
外源氮输入及泥沙沉积对黄河三角洲碱蓬生物量分配及抗氧化性酶活性的影响
2020-06-08 11:01庞晓蝶宋红丽王岩
安徽农业科学 2020年10期
关键词:生物量
庞晓蝶 宋红丽 王岩
摘要 [目的]研究外源氮輸入及泥沙沉积对黄河三角洲碱蓬生物量分配及抗氧化性酶活性的影响。[方法]以黄河口滨岸潮滩湿地先锋植被碱蓬(Suaeda salsa)为研究对象,基于模拟试验研究不同氮输入(0、3、6、9 g/m2)和泥沙沉积(0、3、6、12 cm)对碱蓬生物量分配及抗氧化酶活性的影响。[结果]适度的埋深会使植物根系所占的比例增加,也使碱蓬叶绿素含量增加,利于碱蓬生长。适当的N输入,会促进碱蓬的生长,使蛋白质含量和总生物量增加。而过度深埋和N输入则会对碱蓬生长产生抑制作用。泥沙沉积和外源N输入对抗氧化酶活性有一定的影响,表现为泥沙沉积为0和3 cm时,过氧化氢酶(CAT)、过氧化酶(POD)和超氧化歧化酶(SOD)活性增强,但随着泥沙沉积厚度的增加,3种酶的活性减弱。当外源N输入量为6和9 g/m2时,随埋深深度的增加,3种抗氧化酶的活性均呈下降趋势。其中POD和SOD活性均在N3M3(N为9 g/m2,泥沙深度为12 cm)时明显降低,分别比对照低41.77%和34.08%,CAT活性在埋深深度为6 cm和最大N输入 (9 g/m2)时出现最小值 (2.38 U/mg)。方差分析结果表明,N输入、泥沙沉积及其交互作用对碱蓬总生物量及CAT活性具有显著影响,而N输入、泥沙沉积对POD活性具有显著影响(P<0.05)。[结论]调水调沙带来的外源N输入和泥沙沉积对黄河口滨岸潮滩湿地先锋植被碱蓬生长带来一定的影响。在一定范围内,碱蓬能够通过自身的生理变化,适应多变的环境,但当环境变化超过一定限度,就会对碱蓬的正常生长造成威胁。
关键词 N输入;泥沙沉积;碱蓬;生物量;抗氧化酶活性
中图分类号 X171 文献标识码 A
文章编号 0517-6611(2020)10-0071-05
doi:10.3969/j.issn.0517-6611.2020.10.020
Abstract [Objective]The aim was to study the effects of exogenous nitrogen input and sediment burial on biomass allocation and antioxidative enzyme activities of Suaeda salsa in the coastal wetland of the Yellow River delta.[Method]In this study,we selected S.salsa which is a pioneer vegetation in the coastal wetland as the research object,and based on the simulation experiment,to research the effects of different N inputs (0,3,6,9 g/m) and sediment burial depths (0,3,6,12 cm) on biomass allocation and antioxidant enzyme activity of S.salsa.[Result]The results showed that root proportion and chlorophyll content of S.salsa increased under the moderate burial depth,which was beneficial to the growth of S.salsa.Appropriate N input also promoted the growth of S.salsa and increased its protein content and total biomass,while excessive burial and N input would inhibit S.salsa growth.N input and sediment burial also influenced antioxidant enzymes activities,and showed that the activity of CAT,POD and SOD increased when the sediment burial is 0 and 3 cm,while the activity of the three enzymes decreased with the increase of stress.When the input of exogenous N was 6 and 9 g/m2,the activity of the three antioxidant enzymes showed a decreasing trend with the increase of burial depth.Among them,the activity of POD and SOD was significantly reduced when the N input amount was 9 g/m2 and sediment burial was 12 cm,which were 41.77% and 34.08% lower than the control.CAT activity obtained a minimum value (2.38 U/mg) at the burial depth of 6 cm and the maximum N input (9 g/m2).The results of ANOVA showed that:N input,sediment burial and its interaction had significant effects on the total biomass and CAT activity of S.salsa,while N input and sediment burial had significant effects on POD activity (P<0.05). [Conclusion]We concluded that the exogenous N input and sediment burial brought by water and sediment regulation had certain influence on the growth of S.salsa in the coastal wetland of the Yellow River estuary. S.salsa could adapt to the changing environment through its own physiological changes within a certain range,but the environmental changes exceeded a certain value would threat the normal growth of S.salsa.
