
我国大陆、海南岛及台湾岛野牡丹科植物区系特征比较
2020-06-01 07:58温振英樊晚林周雨琦骆亮彭东辉
江苏农业科学 2020年8期
温振英 樊晚林 周雨琦 骆亮 彭东辉


摘要:野牡丹科植物具有极高的观赏价值和药用价值,开发前景广阔,长期以来对野牡丹科资源分布和生存状况等研究不足,导致学术界对其分类存在较大争议,部分物种生存状况堪忧。为了理清我国野牡丹科植物资源状况,通过分析其区系及分布特征,为其保护利用提供基础的数据。在充分查阅相关资料及野外实地调查的基础上,对我国大陆、海南岛和台湾岛的野牡丹科(Melastomataceae)植物的物种组成、分布区类型及水平垂直分布进行分析。结果表明:(1)我国野牡丹科植物共有24属177种25变种,其中大陆有19属150种24变种,海南岛有14属46种7变种,台湾岛有12属22种1变种;海南岛、台湾岛与大陆野牡丹科植物共有属占其各自总属数的85.7%、75.0%;3个地区共有属有7属,分别是金锦香属(Osbeckia)、野牡丹属(Melastoma)、柏拉木属(Blastus)、肉穗草属(Sarcopyramis)、蜂斗草属(Sonerila)、酸脚杆属(Medinilla)和谷木属(Memecylon);海南岛、台湾岛与大陆共有种的比例分别为64.2%、60.9%,说明2个岛屿与大陆野牡丹科植物种的同源性较高。(2)我国大陆、海南岛、台湾岛的野牡丹科植物的分布区类型分别为5、3、5个,均以热带亚洲分布型属的比例最高。(3)3个地区的野牡丹科植物的水平分布从南到北呈现递减的趋势,垂直分布均主要分布在海拔100~2 000 m的区间内。
关键词:野牡丹科;大陆;海南岛;台湾岛;区系特征;分布特征
中图分类号: S685.110.24文献标志码: A
文章编号:1002-1302(2020)08-0153-06
收稿日期:2018-09-17
基金项目:福建省自然科学基金(编号:2015J01085)。
作者简介:温振英(1993—),女,山东曹县人,硕士研究生,主要从事野牡丹科植物的系统分类研究。E-mail:1317014985@qq.com。
通信作者:彭东辉,博士,教授,主要从事园林植物研究。E-mail:fjpdh@126.com。
广义的野牡丹科(Melastomataceae)包括谷木亚科在内,现约有156~166属,4 500种左右,其中野牡丹亚科150~160属,4 050种,谷木亚科约6属450种,主要集中分布于世界的热带和亚热带地区,以热带美洲最多。该科植物具有极高的观赏价值[1-5]和良好的药用价值[6-9],但目前对该科植物的研究主要集中在种质资源调查与育种[10-12]、繁殖栽培[13-14]、系统分类[15-17]、传粉生物学[18-19]等方面。长期以来学术界对于野牡丹科植物的系统分类学界争论不休,《中国植物志》对野牡丹科植物进行了详细研究,记录我国野牡丹科植物有25属约160种25变种[20],而《Flora of China》对《中国植物志》进行了归并与处理,认为我国野牡丹科植物有21属114种。随着对该类群关注越来越多,近年来不断有新种和分布新纪录出现,分类学也发生诸多修订[21-30]。对于目前我国野牡丹科植物种类、区系特征以及分布未见系统报道。
