
生物蛋白石早期成岩相变特征及对硅质页岩孔隙发育与孔径分布的影响
2020-06-04 02:47卢龙飞刘伟新俞凌杰张文涛申宝剑腾格尔
石油实验地质 2020年3期
卢龙飞,刘伟新,俞凌杰,张文涛,申宝剑,腾格尔
(1.页岩油气富集机理与有效开发国家重点实验室,无锡 214126;2.中国石化 石油勘探开发研究院 无锡石油地质研究所,无锡 214126)
我国中上扬子地区上奥陶统五峰组和下志留统龙马溪组下部发育一套生物成因的硅质页岩[1-3],具有高硅质、高有机碳、高孔隙度和高含气性的“四高”特征[4-6],是页岩气的主要富集层和开发“甜点”层。对该套页岩成因的研究发现其中含有大量的硅质生物——放射虫和海绵骨针的碎屑和残体,属典型的生物成因,由内源生物碎屑的原地沉积作用形成[7]。无独有偶,北美地区多套页岩气富集层也恰好对应于生物成因的硅质页岩层[8-11],显示生物成因硅质页岩与页岩气的富集关系极为密切,表明二者具有普遍的关联性。
成岩作用对储集岩孔隙的形成、演化和保存起着极为重要的控制作用,对储集岩的物性和孔隙结构有决定性影响,作为非常规油气储层的页岩也不例外,其总孔隙度和纳米孔隙的发育依然受控于成岩演化过程。生物成因硅质页岩能够成为优质的页岩气储集岩,正是由于在成岩作用下形成并保存了较高的孔隙和具特征性的孔隙结构[7, 12]。生物成因硅质——蛋白石是一类含水的非晶态矿物,热稳定性极低,随着地层温压条件的变化将向稳定的矿物相石英转化。然而,目前关于高—过成熟演化阶段生物硅质页岩中石英由生物蛋白石转化而来的过程以及该过程对页岩物性与孔隙结构的影响研究还较少,因此,进行相关成岩演化与影响研究对深入认识硅质页岩孔隙演化的全过程非常重要。
我国南方五峰组—龙马溪组页岩现今处于高演化阶段[13-15],其复杂的成岩过程较难恢复,限制了对该套生物成因硅质页岩物性与孔隙系统演化的深入探讨。通过多条剖面筛选选取多个低演化阶段的生物蛋白石硅质岩样品系列,研究早期成岩作用过程中生物蛋白石矿物相变化和与其伴生的硅质页岩孔隙的发育和结构变化过程,并与高演化页岩进行对比,分析早期成岩变化和页岩物性与孔隙系统的协同演化特征,为研究高演化硅质页岩孔隙演化全过程,特别是在中低演化阶段物性和孔隙系统变化以及对储集性能的综合影响提供依据。
1 样品与分析方法
生物蛋白石硅质岩样品采自松辽盆地东北部嫩江地区,层位为下白垩统嫩江组嫩一段。作为对比的五峰组—龙马溪组硅质页岩样品采自四川盆地东南部丁山地区钻井岩心样品。
全岩矿物成分分析在德国布鲁克公司D8 ADVANCE型X 射线衍射仪上完成,在温度25 ℃和相对湿度50%的条件下,依据SY/T 5163-2010标准进行测试。仪器条件:Cu靶,X射线管电压40 kV、电流100 mA,扫描速度4°(2θ)/min,扫描步宽0.02°(2θ)。样品处理是在无污染条件下将页岩样品破碎至200目,取5g左右混合均匀的样品粉末,采用正装法制样并进行X 射线衍射分析,全岩矿物的定量分析在Rockquan 2012软件平台完成。