Key words N input;Sediment burial;Suaeda salsa;Biomass allocation;Antioxidant enzyme activity
湿地介于水体与陆地之间,是一个比较特别的生态交错区,与森林、海洋并称地球3大生态区,具有非常丰富的物种,是我国非常重要的自然资源与国土资源。黄河三角洲作为我国温带地区重要的河口海岸带湿地生态系统,是最具生态保护价值的关键区域之一[1]。在全球变化和人为活动强度日益增强的影响下,滨海湿地结构和功能呈现出不同程度退化状态[2]。尤其是20世纪黄河出现多次断流现象,造成河口湿地结构功能发生改变,天然湿地面积萎缩、生态系统退化、水体污染、土壤盐碱化等一系列生态环境问题,严重制约了河口地区的可持续发展[3]。为恢复黄河三角洲退化湿地,自2002年起小浪底工程,每年进行为期20 d左右的调水调沙。调水调沙的实施,为黄河三角洲带来大量淡水恢复退化湿地的同时,也带来大量泥沙和外源氮的输入,改变了三角洲植被的生存环境。
氮(N)是影响植物生长发育的必需大量营养元素之一,是叶绿素、氨基酸、核酸、次生代谢产物的重要组分[4-5]。缺氮抑制植株生长,造成地上部分生物量显著减少,叶片瘦小、黄化[6],影响植物的正常生长。但是过量的N输入,同样会抑制植物生长。植物吸收的N主要是铵态氮和硝态氮[7]。近年来,随着经济的快速发展,各种污染物的排放量呈增加趋势,据《中国海洋环境状况公报》显示,2010—2014年黄河口入海污染物达1.90×105~5.70×105 t,其中氨氮、硝酸盐氮和亚硝酸盐氮含量占总污染物总量的比例高达4.02%~16.17%,这可能对湿地和近岸生态系统的植物生长和养分循环等产生深刻影响[8]。
沉积是影响滨海湿地植物生长的一个重要因素[9],调水调沙带来的大量泥沙会改变植被原本的生存环境,适宜的浅埋会促进植物的生长[10],但是过度的深埋会阻断幼苗接受外界充足的空气、光照和温度等,从而影响植株生长。植被存在沉积耐受的阈值,在这个阈值之下的沉积会促进种子萌发和植被生长,而超过这个阈值,种子萌发和植被生长会受到严重制约[11]。
目前很多学者针对单因素泥沙沉积或者外源N输入对植被的影响开展了大量研究[8-9,12-13],但单因素并不能全面反映调水调沙带来的影响。为此笔者以黄河三角洲的先锋植被碱蓬(Suaeda salsa)为研究对象,通过模拟试验探索泥沙沉积和外源N输入共同作用对该区典型植被生长的影响。并通过分析碱蓬总生物量、生物量分配及抗氧化酶活性的变化情况,揭示调水调沙带来的泥沙及N对植物生长的影响,研究结果可为调水调沙提供一定的数据基础,实现调水调沙最大的环境效益和生态效益。
1 研究内容与方法
1.1 研究區概况 黄河三角洲自然保护区为1992年经国务院批准建立的国家级自然保护区,总面积15.3 hm2,地理坐标118°33′~119°207′ E,37°35′~38°12′N。北靠渤海,东临莱州湾,位于江淮平原和东北亚内陆之间。该地区为暖温带季风型大陆性气候,夏季炎热多雨,冬季寒冷、降水较少。年降水量为551.60 mm,蒸发量为1 928.20 mm。土壤为隐域性潮土和盐土土类。黄河三角洲是我国三大河口三角洲之一,是我国最年轻、面积最大、保存最完整的新生河口湿地生态系统[14]。保护区内植被丰富,因地处黄河流入渤海的交汇处,水文条件独特,海淡水交汇,离子作用促进泥沙的絮凝沉降,形成了宽阔的湿地。吸引大量过境和栖息繁殖的鸟类,提供大片植物生长的土地,具有重要的研究价值。