海南岛和台湾岛是我国的2大岛屿,也是野牡丹科植物资源较为丰富的地区,它们的植物区系与大陆,尤其是与东南沿海大陆植物区系有着极为相近的亲缘关系[31-32],比较这2个岛屿与大陆野牡丹科植物区系的关系对于揭示我国野牡丹科的地理起源、迁移、演化等具有重要意义。基于此,本研究在野外实地调查的基础上,结合文献和标本查阅鉴定,对我国大陆、海南岛和台湾岛的野牡丹科植物资源进行了整理,并对其区系进行了分析,以期为今后野牡丹科植物的保护与利用提供基础数据。
1 研究区自然概况和研究方法
1.1 研究区自然概况
野牡丹科植物在我国大陆的分布地区主要位于秦岭-淮河以南,青藏高原以东,东临黄海、东海,南临南海。地理坐标为20°~34°N、100°~120°E。地形包括长江中下游平原、东南丘陵、四川盆地、云贵高原;气候主要是亚热带、热带季风气候;植被类型主要是亚热带常绿阔叶林和热带季雨林。
海南岛处于热带,地理坐标为18°10′~20°10′N、108°37′~111°03′E,面积3.4万km2;最高峰五指山位于海南岛中部,海拔1 867 m[10,12]。土壤类型以黄壤和红壤为主,降雨类型以锋面雨和台风雨为主;植被类型为热带雨林或季雨林。
台湾岛处于热带和亚热带的交接地带,地理坐标为21°45′~25°56′N、119°18′~124°34′E,面积3.6万km2,山地约占全岛面积的2/3;主峰玉山位于台湾岛中部,海拔 3 997 m[31-32];土壤类型以黄壤和红壤为主,台湾岛以北回归线为界,北部属副热带季风气候,南部则为热带季风气候[33-34]。夏季高温多雨、冬季则温暖湿润,岛屿的海洋性气候较明显,雨量充沛,岛上水平和垂直方向的生境类型多样。
1.2 研究方法
以相关植物志书[20,35-46]为基础资料,结合近年发表的相关文献[21-30],开展了原产地的实地调查验证和补充,初步整理出我国大陆、海南岛和台湾岛野牡丹科植物名录。根据3个地区的野牡丹科植物名录对各自野牡丹科植物的物种组成(亚科、属和种的组成)进行比较分析。
根据吴征镒等对我国种子植物分布区类型的划分标准,对我国大陆、海南岛和台湾岛野牡丹科植物属的分布区类型进行整理和统计,分析3个地区野牡丹科植物属的区系特征[47]。
根据热温比和相似系数定量分析3个地区植物区系的差异性和组成特点,计算公式为热温比=R/T,式中:R代表熱带分布属数;T代表温带分布属数。
采用Sprensen相似性公式:S=2c/(a+b)计算两地之间的相似性系数,c为A地与B地的公共属或公共种,a为A地的所有属数或种数,b为B地的所有属数或种数[48]。
根据野牡丹科植物分布的海拔范围,同时考虑到统计样本数量的需要和实际分布范围,除第1个按100 m划分外,其余按海拔间隔500 m均匀划分。
2 结果和分析
2.1 我国大陆、海南岛及台湾岛野牡丹科植物的物种组成及分布特征比较分析
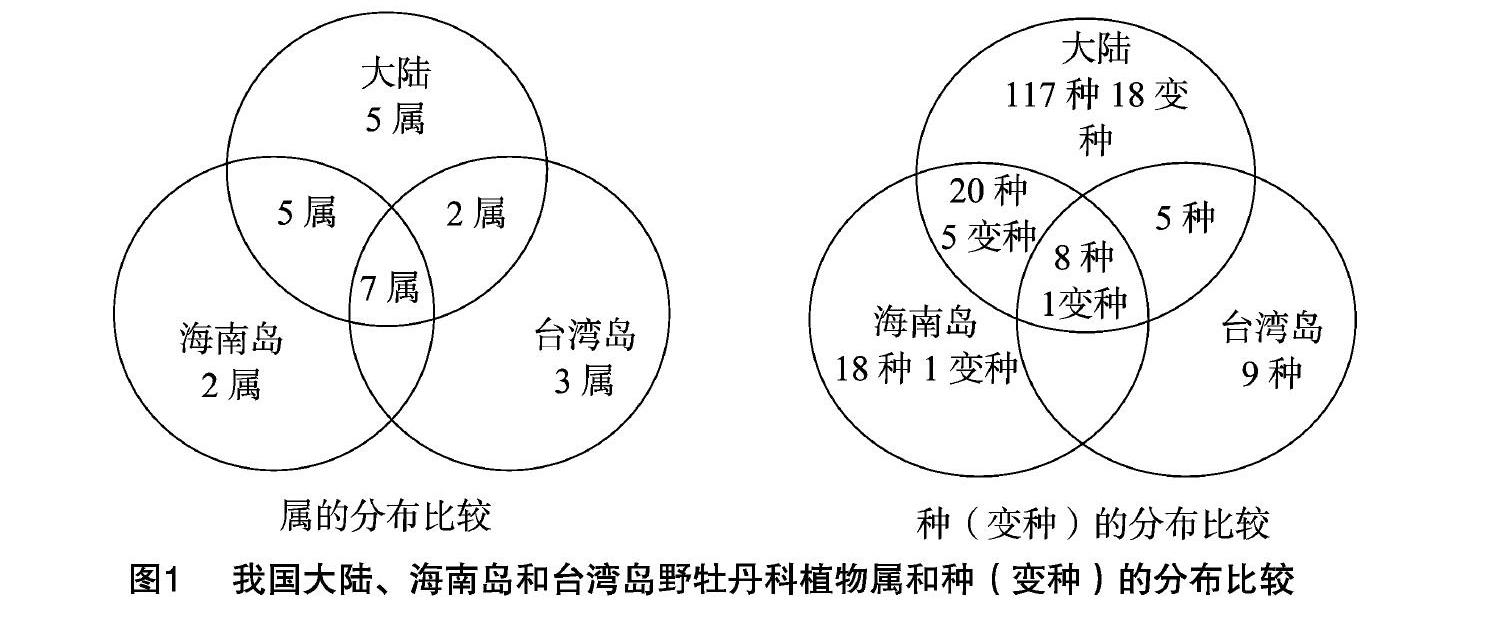
