
不同干旱胁迫处理对燕麦产量和叶片超微结构的影响
2020-05-23 06:40:10张志芬付晓峰刘俊青杨海顺何江峰王力伟
北方农业学报 2020年1期
张志芬,付晓峰,刘俊青,杨海顺,何江峰,王力伟
(内蒙古自治区农牧业科学院,内蒙古 呼和浩特 010031)
燕麦(Avena sativa L.)是禾本科燕麦属草本植物,抗逆性强,燕麦根系发达,吸水能力较强,且根冠比较大,调节水分能力较强,成为治理土地荒漠化的先锋作物[1-2]。燕麦是我国北方干旱和半干旱地区的重要作物,叶绿体和线粒体的形态结构在一定程度上作为表征植物对逆境条件耐受性的依据[3-4]。正常水分条件下,叶绿体数目多,被膜清晰,结构完整,基粒和基质类囊体膜结构清晰排列整齐[5]。轻度短时间干旱胁迫叶绿体的形状由椭圆形变为弓形或圆形,脱离细胞壁,向细胞中央靠拢,如果干旱胁迫持续,叶绿体膜受到破坏,基粒片层排列紊乱,类囊体松散、解体淀粉粒增多,且这些变化会随时间的延长而加重[6-7]。重度干旱胁迫下,细胞核中染色质凝聚,线粒体嵴膜几乎被破坏殆尽,线粒体空化[8-10]。目前关于干旱胁迫对燕麦超微结构的影响研究较少,张志芬等[11]仅研究了拔节期干旱胁迫对燕麦叶片超微结构的影响,而生产中燕麦常常处于全生育期干旱,因此,本试验从拔节期进行控水到灌浆期,以灌浆期燕麦叶片为研究对象,比较不同干旱胁迫下燕麦叶片表皮细胞和叶肉细胞超微结构,旨在进一步揭示燕麦在形态结构上的耐旱特性,从而为耐旱燕麦品种的选育提供理论参考。
1 材料和方法
1.1 水分胁迫处理
盆栽试验于2019年8—10月在温室内进行,设置3 个水分处理,分别为正常水分、中度干旱和重度干旱(土壤含水量分别为田间最大持水量的75%、45%、30%),从拔节期开始控水,利用称重法进行水分控制,重复3 次,每次重复种植10 盆。试验选用直径为23 cm、高18 cm 的塑料盆进行,每盆均匀播种燕麦30 株,三叶期定植为20 株。每天6:30—21:30用补光灯进行补光。
1.2 取样
供试燕麦品种为蒙燕1 号。9月26日燕麦处于灌浆期,10:30 进行取样,选择完好无损健康植株,从上而下取第2 叶片,用夹子夹住叶基部分,放在缓冲液中冲洗干净,在离开叶基部0.5 cm 处截取长1.0 cm 叶片,放入装满2.5%戊二醛的离心管中,冷藏备用。
1.3 扫描电镜观察
取出样品,用磷酸缓冲液漂洗,然后取5 mm2叶片,1%锇酸固定2 h,经乙醇系列梯度脱水,临界点干燥,黏合喷金后在S-530 型扫描电镜下观察和拍照。
1.4 测定产量
成熟时,每个处理取10 株进行考种,测定穗长、小穗数、单株粒数、单株粒重、穗粒重、粒重和产量。每个处理10 盆,其中,5 盆用于测定产量、5 盆用于测定干物质量。
1.5 干物质量的测定
分别在拔节期、抽穗期和灌浆期取20 株测定叶片、茎秆和穗的干重,重复3 次。