1.2 试验材料
供试碱蓬种子采自黄河口滨岸潮滩湿地的碱蓬植物群,2016年11月中旬采集完成处理干净后,带回临沂大学实验室,放置于4 ℃的冰箱内保存备用。试验所用土壤是黄河三角洲生长碱蓬的0~30 cm表层盐碱土。
1.3 试验设置
基于微区模拟平台(位于山东省临沂大学水土保持与环境保育研究所),于2017年4月底,在每个直径为25 cm的模拟小区中,均匀播种30粒籽粒饱满的碱蓬种子,为保持土壤湿润,每天用烧杯浇水且每个模拟小区补充水量一致。分3次去除长势不好的幼苗,最终保留6株长势相同的植株。
历年调水调沙时间为6月底~7月初,因此在6月22日对试验植株进行处理。根据以往研究结果,调水调沙期间在潮滩湿地泥沙沉积的厚度5~6 cm[15]。当埋深深度小于植株高度的2/3时,对碱蓬生长具有一定的促进作用[12]。因此试验设置4个泥沙埋深:0(M0)、3 (M1)、6(M2)、12 cm(M3)。黄河三角洲地区外源N输入量为2.5~3.5 g·N/(m2·a)[8],试验共设4个氮添加量,分别为0(N0)、3(N1)、6(N2)和9 g/m2(N3)。该试验共计16组,每组3个重复。将提前准备好所需的3、6、12 cm的细沙倒入对应的M1、M2、M3相应的模拟小区里,将事先称好的NH4Cl(外源氮)以水溶的方式加入对应的N1、N2、N3模拟小区中。于7月6日,采取新鲜的碱蓬叶片,测定叶绿素、蛋白质含量、过氧化氢酶(CAT)、过氧化酶(POD)和超氧化歧化酶(SOD)活性。测定叶绿素a和叶绿素b时,将0.1 g新鲜叶片剪碎,放在混合液(浓度为95%的乙醇与浓度为80%的丙酮按1∶1混合)中浸提24 h,提取液在波长663和645 nm下测定吸光度。利用南京建成生物工程公司试剂盒测定SOD、CAT、POD活性和蛋白质含量。
9月11日,结束试验。为保护原有的根系结构,用蒸馏水将根上的泥土冲洗干净,分离为根、茎、叶分别装入纸质文件袋内。然后将根茎叶放入烘箱于105 ℃ 下杀青后,于80 ℃下烘干至恒重,并称重。
1.4 数据分析 运用SPSS 20.0对数据进行处理,不同泥沙埋深深度、外源N输入量及其相互作用对碱蓬生物量、抗氧化酶活性、叶绿素含量的影响运用多因素方差分析,并在0.05水平下进行显著性分析。运用软件Rrigin 8.0作图。
2 结果分析
2.1 N输入及泥沙沉积对碱蓬生物量及生物量分配的影响
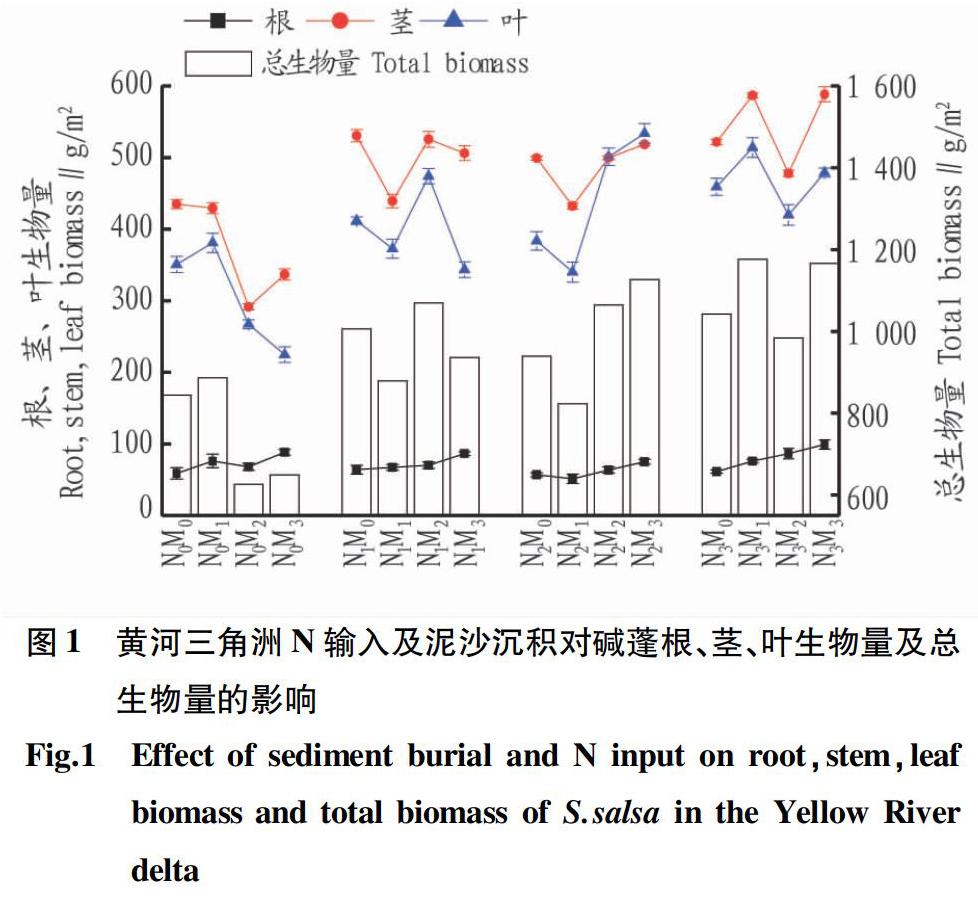
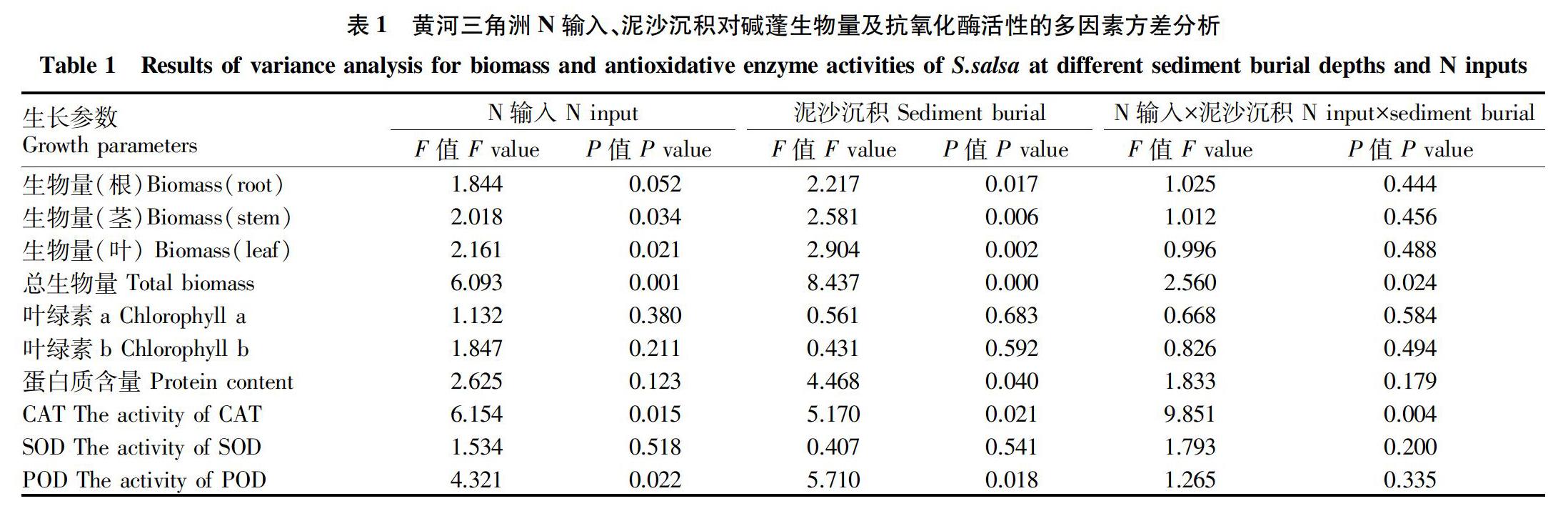