我国大陆、海南岛和台湾岛野牡丹科植物的属和种的数量统计结果见表1。我国大陆、海南岛和台湾岛的野牡丹科植物总计24属177种25变种,其中大陆有19属150种24变种,海南岛有14属46种7变种,台湾岛有12属22种1变种。单种属以海南岛和台湾岛居多,分别为6属和8属,分别占各自总属数的42.9%、66.7%;大陆分布的单种属有5属,占其总属数的26.3%;3个地区无共同分布的单种属。含2~5种的属在我国大陆、海南岛和台湾岛分别有4属、5属和4属,该类属的种数(变种数)分别有14种5变种、20种4变种、14种。6~10种的属只在我国大陆和海南岛有分布,分别有2属和3属,我国大陆和海南岛共同分布的该类属是野牡丹属(Melastoma),该类属的种数(变种数)在我国大陆和海南岛分别有13种1变种和21种2变种。含11种及以上的属仅在中国大陆有分布,共有8属118种17变种,占其总属数的42.1%,总种数和变种数的77.6%。
我国大陆、海南岛和台湾岛野牡丹科植物在3个地区属的分布比较见图1。结果表明,3个地区的共有属有7个,分别占我国大陆、海南岛和台湾岛的野牡丹科植物总属数的36.8%、50.0%和58.3%,占3个地区野牡丹科植物总属数的29.2%。在3个地区分布的野牡丹科植物中,有2个属为海南岛特有,即藤牡丹属(Diplectria)和翼药花属(Pternandra);有3个属为台湾岛特有,分别是耳药花属(Otanthera)、厚距花属(Pachycentria)及褐鳞木属(Astronia)。在大陆有分布但在海南岛和台湾岛无分布的野牡丹科植物有6属,分别是长穗花属(Styrophyton)、尖子木属(Oxyspora)、药囊花属(Cyphotheca)、八蕊花属(Sporoxeia)、虎颜花属(Tigridiopalma)、卷花丹属(Scorpiothyrsus)。在大陆和海南岛有分布但在台湾岛无分布的有5属,分别是异形木属(Allomorphia)、偏瓣花属(Plagiopetalum)、锦香草属(Phyllagathis)、异药花属(Fordiophyton)、卷花丹属(Scorpiothyrsus)。在大陆和台湾岛有分布但在海南岛无分布的有2属,分别是棱果花属(Barthea)、野海棠属(Bredia)。大陆与海南岛和台湾岛属的相似性系数分别为30.3%和12.9%。我国大陆、海南岛和台湾岛的野牡丹科植物在3个地区种(变种)的分布见图1。结果表明,3个地区总计177种25变种,3个地区共有8种1变种,占3个地区野牡丹科植物总种数的4.5%。在中国大陆有分布而海南岛和台湾岛无分布的有117种18变种;在海南岛有分布而中国大陆和台湾岛无分布的有18种1变种;在台湾岛有分布而中国大陆和海南岛无分布的有9种;中国大陆和海南岛共有而台湾岛没有的有20种5变种;中国大陆和台湾岛共有而海南岛没有的有5种。中国大陆与海南岛和台湾岛种的相似性系数分别为35.2%和15.3%。