氦气孔隙度在S-Pore 200型基质孔隙度测定仪上进行,压力传感器精度0.089 kPa,样品室体积为50 cm3,孔隙最大压力0.2 kPa,孔隙度测量范围0~40%。利用氦气测得岩石的骨架体积,通过岩石总体积与骨架体积计算求得孔隙度。钻取直径为2.5 cm,长度为10 cm的柱塞样品,放入烘箱内在105 ℃条件下烘干24 h以上至恒重,记录质量。测试前先进行样品室和参比室体积标定,然后将柱塞样品放入样品室,通入氦气后压力平衡时间在30 min以上。
氮气等温吸附在精微公司JWBK-200C上进行,吸附质为N2。岩石样品预先在脱气站中进行脱气预处理,脱气在150 ℃的N2环境中进行,平衡压力智能控制法,吸脱附孔径分布分析六段优化设置,可设压力间隔范围0.1~133 kPa,可灵活设置,全自动调节。比表面积由Brunauer-Emmett-Teller (BET)方程计算得出,总孔体积为P/P0=0.995的氮气吸附量转换为液氮的体积。孔隙分布特征采用密度泛函理论方法(QSDFT法)进行计算。仪器的精度:比表面积重复精度≤±1.0%,微孔孔径偏差≤0.02 nm。
高压压汞在麦克公司Autopore IV 9520型压汞仪上进行,进汞压力范围0.003~413 MPa,进汞/退汞体积测量精度为0.1 μL,孔径测量范围10 nm~1 000 μm。首先将页岩样品敲碎至3~5 cm左右碎块,放入烘箱内在110 ℃条件下烘干24 h以上。取出样品稍作冷却,将样品放置压汞专用样品杯中,杯口密封后置于压汞仪上进行分析,初始抽真空条件为10-6kPa。
2 结果与讨论
2.1 生物蛋白石相变与硅质页岩早期成岩演化
X射线衍射分析的结果显示,不同剖面的硅质页岩均主要由硅质矿物组成,其他矿物含量较低,在10%以下。这些硅质主要为蛋白石-A、蛋白石-CT和石英3种类型,它们的相对含量在各剖面也存在较明显差别。木河剖面、恰塔剖面和嫩江剖面页岩(MH、QT和NJ样品系列)的蛋白石-A含量占总硅质含量的35%以上,蛋白石-CT占55%以上,石英含量在10%以下。金家沟剖面页岩(JJG样品系列)蛋白石-A含量在10%以下,蛋白石-CT在35%~50%之间,石英含量在40%以上。鸟河和姚家村剖面页岩(NH和YJC样品系列)样品石英含量占85%以上,蛋白石-CT在15%以下,不含蛋白石-A(图1)。蛋白石-A和蛋白石-CT的存在指示各剖面页岩均处于较低的成岩演化阶段,但各硅质矿物的不同相对含量则显示它们的演化程度又略有不同。同时,镜质体反射率的测试结果显示它们的Ro在0.3%~0.5%之间,亦表明它们的演化程度较低且稍有差异。