N输入及泥沙沉积对碱蓬生物量的影响如图1所示。当N输入量为N1、N2、N3时,除N2M1(823.44 g/m2)外,碱蓬的总生物量均高于对照组N0M0(844.32 g/m2)。N输入为N3和N0时的变化趋势一致,均随着埋深深度增加,碱蓬总生物量呈现先增加后减少再增加的趋势,在N0M1和N3M1时取得最大值886.35、1 176.53 g/m2,最大生物量N3M1(1 176.53 g/m2)比对照组高39.35%。当N输入量为N1和N2时,总生物量会随着埋深的增加呈现先减少后增加的趨势,在N1M2和N2M3取得最大值1 069.56和1 126.93 g/m2。当埋深深度为M0、M2时,随着N输入量的增加,碱蓬总生物量呈现先增加后减少的趋势。埋深深度最深M3(12 cm)时,随着外源N输入的不断增加,碱蓬的总生物量也一直增加,最大值N3M3(1 166.37 g/m2)比最小值N0M3(649.51 g/m2)高79.58%。方差分析结果表明,外源N输入、泥沙沉积及其交互作用对碱蓬总生物量的影响达显著水平,N输入对茎和叶生物量的影响也达显著水平,泥沙沉积对碱蓬根、茎、叶生物量的影响均达显著水平(P<0.05)(表1)。
外源N输入及泥沙沉积对碱蓬生物量分配的影响主要表现在:随着埋深深度的增加碱蓬根所占的比重呈波动增加,N输入量为N2时变化最不明显,当无外源N输入(N0)和输入量为3 g/m2(N1)时,随着埋深深度的增加,根所占的比例由6.97%和6.37%分别增加至13.59%和9.24%。茎和叶所占比例无明显变化。
2.2 N输入及泥沙沉积对叶绿素含量的影响
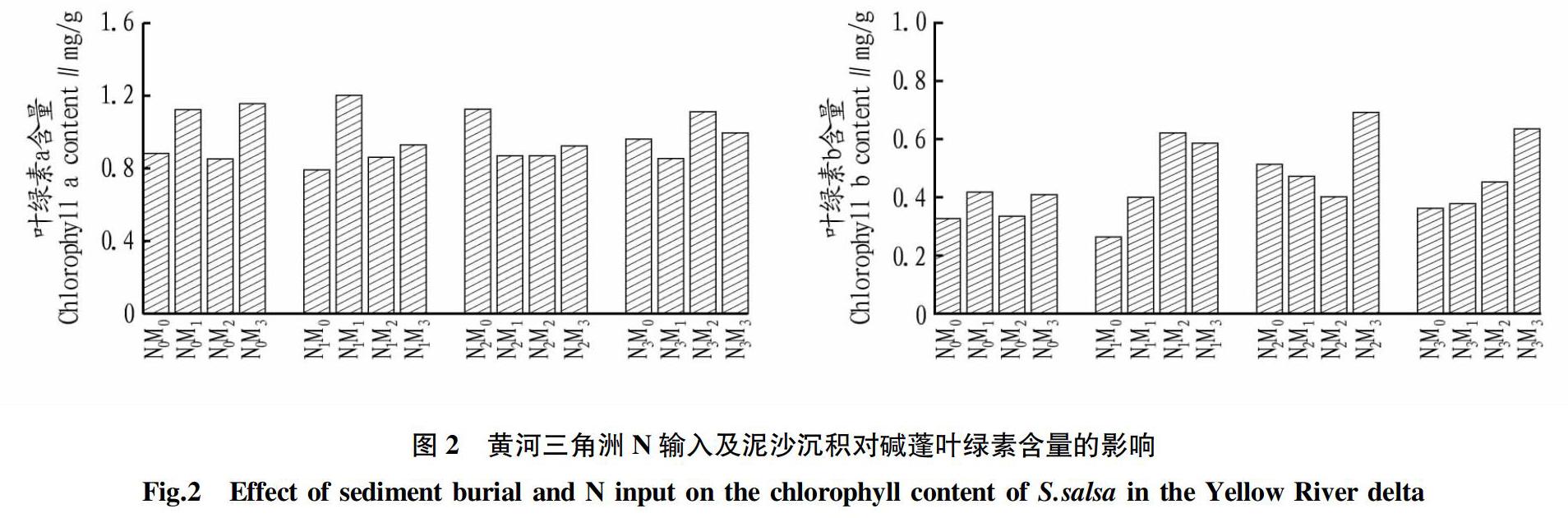
由图2可看出,外源N输入和泥沙沉积对碱蓬叶绿素的含量有一定影响。当N输入量为N1时,叶绿素a的变化趋势与对照组N0的变化趋势相同,均随着埋深深度的增加呈现先增加后减少再增加,并在N1M1时叶绿素a取得最大值(1.20 mg/g)。N输入量为N2和N3时,随埋深深度的增加,碱蓬叶绿素a含量先减少后增加。在埋深深度相同时,随着N输入的增加,叶绿素a的含量变化规律不明显。
N输入量为N0和N1 时,随埋深深度的增加,叶绿素b的含量呈先增加后减少的趋势,其中N1M2 (0.62 mg/g)比对照组N0M0(0.33 mg/g)的叶绿素含量高89.77%。在N输入量为N3时,随着埋深深度的增加,叶绿素b的含量一直增加,由N3M0(0.36 mg/g)到N3M3(0.64 mg/g)增加75.37%。埋深深度相同时,随N输入量的增加,叶绿素b的含量呈波动增加然后减少的趋势,除N1M1外,所有处理均高于对照组。方差分析结果表明:N输入、泥沙沉积及其交互作用对碱蓬叶绿素a和叶绿素b含量的影响均未达显著水平(P>0.05)(表1)。
2.3 N输入及泥沙沉积对碱蓬抗氧化性酶活性及蛋白质含量的影响