总的来看,我国大陆野牡丹科植物的多样性更高,种类更多,其中以云南、广西、广东和海南岛分布最多,其次是福建、湖南、江西、贵州、西藏和台湾;四川、浙江分布的属数和种类都较少;湖北、安徽、江苏都有很少的广布种分布;但是海南岛和台湾岛却有在大陆无分布的特有属和特有种,同时,中国野牡丹科植物的分布随着纬度的升高,种类依次减少。此外,3个地区分布的野牡丹科植物均以野牡丹亚科(Melastomatoideae Ser. ex DC.)为最多;谷木亚科(Memecyloideae Naud.)均较少且只有谷木属1属,台湾岛谷木属植物仅有狭叶谷木1种;褐鳞木亚科(Astronioideae Naud.)在大陆无分布,在海南岛和台湾岛分别有1属1种,分别是翼药花属的翼药花(P.caerulescens)和褐鳞木属的褐鳞木(A. ferruginea)。
2.2 我国大陆、海南岛及台湾岛野牡丹科植物属的分布区类型分析
我国大陆、海南岛和台湾岛分布的野牡丹科植物属的分布区类型的统计结果见表2。结果显示,3个地区的野牡丹科植物可分别划分为5、3、5个分布区类型;大陆和台湾岛分布区类型相同,而海南岛,缺少东亚分布和中国特有分布类型。
我国大陆的野牡丹科植物以热带分布型(旧世界热带分布型、热带亚洲至热带澳大利亚分布型、热带亚洲分布型)占绝对优势,共有14属,占其总属数的73.7%;温带成分(东亚分布型和中国特有分布型)仅有5属,占其总属数的26.3%,其中,中国特有分布型的属数有4属,分别是長穗花属(Styrophyton)、药囊花属(Cyphotheca)、棱果花属(Barthea)和虎颜花属(Tigridiopalma),占其总属数的21.1%。热带成分与温带成分的属数量比为2.8(>1),说明我国大陆野牡丹科植物的分布以热带成分为主,温带成分为辅。
海南岛分布的野牡丹科植物中,分布型全为热带成分,与大陆和台湾岛相比,缺少东亚分布和中国特有分布类型。
台湾岛野牡丹科植物的分布区类型与大陆相同,既有热带成分也有温带成分。其中,热带分布型有10属,占其总属数的83.4%;温带成分(东亚分布型和中国特有分布型)仅有2属,占其总属数的16.6%。热带成分与温带成分的属数量比为5,明显大于1,说明台湾岛分布的野牡丹科植物以热带成分为主,有绝对优势。
2.3 我国大陆、海南岛及台湾岛野牡丹科植物的垂直分布
对我国大陆、海南岛和台湾岛分布的野牡丹科植物的垂直分布进行整理结果见图2。野牡丹科植物在3个地区从低海拔至高海拔均有分布,且整体上均呈现先升后降的趋势。3个地区的野牡丹科植物都集中分布在100~<2 000 m的中低海拔地区,高海拔地区相对较少, 这与野牡丹科植物自身对生境的要求有关。同时,我国大陆的野牡丹科植物在中海拔的分布有明显的差距;而我国海南岛和台湾岛的野牡丹科植物在中海拔很大的梯度范围内种类都是相当的,这一方面说明大陆本身的种类多,适应各种生境的种类也多,也说明2个岛屿特殊的地理位置与气候条件满足了这一类植物的生长。
3 讨论与结论
我国大陆、海南岛及台湾岛的野牡丹科植物种类丰富,经整理3个地区总计24属177种25变种,与《中国植物志》记录的25属约160种25变种[20]和《Flora of China》记录的21属114种不同;其中大陆有19属150种24变种,海南岛有14属46种7变种,台湾岛有12属22种1变种。在亚科的组成上,我国野牡丹科植物分为3个亚科,分别是野牡丹亚科、褐鳞木亚科和谷木亚科;中国大陆无褐鳞木亚科的分布,海南岛和台湾岛则各分布有1属1种,分别是翼药花属翼药花(P.caerulescens)和褐鳞木属褐鳞木(A. ferruginea);谷木亚科只有谷木属1属,在中国大陆和海南岛分别分布有11种1变种和8种,而台湾岛则只有1种狭叶谷木(Memecylon);说明3个地区的野牡丹科植物都有其特殊的地方。