1.6 数据处理
用SPSS 17.0 进行显著性分析。
2 结果与分析
2.1 干旱胁迫对燕麦产量及其构成因子的影响
由表1 可知,中度和重度干旱胁迫处理主穗长、单株粒数均显著(P<0.01)低于正常水分,但中度和重度干旱胁迫处理之间差异不显著(P>0.01);中度干旱胁迫处理百粒重显著(P<0.01)高于重度干旱胁迫和正常水分处理,说明一定程度的干旱胁迫使得结实率下降,但有利于结实籽粒的灌浆,籽粒更饱满。中度干旱胁迫比正常水分条件减产26.3%,重度干旱胁迫较正常水分条件减产35.3%。
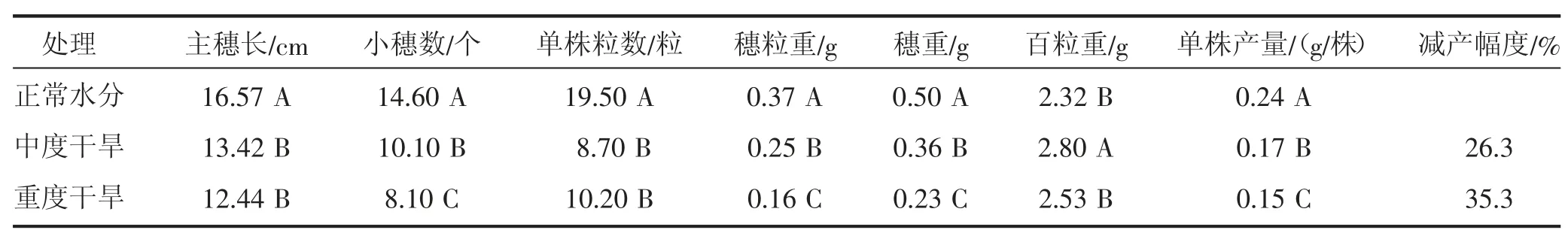
表1 干旱胁迫对燕麦产量及其构成因子的影响
2.2 干旱胁迫对燕麦地上部分干物质量的影响
干旱胁迫对燕麦拔节期、抽穗期和灌浆期茎秆、叶片和穗的干物质量影响处理之间差异显著性见表2。干旱胁迫对干物质量的影响随着干旱胁迫时间的延长而增加,对叶片干物质量的影响大于对茎秆和穗的影响,其中,抽穗期和灌浆期叶片干物质量处理之间达到显著水平(P<0.05),拔节期叶片干物质量处理之间不显著(P>0.05),而茎秆和穗干物质量不同生育时期处理之间差异均不显著(P>0.05)。
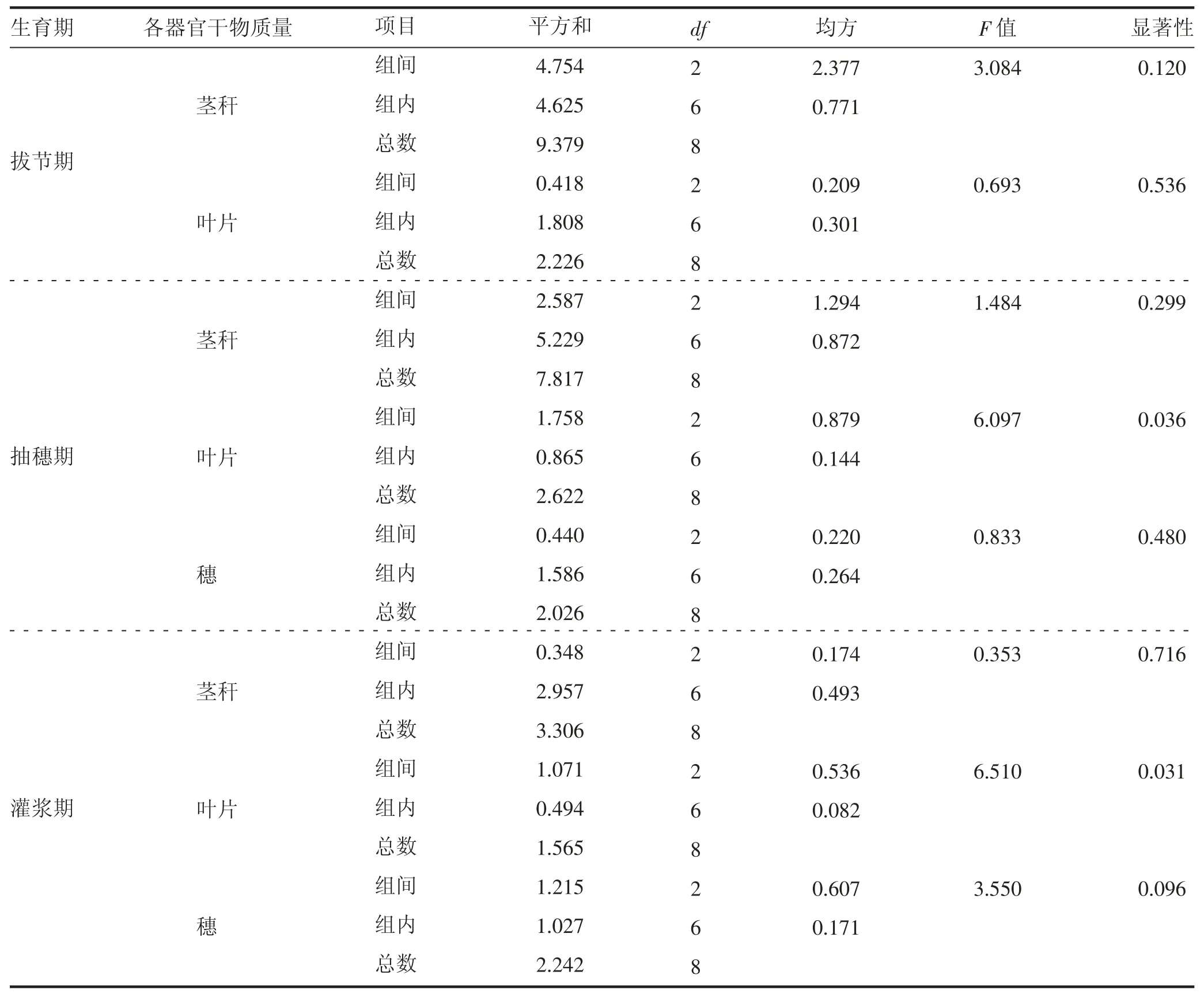
表2 不同生育时期茎秆、叶片和穗干物质量的差异显著性
2.3 干旱胁迫对叶片表皮细胞的影响
干旱胁迫对燕麦叶片上表皮和下表皮的影响如图1 所示,正常水分条件下,叶片上下表皮气孔较多,气孔结构明显,气孔由两个保卫细胞组成;表皮细胞由长细胞和短细胞组成,表皮细胞饱满,长、短细胞顺着主叶脉整齐排列,表皮细胞较长,短细胞较少;上表皮的表皮毛密集,顺着主叶脉整齐排列,形成表皮毛带,下表皮的表皮毛较少。
中度干旱胁迫处理,上表皮分泌物较多,气孔数量减小;上、下表皮均短细胞增多,下表皮细胞干瘪,出现皱褶,较正常水分条件下气孔器不明显。
重度干旱胁迫处理,上表皮的表皮毛较少,表皮细胞干瘪、皱褶,下表皮气孔器分化不明显,在放大150 倍的情况下很难辨认气孔和表皮细胞,除嵴为长细胞以外,大部分表皮为短细胞,而且皱褶严重。因此认为燕麦下表皮的结构对干旱胁迫更为敏感,长时间的重度干旱胁迫导致燕麦气孔器退化、保卫细胞较小。
2.4 干旱胁迫对叶肉细胞超微结构的影响
在正常水分条件下(图2A、图2B),燕麦灌浆期叶肉细胞较大,细胞膜完整,核染色质均匀分布,叶绿体较大,外被膜叶绿体内类囊体基粒较大、数目较多、排列整齐。但在灌浆期部分叶肉细胞叶绿体边缘化。
中度干旱胁迫处理(图2C、图2D),细胞膜断裂,核染色质凝聚,淀粉粒增多,线粒体数目较少、体积变小,叶绿体外被膜完整,内部类囊体基粒数目较正常水分条件下少,部分类囊体基粒结构模糊,线粒体较正常水分条件下开始膨大。