外源N输入及泥沙沉积对碱蓬CAT、POD和SOD 3种抗氧化性酶的影响如图3所示。在外源N处理下,随着埋深深度的增加,3种抗氧化性酶的活性波动趋势基本一致,含量均先增加后降低。当N输入量在N0和N1时,随着埋深深度的增加,3种酶的活性均先增强后减弱再增强,CAT、SOD、POD均在N1M1取得最大值(分别为16.29、14.91、68.77 U/mg)。在N输入为N2、N3时,随着埋深深度的增加,酶的活性均呈现一种波动减少再增加的趋势。当外源N输入较高(N2和N3)时,泥沙埋深较深(M2)时,3种抗氧化性酶的活性显著减少。方差结果分析表明,N输入、泥沙沉积及其交互作用对CAT活性的影响均达显著水平(P<0.05),三者对SOD活性的影响均未达显著水平(P>0.05),N输入和泥沙沉积对POD活性的影响达显著水平(P<0.05)(表1)。
图3a表明,随着埋深的增加,碱蓬的蛋白质含量都呈现先增加后减少的趋势,N2M1时增加的比例最大(比N2M0增加78.2%)。当埋深深度为M0和M1时,随着N输入的增多,碱蓬的蛋白质含量也呈增加趋势,并在N2M1时取得最大值43.21 mg/g。方差分析结果表明:泥沙沉积对碱蓬蛋白质含量的影响达到显著水平(P<0.05)。
3 讨论
碱蓬是一种典型的盐碱地指示植物,盐地碱蓬有改善土壤孔隙度、容重等物理性质和含水量的功能,在不同类型盐碱地上种植碱蓬后,土壤的化学性质均得到一定程度的改善,除此之外,碱蓬对改良土壤微生态环境还有良好的效果[16-17]。研究碱蓬对保护黄河三角州湿地环境显得尤为重要。在快速生长的6、7月份,碱蓬会受到调水调沙工程带来的泥沙[18]和一定的外源物质(如氮元素),以及潮汐带来的泥沙作用[19]。在野外条件下,很难直接研究泥沙沉积和外源氮输入对碱蓬生长的影响,因为泥沙沉积、外源N输入对植物的影响非常复杂并且很容易受盐分、温度、水分条件等其他因素的干扰。因此,笔者利用模拟试验,探讨外源N输入及泥沙沉积对碱蓬总生物量、生物量分配比及抗氧化性酶活性的影响。
3.1 对总生物量及生物量分配比的影响
在有外源N输入的情况下,除N2M1(823.44 g/m2)外,碱蓬的总生物量均高于对照组(N0M0),特别是在N3M1,碱蓬的总生物量(1 176.53 g/m2)比对照组高39.35%,说明外源氮输入能够促进盐地碱蓬生物量的积累。当埋深深度为M0、M2时,随着N输入量的增加,碱蓬总生物量呈现先增加后减少的趋势。该结果与刘德燕等[20]的研究结果一致,小叶章地上部分生物量均随着外源氮输入量的增加而增大,但是连续高氮处理的小叶章地上部分生物量呈先增后减的趋势。氮作为生物体生命活动必需的大量元素之一,其供应影响植物的生长水平。适当的外源氮输入可以促进植物的生长,但当植物生长所需的氮达到饱和状态时,过量的外源氮输入则会抑制植物体的生长。氮素也是植物生理代谢过程中起催化作用的主要物质[21],没有这些催化物质,植物体便不能正常生长,N元素通过这些催化物质间接影响着植物生长和发育。
当N输入为N0和N3时,随着埋深深度的增加,碱蓬总生物量呈现先增加后减少的趋势,并均在N0M1、N3M1取得最大值,证明M1(3 cm)的埋深深度是最适合碱蓬生长的。研究表明:适当的浅埋对碱蓬的生长有促进作用,但过量的深埋会对碱蓬的生长产生抑制作用,这一结论与Sun等[13]的结论一致。适当的浅埋能促进植物根系的生长,故根系所占的比例增加,该研究中在无外源N输入(N0)时,随着埋深深度的增加,根所占的比例由6.97%增至13.59%。根系的增加能使碱蓬从外界吸收更多的营养物质,用于自身的生长发育,因此会使碱蓬的总生物量增加。这一研究结果与李秋艳等[22]沙埋对红砂幼苗出土和生长的影响结果一致,即生物量分配随沙埋深度增大会更多地分配给地下部分,掩埋部分的根状茎和根的干重增加。