林有润对广东、海南2个省的野牡丹科植物的系统演化和区系地理进行了深入的探讨,认为野牡丹科种属地理隔离与分布区具有狭窄性特点,因此在南方省份有许多特有或狭域分布的属[49];同时植物是否在一个地区有分布,受自然原因和环境干扰等多方面因素影响[50]。褐鳞木亚科主要分布于东南亚,而在我国只在2个岛屿有分布,说明这2个岛屿与东南亚的气候条件比较相似;狭叶谷木在菲律宾和印度尼西亚也有。
3个地区的区系特征:大陆和海南岛共有而台湾岛无分布的属有5属,大陆和台湾岛共有而海南岛无分布的属有2属;大陆和海南岛共有而台湾岛无分布的种有20种5变种;大陆和台湾岛共有而海南岛无分布的种有5种;说明海南岛与大陆的共有性更高。此外,特有属对许多生物多样性的保护理论和实践都具有重要的指导意义,如许多特有属的植物经历了第四纪冰期并持续生存下来,它们包含了植物系统发育和演化的重要信息[51];特有种是一个地区植物区系和植被的最重要特征,对于该地区的植物研究具有特殊意义[52]。在海南岛分布的野牡丹科植物中,特有种数量为18种1变种,占该岛总种数和变种数的35.8%;在台湾岛分布的野牡丹科植物中,特有种数量有9种,占该岛总种数和变种数的 39.1%;表明海南岛野牡丹科植物区系成分的特有性低于台湾岛。因此,建议对2个岛屿的野牡丹科植物的特有种及其产生形式进行深入比较研究,这将有助于了解野牡丹科植物的起源和扩散形式等。
我国大陆、海南岛和台湾岛的野牡丹科植物区系成分并不复杂,分别有5、3、5个分布区类型,并且主要分为热带成分和温带成分2大类。田怀珍等认为,热带成分与温带成分属的数量比能够大体反映植物区系的性质,比值越大则热带性质越强,反之则温带性质越强[53]。海南岛分布的野牡丹科植物全部为热带成分;大陆和台湾岛分布的野牡丹科植物属的数量比均大于1,且台湾岛野牡丹科植物热带成分与温带成分属的数量比明显高于中国大陆,说明这2个地区分布的野牡丹科植物也均以热带成分为主,但大陆野牡丹科植物的热带性质小于台湾岛并表现出一定的温带性质;2个岛屿的热带成分均较明显,这与2个岛屿的地理位置和气候相对应(台湾岛以北回归线为界,北部属副热带季风气候,南部为热带季风气候;而海南岛主要为热带季风气候[31])。
对我国大陆、海南岛和台湾岛的野牡丹科植物各种类的分布范围的分析结果显示:我国野牡丹科植物的分布呈现出从南到北递减的趋势,越靠近南部热带地区种类越多,随着纬度的升高,种类依次减少。从垂直分布来看,野牡丹科的分布整体上呈现先升后降的趋势,有的种类分布范围较广,从低海拔到高海拔都有分布,而有的种类只在高海拔地区有分布,这与植物不同的习性有关。比如野牡丹属的地菍(Melastoma dodecandrum)、细叶野牡丹(M. intermedium)、展毛野牡丹(M. normale)、多花野牡丹(M. affine)垂直分布范围很广,从平地至海拔1 200 m左右均有分布,展毛野牡丹还可分布到海拔 2 800 m 地带。