从图1可以清楚地看出生物蛋白石在成岩作用过程中逐渐向晶体石英转化的阶段性过程。首先是蛋白石-A、蛋白石-CT和石英晶体3种物相共存阶段,由于蛋白石-A非常不稳定,略经成岩就开始向蛋白石-CT转化,这一阶段大部分蛋白石-A已转化为蛋白石-CT,仅有少量蛋白石-A残余,同时含有极少量的石英晶体;随着成岩的继续进行,即进入蛋白石-CT和石英晶体2种物相共存阶段,该阶段蛋白石-A已完全转化为蛋白石-CT,少部分蛋白石-CT转化为石英晶体,但仍以蛋白石-CT为主体;第三阶段则为石英晶体单一物相存在阶段,蛋白石-CT全部演化为晶体石英。事实上,在硅质生物骨架进入沉积物埋藏的初始阶段生物蛋白石全部以蛋白石-A形式存在,并经演化进入蛋白石-A和蛋白石-CT两种物相共存阶段[16],由于本文所研究的样品已经历了一定的演化,所处的是紧随其后的3个演化阶段,它们结合在一起就构成了一套完整的生物蛋白石成岩演化序列。
蛋白石-A是一种高度无序,近乎为无定形结构的含水的非晶质硅质物质,其结构式为SiO2·nH2O。蛋白石-CT 则是由低温鳞石英和低温方石英2种结构畴成一维堆垛无序形态所构成的准晶态硅质矿物[17]。当进入沉积地层后,随着地层温度、压力的增加或沉积年龄的增长,生物蛋白石的溶解度和含水率逐渐降低,密度逐步增大,从不稳定的非晶体结构相蛋白石-A 逐步向化学性质相似但结构更为有序、稳定程度更高的矿物相蛋白石-CT 转化,再经进一步的脱水、结晶,最终转化为具有晶体结构的石英矿物。化学动力学分析表明,层状硅质岩由硅质软泥在成岩阶段重结晶而成,主要受控于温度、时间和埋深等,通常情况下温度起主导作用。在较年轻的沉积地层中蛋白石-A向蛋白石-CT发生转换反应发生的温度在30~50 ℃之间(对应埋深数百米),而在古老地层中则可发生在10~20 ℃范围内[18-20]。在某些沉积环境下,该转化温度还会降低,甚至在4 ℃之下也可以发生[21]。蛋白石-CT向石英的转化温度则较高,大多在60~80 ℃之间[22-23]。蛋白石-CT向燧石的转变速率与温度密切相关,温度越高演化越快。BLATT等[24]的实验研究表明,在100 ℃时,该转变需要40 ka,而在50℃时,这种固态相变就要约4Ma 的时间。由于蛋白石-A和蛋白石-CT的稳定性都不高,成岩转化的温度和压力条件相对较低,成岩较为迅速,因此在进入沉积地层后不久就开始发生蛋白石-A向蛋白石-CT的转化,在早期成岩阶段即开始通过脱水和重结晶作用逐步向晶态的石英转化,转化完全后就基本上完成了硅质页岩的成岩定型,后期的成岩改造作用较弱。
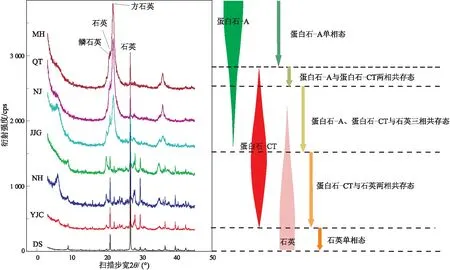
图1 生物蛋白石硅质页岩X射线衍射图谱
2.2 蛋白石成岩演化对页岩总孔隙的影响
硅质生物主要营浮游生活,体内多孔,如放射虫仅由几支较大的硅质骨骼构成生物骨架,骨架内部则由中空的囊体组成,囊体表面和囊内均发育小孔和空腔[25-26],海绵骨针则发育中空的体腔,它们死亡后沉降进入沉积物,使整套沉积地层具有疏松、多孔特征,原生孔隙度较高,最高可达80%以上[27]。随着埋深和温度的增加,生物成因硅质的主要矿物组分—蛋白石-A从不稳定的矿物相逐步发生相变转化为更加稳定的矿物相,该相变过程硅质页岩的物性与孔隙结构及岩石物理性质也随之发生变化。