3.2 对叶绿素含量的影响 该研究中4个不同的埋深深度,随N输入量的增多,叶绿素b的含量均呈波动增加然后减少的趋势,除埋深深度为6 cm(M2)时,均在N输入为6 g/m2时,取得最大值,说明N输入为9 g/m2时,对碱蓬叶绿素b的产生有一定的抑制作用。研究结果表明:适当的N输入会促进碱蓬叶绿素含量的升高,但是过度的N输入会抑制植物叶绿素的产生。这与李斌等[23]对闽楠幼苗的研究结果一致,随着氮浓度的增加,叶绿素总含量出现先升高后降低的变化趋势,即在适当供氮水平下,闽楠幼苗的叶绿素含量呈现上升趋势,对植物的生长状况起促进作用,但当外界的氮素输入超过阈值后,叶绿素含量出现降低,阻碍植物的合成代谢,对植物生长产生抑制,说明氮素是植物体叶绿素的重要限制因子。在N输入为N0和N1时,叶绿素a和叶绿素b在埋深为3 cm(M1)时的含量均高于不埋深时的含量,特别是叶绿素a含量在N1M1比N1M0高出51.74%,这说明当N输入相同时,不埋深或者适当浅埋会促进叶绿素含量的增多。但是过量的深埋也会使叶绿素的含量降低,光合作用减弱。李强[24]在研究三峡库区消落带掩埋对狗牙根生长时发现,过度的埋深使其光合电子传递速率呈显著降低趋势,热耗散量显著增大,光合作用能力减小,发育呈显著降低趋势。
3.3 对抗氧化酶活性和蛋白质的影响
超氧化物歧化酶(SOD)和过氧化氢酶(CAT)是植物体内的保护酶,两者相互协同对细胞膜有害的氧自由基转化形成无害的分子水和氧,过氧化物酶(POD)存在于植物的各个组织器官中,POD的主要功能是清除SOD和超氧阴离子反应生成的H2O2,保护植物细胞膜系统,从而增强植物的抗逆性[25-26]。邬燕等[27]在模拟干旱胁迫对葡萄生理指标的影响中发现,SOD、POD、CAT 3种酶在消除H2O2方面有重要作用,能够降低超氧阴离子的数值,减少对细胞膜系统的伤害,随着胁迫加深,保护酶SOD、POD、CAT的活性呈现升高趋势,但如果胁迫超出一定范围3种保护酶的活性就会下降,这样就出现了酶活性先升高后降低的变化趋势。这与该文研究的泥沙沉积对碱蓬的影响结果一致,CAT和SOD的活性除N0M1(N为0 g/m2、泥沙为3 cm)和N1M1(N为3 g/m2、泥沙为3 cm)高于对照组外,随着N输入和泥沙埋深深度的不断增加,2种酶的活性降低,其中POD和SOD活性均在N3M3时明显降低,分别比对照低41.77%和34.08%,即适当的浅埋(M1)会使抗氧化酶的活性增强,但是随着胁迫加深,即过度的深埋和N输入,会使抗氧化酶的活性降低。在有外源N输入的情况下,碱蓬的蛋白质含量呈增加趋势,在N2M1(N输入为6 g/m2)时,碱蓬蛋白质的含量增加最明显。王爽等[28]在对大麦的研究中也指出,大麦籽粒蛋白质含量随施氮量增加而上升。在作物生长发育过程中,细胞的增长和分裂以及新细胞的形成都离不开蛋白质的参与,N是构成蛋白质的主要元素,故氮元素的增加会促进蛋白质含量的增加。