柏拉木属的柏拉木(Blastus cochinchinensis),酸脚杆属的北酸脚杆(Medinilla septentrionalis),谷木属的谷木(M. ligustrifolium),金锦香属金锦香(Osbeckia chinensis)、锦香草属的叶底红(Phyllagathis fordii),蜂斗草属的小蜂斗草(Sonerila laeta)等垂直分布也比较广,从低海拔到高海拔地带均有分布。多数野牡丹种类垂直分布范围很窄,仅分布于一定的海拔之内,如野牡丹属野牡丹(M. malabathricum),仅分布在海拔120 m以下的山坡林下或开阔的灌草丛中,而细叶谷木(M. scutellatum)仅在海拔约3 000 m之处可见。这与野牡丹科植物具有一定的气温需求有关,在海拔较高地区,因气温较低满足不了野牡丹科植物对温度的要求,所以野牡丹科植物种类分布较少,而较低海拔较少野牡丹科植物分布可能与土地利用格局、人为破坏较多有关。Rahbek发现,很多地域物种丰富度并不随着海拔的上升而降低,而是在中间海拔物种丰富度最高,即中域效应理论[54]。
综上所述,本研究的结论如下:(1)我国野牡丹科植物共有24属177种25变种,其中大陆有19属150种24变种,海南岛有14属46种7变种,台湾岛有12属22种1变种;海南岛、台湾岛与大陆野牡丹科植物共有属占其各自总属数的85.7%、75.0%,3个地区共有属有7属,分别是金锦香属(Osbeckia)、野牡丹属(Melastoma)、柏拉木属(Blastus)、肉穗草属(Sarcopyramis)、蜂斗草属(Sonerila)、酸脚杆属(Medinilla)和谷木屬(Memecylon);海南岛、台湾岛与我国大陆共有种的比例分别为64.2%、60.9%,说明2个岛屿与中国大陆野牡丹科植物种的同源性较高。(2)我国大陆、海南岛和台湾岛的野牡丹科植物的分布区类型分别可分为5、3、5个,并且均以热带亚洲分布型属的比例最高。(3)3个地区的野牡丹科植物的水平分布从南到北呈现递减的趋势,垂直分布均主要分布在海拔100~2 000 m的区间内。
参考文献:
[1]范建红,冯志坚,向春玲. 广东野牡丹科野生观赏植物资源[J]. 中国野生植物资源,2002,21(4):19-21.
[2]尹俊梅,王祝年,杨光穗,等. 海南野牡丹科野生观赏植物种质资源及其开发利用[J]. 热带农业科学,2006,26(6):63-66.
[3]彭东辉,张启翔,黄俊婷. 中国野牡丹科观赏植物种质资源及其在福建省的分布初步调查[J]. 中国园林,2007,23(11):83-88.
[4]仇 硕,赵 健,张翠萍,等. 中国野牡丹科观赏植物种质资源的研究现状与展望[J]. 安徽农业科学,2008,36(22):9471-9472,9519.
[5]赵 健,仇 硕,李秀娟,等. 广西野牡丹科观赏植物种质资源及开发前景[J]. 河北农业科学,2008,12(7):80-82.
[6]林秀香,苏金强,黄阿凤. 福建野牡丹科植物资源初步调查及评价[J]. 福建热作科技,2003,28(4):17-19.
[7]陈红锋,邢福武,刘东明,等. 广东省野牡丹科药用植物资源[J]. 中药材,2003,26(5):321-323.