从表1可以看出在蛋白石-A残留阶段,硅质页岩孔隙度在45%以上,甚至高达55%,孔容降最高达0.289 mL/g,比表面积最大达93.89 cm2/g。在蛋白石-A完全转化为蛋白石-CT阶段,页岩的孔隙度降低至29%~45%之间,孔容降至0.076~0.098 mL/g,比表面积降至46.55~68.42 cm2/g。前人的研究结果显示这一阶段孔隙度值可降低至40%以下[28-29],损失幅度较大,与本文研究结果基本一致。在石英单一物相存在阶段,页岩孔隙度进一步降低至25%~29%,孔容降至0.040~0.063 mL/g,比表面积32.73~55.43 cm2/g,孔隙损失幅度明显降低。上述变化说明,在成岩作用下生物蛋白石发生较快速的物相变化,相变过程对硅质页岩的孔隙结构及岩石物理性质等影响较大,生物硅质页岩的孔隙度和孔容迅速降低,比表面积迅速减小,页岩结构从疏松状态通过压实固结趋于致密,同时由于石英的硬度较高使页岩的硬度也随之增大,抗应力改造能力增强。
四川盆地现今高演化的五峰组—龙马溪组底部硅质页岩中的硅质矿物均以晶体石英形式存在[7],孔隙度多在4%~8%(表1)。与低演化生物硅质页岩孔隙度变化进行对比发现,从沉积物开始到蛋白石-A转化为蛋白石-CT阶段的孔隙度损失幅度较大,损失率在35%~50%以上,但从蛋白石-CT转化为晶体石英阶段孔隙度损失幅度明显减小,损失率在5%~15%,再到后期高演化阶段这一漫长演化过程孔隙度损失也极为缓慢(表1),损失率在15%左右。由于所分析高演化硅质页岩中的孔隙主要是充注于孔隙中的原油裂解后在原位形成的有机质孔隙[30-32],而并非所有的沥青均发育孔隙,也就是说仍有部分原油裂解形成的沥青占据着无机孔隙的空间,因此该演化阶段硅质页岩原生孔隙度的损失量应在15%以下,说明一旦生物蛋白石-A完成向准晶体蛋白石-CT和晶体石英的重结晶转化后,后期发生的成岩作用对硅质页岩原生孔隙的破坏程度就变得较为有限。
诸多研究发现,富含蛋白石的沉积地层在埋藏过程中会保持异常高的孔隙度,这是由于硅质生物骨壳会形成坚硬且开放的基质,具有较强的抵御机械压实作用的能力[33-34]。VOLPI等[27]证实,富含蛋白石沉积层在500 m的埋藏深度下孔隙度仍可保持在75%左右。随着蛋白石经重结晶作用向石英晶体不断转化,硅质矿物的硬度不断增强,由大量石英颗粒组成的硅质页岩的硬度持续增强,整体上构成一个刚性的格架,从而形成了有效支撑系统,使得生物成因硅质页岩的抗压实能力大大提高,从而使大量原生孔隙得以良好保存,这为后期液态烃类的原位储集提供了重要储集空间。当演化程度继续增高时,充注其内的液态烃开始裂解进而形成有机孔隙,这时生物成因硅质已完成向石英晶体的转变,页岩抗压实能力增强,非常有利于塑性有机孔隙的保存,并使其在晚期强烈的应力挤压下仍能够有效保持。生物成因硅质页岩早期成岩阶段机械压实和化学压实(压溶)几乎同步进行,对页岩物性改造作用强,虽然使相当一部分的页岩孔隙损失和减少,但同时使页岩硬度增大,支撑和抗压实能力增强,降低了成岩中后期原生孔隙的损失率,并对次生有机孔隙提供了有效的保护作用。生物成因硅质页岩在早期成岩过程中快速成岩定型的特征是硅质页岩在成岩中后期和晚期仍能保持高孔隙的重要原因。