4 结论
(1)泥沙沉积及N输入对碱蓬生物量及生物量分配有影响,表现为适当浅埋和N输入促进碱蓬生长,深度埋深及大量N输入抑制碱蓬生长;随着埋深的增加,碱蓬根所占的比例增大,尤其是无外源N输入(N0)和输入量为3 g/m2 (N1)时,随着埋深深度的增加,根所占的比例由6.97%和6.37%分别增加至13.59%和9.24%。
(2)随着外源N的输入,叶绿素b的含量明显增多,特别是N1M2 (0.62 mg/g)比对照组N1M0(0.26 mg/g)的叶绿素含量高136%。在N输入量为N3时,埋深深度的增加,叶绿素b的含量一直增加,由N3M0(0.36 mg/g)到N3M3(0.64 mg/g)增加了75.37%。
(3)轻微泥沙沉积时,CAT、POD和SOD活性增强,随着胁迫的增加,3种酶的活性减弱。在有外源N输入的情况下,碱蓬的蛋白质含量呈增加趋势。
(4)调水调沙过程中带来的外源N输入和泥沙沉积,使碱蓬生存的环境条件发生了改变,对碱蓬的生长产生一定的影响。为适应变化的环境,在一定范围内碱蓬可以改变生物量分配比例以及体内抗氧化酶的含量,这是碱蓬为适应黄河口滨岸潮滩湿地多变环境的一种适应机制。
参考文献
[1]张希涛,毕正刚,车纯广,等.黄河三角洲滨海湿地生态问题及其修复对策研究[J].安徽农业科学,2019,47(5):84-87,91.
[2] 宋红丽,王立志,郁万妮,等.黄河口滨岸潮滩湿地泥沙沉积及外源镉Cd输入对碱蓬物质量分配及抗氧化酶活性的影响[J].环境科学,2018,39(8):3910-3916.
[3] 邢子强.黄河口典型湿地生态需水及生态水文调控[D].青島:中国海洋大学,2011.
[4] ROMME W H,TINKER D B,STAKES G K,et al.Does inorganic nitrogen limit plant growth 3-5 years after fire in a Wyoming,USA,lodgepole pine forest?[J].Forest ecology and management,2009,257(3):829-835.
[5] REDDY K R,MATCHA S K.Quantifying nitrogen effects on castor bean (Ricinus communis L.) development,growth,and photosynthesis[J].Industrial crops and products,2010,31(1):185-191.
[6] ZHANG L H,SONG C C,WANG D X,et al.Effects of exogenous nitrogen on freshwater marsh plant growth and N2O fluxes in Sanjiang Plain,Northeast China[J].Atmospheric environment,2007,41(5):1080-1090.