[8]常章富,吴嘉瑞,滕云霞,等. 中国野牡丹科药用植物性能主治的研究[J]. 中国中药杂志,2008,33(7):854-859.
[9]田建平,胡远艳. 海南野生野牡丹科药用观赏植物资源的调查[J]. 海南医学院学报,2008,14(1):24-26,31.
[10]代色平,刘连海,刘 慧,等. 广东省野牡丹科植物资源调查与评价[J]. 福建林业科技,2012,39(4):121-126.
[11]江鸣涛. 几种野牡丹属植物杂交育种技术研究[D]. 福州:福建农林大学,2015.
[12]代色平,张继方,贺漫媚,等. 野牡丹属新品种‘天骄和‘心愿[J]. 园艺学报,2016,43(9):1847-1848.
[13]唐淑玲,徐江宇,江鸣涛,等. 毛稔叶片离体培养及植株再生研究[J]. 森林与环境学报,2016,36(1):67-72.
[14]Nasir W M,Ahmad W W,Musa N W. Growth responses of Melastoma malabathricum to elevated carbon dioxide and water regime[J]. American Institude of Physics,2016,1784(1):060052.
[15]Liu T,Chen Y Y,Chao L F,et al. Extensive hybridization and introgression between Melastoma candidum and M. sanguineum[J]. PLoS One,2014,9(5):e96680.
[16]Chao L F,Chen Y Y,Wang S Q,et al. One species or two? Multilocus analysis of nucleotide variation of Melastoma penicillatum and Melastoma sanguineum (Melastomataceae) in Hainan,China[J]. Biochemical Systematics and Ecology,2014,55(2):275-282.
[17]白岳峰. 中国野牡丹属植物系统分类研究[D]. 福州:福建农林大学,2015.
[18]金 红,焦根林,陈 刚. 白花野牡丹的开花进程、花部形态特征及访花昆虫观察[J]. 植物资源与环境学报,2015,24(2):73-79.
[19]彭东辉,兰思仁,吴沙沙. 中国特有种枝毛野牡丹传粉生物学及繁育系统研究[J]. 林业科学研究,2014,27(1):11-16.
[20]中国科学院研究所. 中国植物志:第53卷 第一分册[M]. 北京:科学出版社,1984:135-293.
[21]Zheng Y L,Xia N H. Blastus longiflorus var. apricus,a new combination in Chinese Melastoma-taceae[J]. Journal of Tropical and Subtropical Botany,2003,11(3):275-276.
[22]Yeh C L,Yeh C R. Bredia quadrangularis Cogn. (Melastoma-taceae),a newly recorded plant in Taiwan[J]. Taiwan Journal of Forest Science,2006,21(1):119-123.
[23]Yeh C L,Chung S W,Hsu T C,et al. Two new species of Bredia (Melastomataceae) from Taiwan[J]. Edinburgh Journal of Botany,2008,65(3):393-405.
[24]寧小清,刘寿养. 广西野牡丹科一新种——大明山异药花[J]. 广西植物,2010,30(6):825-826.
[25]兰思仁,彭东辉,张启翔,等. 福建野牡丹科一新纪录种——短葶无距花[J]. 福建农林大学学报(自然科学版),2011,40(5):485-487.
[26]林沁文. 福建蜂斗草属(野牡丹科)一新种[J]. 植物研究,2015,35(6):803-806.
[27]Zeng S J,Zou L H,Wang P,et al. Preliminary phylogeny of Fordiophyton (Melastomataceae),with the description of two new species[J]. Phytotaxa,2016,247(1):45-61.
[28]Tian J,Pen L,Zhou J C,et al. Phyllagathis guidongensis (Melastomataceae),a new species from Hunan,China[J]. Phytotaxa,2016,263(1):58-62.
[29]Zeng S J,Huang G H,Liu Q,et al. Fordiophyton zhuangiae (Melastomataceae),a new species from China based on morphological and molecular evidence[J]. Phytotaxa,2016,282(4):259-266.