表1 蛋白石硅质岩的孔隙度、孔容和比表面积变化特征
2.3 蛋白石成岩演化对硅质页岩孔隙结构的影响
将N2吸附密度泛函理论方法(DFT)的计算结果和高压压汞分析结果进行联合,就得到硅质页岩孔隙体积的全孔径分布(图2),即利用孔隙直径和孔容增量作图,反映的是页岩中不同大小孔隙的孔体积随孔径变化的分布特征。从图3可以看出,对于不同演化程度的硅质页岩,在微孔(<2 nm)、介孔(2~50 nm)和大孔(>50 nm)3个孔隙区间范围内均有明显的孔隙体积峰存在,在0.8~2 nm之间存在孤立不连续的数个孔体积峰,在4~30 nm之间存在一个连续分布的较大孔体积峰,而在50~10 000 nm之间的孔体积峰呈不连续状态,单峰值变化较大。随着演化程度的不断增高,硅质页岩中所有孔隙均出现较大幅度的损失,但微孔保存程度最好,甚至有所增加,可能由部分损失的介孔和大孔转化而来;其次是介孔,尤其是2~30 nm区间的介孔保存程度较好,大孔则在成岩演化过程中损失程度最大。
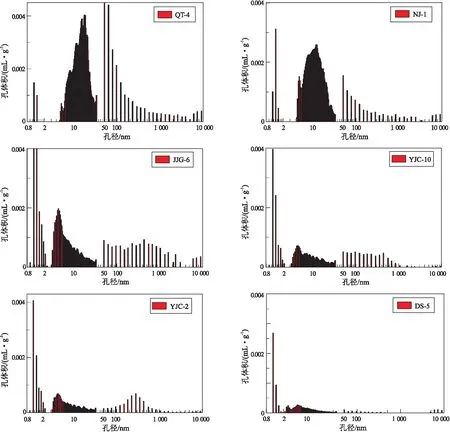
图2 蛋白石硅质页岩与川东南五峰组—龙马溪组硅质页岩孔体积—孔径分布
结合X射线衍射分析所揭示的成岩变化来看,蛋白石-A、蛋白石-CT和石英三相共存态的介孔孔体积最大,其次是大孔和微孔,介孔体积的主峰在18 nm附近,大孔体积主峰在50 nm附近。在蛋白石-CT和石英两相共存态,在介孔整体损失的情况下,10 nm以上介孔的损失尤为明显,介孔体积的主峰发生明显左移,出现在4~5 nm区间,同时大孔也有一定的损失,50~100 nm的大孔较100 nm以上大孔损失程度高,微孔体积有所增加,但孔体积贡献最大的仍然是介孔,其次是大孔。在石英单相态,微孔、介孔和大孔损失幅度减小,总体结构与蛋白石-CT和石英晶体共存态相似。由于用于分析的该阶段的页岩热成熟度已达2.2%左右,其孔隙已由原生的无机孔隙经液态烃充注和裂解在其内形成有机孔隙,孔体积分布总体向小孔径方向变化(图2)。通过上述分析可以看出,在生物蛋白石成岩的物相变化过程中,不同孔径大小的孔隙受成岩改造的程度存在较大的差异性。
2.4 硅质页岩总孔隙和孔径分布演化特征
结合不同演化程度生物成因硅质页岩总孔隙和孔径分布特征,初步建立了生物成因硅质页岩的孔隙和孔径分布演化模式(图3)。硅质页岩孔隙演化主要分为三大阶段:(1)第一大阶段为生物蛋白石成岩物相转化阶段,这一阶段页岩孔隙大幅损失,由80%以上迅速降低至25%左右,降低了约55%。在蛋白石-A向蛋白石-CT转化过程中,页岩总孔隙度先从80%以上快速降低至30%左右,在继续向石英转化的过程中则缓慢降至25%左右,呈先快速和后缓慢的两段式变化。这一阶段作为孔隙主体的大孔和介孔损失十分明显,但微孔却呈略为增加的趋势。