重度干旱胁迫处理(图2E、图2F),叶肉细胞较小,大部分细胞膜断裂,核染色质较中度干旱胁迫处理下更为凝聚,叶绿体出现团聚现象、形态各异、数目较少、叶绿体内有较少完整的类囊体基粒和基质结构,线粒体较中度干旱胁迫处理下体积更大。
3 结论与讨论
干旱胁迫是影响燕麦产量的主要因素,本研究结果表明,随着土壤含水量的下降,燕麦产量显著降低,对叶片干物质量的影响大于对茎秆和穗的影响,但一定程度的干旱胁迫有利于燕麦籽粒的灌浆,籽粒更饱满,这可能是干旱地区燕麦籽粒性状更好的原因。气孔关闭是植物对干旱胁迫的一个普遍反应[12]。已有研究表明,大麦叶片表皮层的气孔器下凹,气孔常常处于关闭状态或开得很小,表现出大麦是一种较耐旱的作物[13]。本研究表明,燕麦表皮层结构与大麦相似,在灌浆期非干旱胁迫下气孔器下凹,常常处于关闭状态或者开的很小,而从拔节期到灌浆期一直处于干旱胁迫下,同样表现为闭合状态,且闭合较紧,但与非胁迫相比气孔器分化不明显。陈温福等[14]研究表明,水稻气孔密度与净光合速率呈明显的正相关。本研究结果与上述研究结果一致,随着干旱胁迫程度增加,气孔密度降低。已有研究表明,燕麦拔节期干旱胁迫7 d,随着干旱胁迫程度的增加,叶片叶绿体首先出现形状改变,然后逐渐内部结构降解,出现较大的空隙;线粒体首先出现膨胀,然后内部结构降解,最终外被膜断裂,内含物外流;重度干旱胁迫下出现细胞器周缘化[15]。本研究表明,在灌浆期即使在非水分胁迫下,细胞器也同样略有周缘化。细胞壁排列的周缘化及形态结构的球状化是其类囊体片层膜结构退化与降解过程启动的“前奏”,对叶片的光合作用也有一定程度的影响。因此认为在灌浆期燕麦倒数第二叶叶片刚刚开始老化,叶绿体的光合作用功能开始降低。另外本研究表明,从拔节期开始控制水分到灌浆期,重度干旱胁迫导致叶绿体形态各异,说明长时间的重度干旱胁迫因叶绿体的形态结构受损严重导致光合作用的下降,但同时叶绿体出现团聚和融合现象,这有助于加强细胞的支撑力,是燕麦对干旱胁迫的适应性的表现,因此认为燕麦对干旱胁迫适应能力较强。
猜你喜欢
中老年保健(2022年5期)2022-11-25 14:16:14
现代畜牧科技(2021年10期)2021-11-19 08:42:24
今日农业(2021年7期)2021-07-28 07:07:24
人民黄河(2020年12期)2020-12-30 12:55:30
山西农业科学(2020年9期)2020-09-14 08:20:08
山西农业科学(2019年12期)2019-12-19 04:03:30
延河(2017年7期)2017-07-19 21:01:10
电子制作(2017年10期)2017-04-18 07:23:14
广西林业科学(2016年1期)2016-03-20 05:33:01
食品科学(2013年6期)2013-03-11 18:20:13