[30]Zhao W Y,Fan Q,Ye H G,et al. Bredia changii,a new species of Melastomataceae from Jiangxi,China[J]. Phytotaxa,2017,307(1):36-42.
[31]廖文波,金建华,王伯荪,等. 海南和台湾蕨类植物多样性及其大陆性特征[J]. 西北植物学报,2003,23(7):181-189.
[32]苏缨涛,张华荣,李俊清,等. 海南与台湾中国种子植物特有属多样性研究[J]. 北京林业大学学报,2001,23(4):13-19.
[33]蔡 飞,徐国士. 台湾的植物生物多样性及其特点之探讨[J]. 浙江大学学报(理学版),2002,29(2):184-189.
[34]应俊生,徐国士. 中国台湾种子植物区系的性质、特点及其与大陆植物区系的关系[J]. 植物分类学报,2002,40(1):1-51.
[35]中国科学院昆明植物研究所. 云南植物志[M]. 北京:科学出版社,1979:77-137.
[36]中国科学院华南植物园. 广东植物志[M]. 广州:广东科技出版社,2007:9-42.
[37]中国科学院广西植物研究所. 广西植物志[M]. 南宁:广西科学技术出版社,2005:1-32.
[38]福建省科学技术委员会. 福建植物志[M]. 福州:福建科学技术出版社,1989:106-128.
[39]江西植物志编辑委员会. 江西植物志[M]. 北京:中国科学技术出版社,2004:278-279.
[40]贵州植物志编委会. 贵州植物志[M]. 成都:四川省民族出版社,1989:174-202.
[41]浙江植物志编辑委员会. 浙江植物志[M]. 杭州:浙江科学技术出版社,1993:289-298.
[42]吴征镒. 西藏植物志[M]. 北京:科学出版社,1986:345-354.
[43]Wu Z Y,Raven P H. Flora of China[M]. Beijing:Sciences Press,2009:1-506.
[44]中國科学院华南植物研究所. 海南植物志[M]. 北京:科学出版社,1965:23-41.
[45]陈 介. 中国野牡丹科植物志资料[J]. 植物研究,1984,4(3):33-68.
[46]傅立国,陈潭清,郎楷永,等. 中国高等植物[M]. 青岛:青岛出版社,1998:614-662.
[47]吴征镒,周浙昆,孙 航,等. 种子植物分布区类型及其起源和分化[M]. 昆明:云南科技出版社,2006:1-451.
[48]张镱锂,张雪梅. 植物区系地理研究中的重要参数——相似性系数[J]. 干旱区研究,1998,17(1):59-63.
[49]林有润. 广东、海南两省樟科、野牡丹科及菊科的系统演化与区系地理的热带亲缘[J]. 植物研究,1996,16(3):250-272.
[50]田海芬,刘华民,王 炜,等. 大青山山地植物区系及生物多样性研究[J]. 干旱区资源与环境,2014,28(8):172-177.
[51]李文周. 福建戴云山国家级自然保护区种子植物区系分析[J]. 武夷科学,2017,33(0):106-112.
[52]左家哺,傅德志. 植物区系学中特有现象的研究进展(Ⅰ)——概念、类型、起源及其研究意义[J]. 湖南环境生物职业技术学院学报,2003,9(1):11-20.
[53]田怀珍,董全英,李宏庆. 中国38个地区兰科植物区系成分的比较分析[J]. 西北植物学报,2013,33(4):808-823.
[54]Rahbek C. The role of spatial scale and the perception of large-scale species-richness patterns[J]. Ecology Letters,2005,8(2):224-239.
猜你喜欢
小学生学习指导(中年级)(2018年10期)2018-10-10
知识经济·中国直销(2018年3期)2018-04-12
儿童故事画报·自然探秘(2016年2期)2016-03-15
中学历史教学(2015年7期)2015-11-11
河北渔业(2015年9期)2015-09-15
河北渔业(2015年6期)2015-06-24
雕塑(1999年2期)1999-06-28
军事历史(1994年6期)1994-08-15
军事历史(1987年2期)1987-08-20
军事历史(1981年1期)1981-08-21
