择,Verzi等[23]研究发现由MEF2C与DLX5-6形成的前馈环是颅面骨发育中所必须的,Wnt基因在鸡眼睛结构的发育中发挥着重要作用[24],这些基因的作用形成了河南斗鸡头盖骨厚、脸部狭长及独特的眼部结构特征。
心室发育与心脏收缩力调节基因FOXF1、MYL3,氧化还原酶活性基因PTGS2及线粒体呼吸链基因NDUFV2、STEAP4也受到了选择作用,发达的心血管系统和能量代谢系统使得河南斗鸡能够适于长时间激烈的打斗。另外,视觉及声音感知基因RGS9BP、PDE6H,运动行为调节基因DSCAM也显示出强烈的选择信号,表明河南斗鸡仍保持着野外生存状态下必须的机警与敏感。
研究结果为全面评价河南斗鸡种质特性提供了重要遗传信息,诸多受选择基因的功能尚未有清晰的注释,需要进一步的深入研究。
猜你喜欢
小学生作文(低年级适用)(2020年6期)2020-07-24 08:36:18
北极光(2019年12期)2020-01-18 06:22:10
意林(2019年17期)2019-10-07 12:28:52
上海农业学报(2019年4期)2019-09-18 06:46:48
青年歌声(2019年7期)2019-07-26 08:34:58
金桥(2018年3期)2018-12-06 09:05:34
文体用品与科技(2017年5期)2017-05-17 05:31:04
浙江农业学报(2017年4期)2017-04-26 02:28:22
人大建设(2017年11期)2017-04-20 08:22:39
人大建设(2017年5期)2017-04-18 12:57:43