[7] HU Y L,ZENG D H,LIU Y X,et al.Responses of soil chemical and biological properties to nitrogen addition in a dahurian larch plantation in Northeast China[J].Plant and soil,2010,333(1/2):81-92.
[8] 胡星云,孙志高,孙文广,等.黄河口新生湿地碱蓬生物量及氮累积与分配对外源氮输入的响应[J].生态学报,2017,37(1):226-237.
[9] DENG Z F,AN S Q,ZHAO C J,et al.Sediment burial stimulates the growth and propagule production of Spartina alterniflora Loisel.[J].Estuarine,coastal and shelf science,2008,76(4):818-826.
[10] JARVIS J C,MOORE K A.Effects of seed source,sediment type,and burial depth on mixed-annual and perennial Zostera marina L.seed germination and seedling establishment[J].Estuaries and coasts,2015,38(3):964-978.
[11] ZHANG J H,MAUN M A.Potential for seed bank formation in seven Great Lakes sand dune species[J].American journal of botany,1994,81(4):387-394.
[12] SUN Z G,SONG H L,SUN W G,et al.Effects of continual burial by sediment on morphological traits and dry mass allocation of Suaeda salsa seedlings in the Yellow River estuary:An experimental study[J].Ecological engineering,2014,68:176-183.
[13] SUN Z G,SONG H L,SUN J K,et al.Effects of continual burial by sediment on seedling emergence and morphology of Suaeda salsa in the coastal marsh of the Yellow River estuary,China[J].Journal of environmental management,2014,135:27-35.
[14] 王建步,張杰,吴培强,等.高光谱影像黄河口裸滩与黄河水区分能力评价[J].遥感信息,2015,30(4):81-84.
[15] 牟晓杰,孙志高,王玲玲,等.黄河口滨岸潮滩不同生境下翅碱蓬氮的累积与分配特征[J].湿地科学,2010,8(1):57-66.
[16] 邹桂梅,苏德荣,黄明勇,等.人工种植盐地碱蓬改良吹填土的试验研究[J].草业科学,2010,27(4):51-56.
[17] 敖雁,吴启.3种盐生植物对盐渍响应及利用的研究进展[J].安徽农业科学,2019,47(4):40-45.
[18] 王宁,刘光兴,刘晓彤,等.2010年夏末黄河口及其邻近水域浮游植物群落结构[J].海洋环境科学,2017,36(1):48-55.
[19] CUI B S,YANG Q C,YANG Z F,et al.Evaluating the ecological performance of wetland restoration in the Yellow River Delta,China[J].Ecological engineering,2009,35(7):1090-1103.
[20] 刘德燕,宋长春.湿地植物小叶章对外源氮输入的响应[J].应用生态学报,2008,19(12):2599-2604.
[21] 陶爽,华晓雨,王英男,等.不同氮素形态对植物生长与生理影响的研究进展[J].贵州农业科学,2017,45(12):64-68.
[22] 李秋艳,方海燕.沙埋对红砂幼苗出土和生长的影响[J].水土保持通报,2008,28(1):30-33.
[23] 李彬,洪滔,陈欣凡,等.氮梯度加载下闽楠幼苗生长与叶绿素荧光变化[J].森林与环境学报,2019,39(3):241-247.
[24] 李强.泥沙掩埋和干旱对三峡库区消落带狗牙根生长恢复的影响[J].生态学报,2016,36(1):200-208.
[25] 井大炜.杨树苗叶片光合特性和抗氧化酶对干旱胁迫的响应[J].核农学报,2014,28(3):532-539.
[26] 张军,王文科,耿雅妮,等.黑麦草抗氧化酶对Cd、Zn和Pb复合污染的响应[J].农业环境科学学报,2018,37(6):1117-1124.
[27] 邬燕,刘志华,刘钊.PEG模拟干旱胁迫下4种葡萄生理指标的变化及其抗旱性评价[J/OL].分子植物育种,2019-04-04[2019-05-08].http://kns.cnki.net/kcms/detail/46.1068.S.20190402.1336.004.html.
[28] 王爽,李赢,吕超,等.施氮量和密度对大麦籽粒蛋白质含量及组分的影响[J].分子植物育种,2019,17(11):3788-3793.
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
今日农业(2020年19期)2020-12-14
农家科技中旬版(2020年7期)2020-07-24
甘肃科技(2020年21期)2020-04-13
中国野生植物资源(2019年2期)2019-06-11
湖南林业科技(2017年1期)2017-02-06
中国科技信息(2015年2期)2015-11-16
植物营养与肥料学报(2014年1期)2014-03-11
植物营养与肥料学报(2014年1期)2014-03-11
中国烟草学报(2012年3期)2012-04-10