(2)第二大阶段为干酪根生烃阶段,这一阶段孔隙继续减少[35],根据四川盆地五峰组—龙马溪组高演化页岩孔隙度范围推断可下降至5%以下,降低了约20%,这一阶段在由成岩作用引起孔隙继续少量损失的同时,随着干酪根热解生烃并在生烃高峰期液态烃占据了页岩内部大部分孔隙而干酪根本身形成的孔隙又少于液态烃占据孔隙的量,导致页岩总孔隙降至最低。此时大部分大孔和介孔被液态烃充注,只剩余部分微孔,总孔隙空间很小。(3)第三大阶段为液态烃热裂解阶段,该阶段液态烃开始裂解生气,大量有机孔隙形成,出现液态烃所占据孔隙的内部扩容现象,使页岩总孔隙逐渐增大,最大可增至8%~10%左右,然后随着成岩与热演化的继续进行,总孔隙又趋于缓慢降低,此时孔隙以介孔和微孔为主,大孔数量较少。
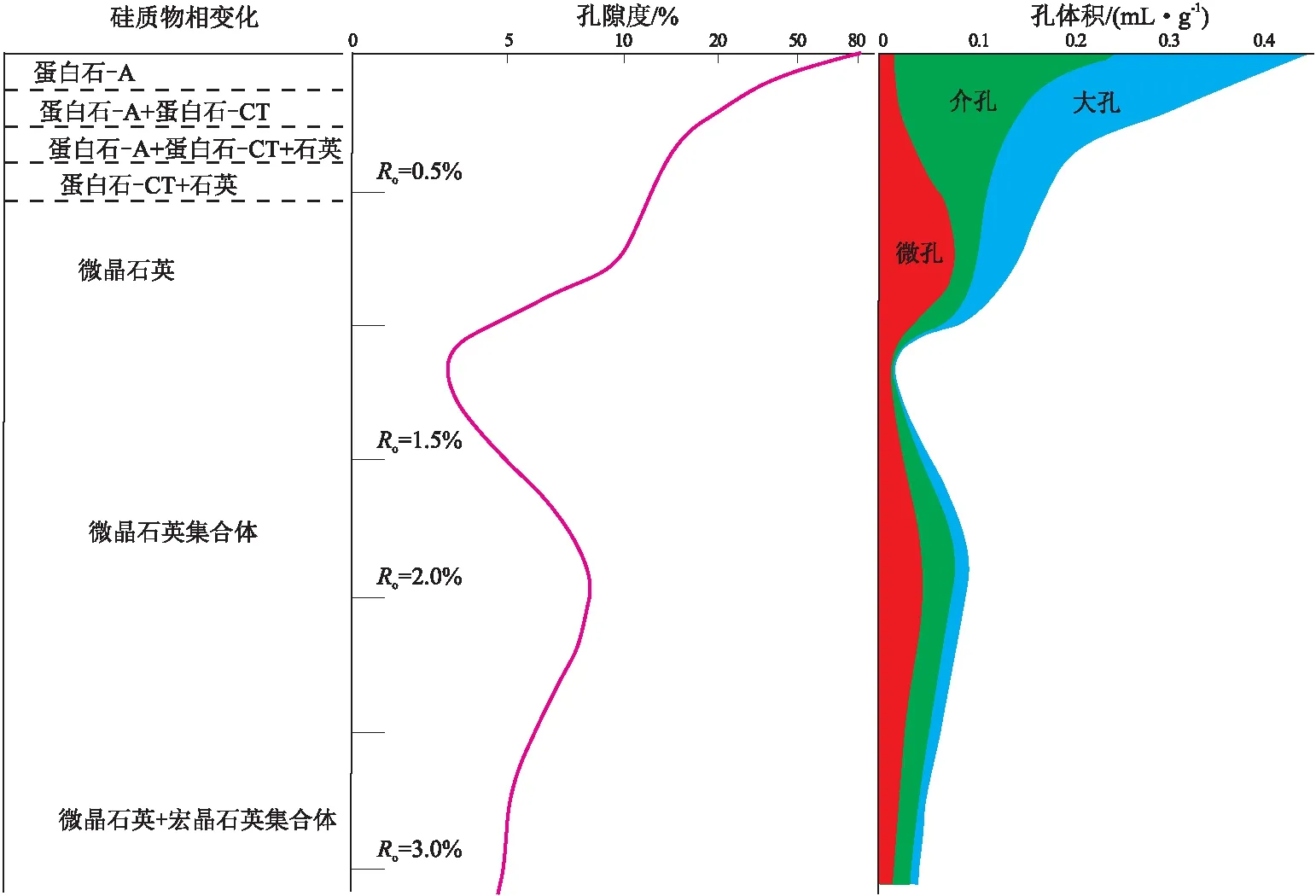
图3 成岩过程生物蛋白石物相转化序列与页岩孔隙演化特征
3 结论
(1)生物成因硅质矿物以非晶态的蛋白石形式存在,随着成岩作用进行逐步向稳定的晶态矿物相转变。生物蛋白石-A发生脱水和重结晶作用较早,在成岩作用早期即完成了向准晶态蛋白石-CT和晶态石英的转变过程。其成岩演化序列共包括蛋白石单相态、蛋白石-A与蛋白石-CT两相共存态、蛋白石-A、蛋白石-CT与石英三相共存态、蛋白石-CT与石英两相共存态和石英单相态五大矿物相态。
(2)在非晶体蛋白石-A向准晶体蛋白石-CT和晶体石英转化过程中,除页岩物理性质有所改变外,硅质页岩的总孔隙和孔隙结构也发生较明显变化。在蛋白石-A向蛋白石-CT转变过程中,总孔隙从80%以上快速降至30%~45%左右,在蛋白石-CT向石英转化过程中孔隙度缓慢降至25%~30%,降低速率迅速减小,表现出快速降低和慢速降低两段式变化特征。同时前一阶段大孔损失较多,微孔损失较小,后一阶段介孔损失较多,大孔损失较少,微孔则略有增加,应为部分所损失的大孔和介孔转化而来。孔隙组成也从以大孔和介孔为主开始向以介孔和微孔为主转变。
(3)生物蛋白石稳定性低,成岩转化迅速,生物成因硅质页岩在成岩作用早期就完成了成岩定型。生物成因硅质页岩早期成岩阶段机械压实和化学压实(压溶)近乎同步进行,对页岩物性改造作用强,使页岩孔隙减小同时硬度增大,支撑和抗压实能力增强,进而使中后期和晚期成岩作用的改造和破坏变弱。生物成因硅质页岩早期快速成岩定型的特性是其在成岩中后期与晚期仍能够保持高孔隙特征的根本原因。
猜你喜欢
化工管理(2021年20期)2021-08-17
西南石油大学学报(自然科学版)(2021年3期)2021-07-16
陶瓷学报(2020年6期)2021-01-26
世界有色金属(2020年1期)2020-03-26
石油地质与工程(2019年4期)2019-09-10
中成药(2018年10期)2018-10-26
制造技术与机床(2018年10期)2018-10-13
资源节约与环保(2018年2期)2018-02-02
幼儿教育·教育教学版(2017年10期)2017-12-13
三联生活周刊(2016年22期)2016-05-27
