
西洞庭湖季节性淹水和植被类型对温室气体排放通量的影响*
2020-05-08 03:05:54晁碧霄雷光春王玉玉
湖泊科学 2020年3期
郭 佳,晁碧霄,张 颖,雷光春,王玉玉
(1:北京林业大学生态与自然保护学院,北京 100083) (2:中国农业大学资源与环境学院,北京 100094)
由温室气体引起的全球变暖目前仍是各国政府和科学家关注的重点,CO2、CH4和N2O是影响全球变暖的关键温室气体[1]. 近100年内CH4的全球增温潜势(GWP)大约是CO2的28倍,而N2O的GWP大约是CO2的265倍. 近年来研究表明这3种温室气体对全球气候变暖的贡献率已经超过了90%[2]. 湿地不仅是陆地上巨大的有机碳储库,同时,严格的厌氧环境、巨大的物质与能量循环,使湿地排放大量的CH4和N2O等温室气体[3],其中CH4排放量占全球排放总量的20%~39%,CO2排放量占全球排放总量的5%~8%[4].
已有研究表明,湿地内生长的植被对温室气体的产生、消耗和传输过程有重要影响[4-5]:一方面,植被根系及枯落物的存在为微生物的附着和活动提供了环境和有机物[6],促进了细菌群落组成的多样性,改变了土壤碳含量和pH值[7],从而对温室气体的产生造成影响;另一方面,不同的湿地类型植被,解剖结构和生理学特征各异,对温室气体的运输能力不同[8]. 维管束植物的通气组织向根部运输氧气以维持根的呼吸作用,同时也是植物传输温室气体,特别是CH4的主要通道[9]. 此外,植被类型及植物的生长状况都会对湿地生态系统温室气体的排放产生重要影响[10]:印杰等通过实验研究揭示了有植物群落存在的沉积物中CO2的累积排放量显著高于无植物群落存在的光滩[11];李杨杰通过植被对长江口温室气体排放影响机制的研究表明,不同植被类型在不同的生长阶段对温室气体的源汇功能不一致,长江口海三棱藨草、芦苇以及互花米草这3种主要植被在CH4旳排放过程中均起着重要的促进作用;在CO2的排放中,芦苇表现为汇,海三棱藨草未萌发时表现为源,萌发后表现为汇,等到凋亡后再次表现为源[5]. 湿地植被对于N2O的运输和排放过程同样起着关键作用,植物根系可以吸收其周围的N2O并向上运输[12],最终通过叶片上的气孔将N2O排到空气中[13].
水文条件作为湿地重要的生态属性,水位波动会对湿地大气-植被-土壤之间碳的生物地球化学过程中各环节的方向与强度造成影响[14]. 厌氧环境和有氧环境的转变,使得温室气体的排放量及3种温室气体比例发生较大的变化. 许多研究表明,干燥-再湿润循环强烈影响着土壤的碳氮循环过程[15],微生物的生物量及其活动[16],以及CO2、CH4和N2O等温室气体的排放[17]. Altor 等研究表明,与永久性淹没的地区相比,水位波动使得土壤间歇性暴露在大气中的区域,CH4的排放量显著增高[18];Mander等研究发现,在低水位时,CO2和N2O的排放量更高,而在高水位时,CH4的排放量更高[19]. 水位降低到一定程度,湿地会从CH4的释放源转变为CH4的吸收汇,一定范围内的水位下降会导致N2O排放量增加,而当水位进一步下降时,N2O排放量将减少[20]. 但已有的研究多集中在陆地及滨海地区,而对于长江流域通江湖泊的研究较少.
洞庭湖是我国的第二大淡水湖,也是长江中游仅存的两个通江湖泊之一[21],受自然地理条件、气候条件、江湖关系变化等因素的影响,其水位及消落带面积呈现出周期性变化[22]. 一般4-9月是丰水期,湖水水位偏高,其中7-8月呈现高水位;10月至翌年3月为枯水期,水位较低. 史璇研究发现洞庭湖1961-2008年间,湖滨消落带面积在1336~2920 km2之间波动,多年平均消落带面积为2434.5 km2[23]. 西洞庭湖湿地位于洞庭湖西部,湿地内不仅分布着芦苇、苔草等典型湿地植被,还分布有杨树人工林. 在1990s末期,为了经济利益通过挖沟抬垄在西洞庭湖种植了大量的杨树林,造成了湿地面积萎缩、水鸟栖息地减少等一系列生态问题. 2013年起西洞庭湖开展“退林还湿”工程,开始大量砍伐杨树林并进行湿地生态修复[24],目前,西洞庭湖国家级自然保护区核心区范围内的杨树林已经基本被移除,缓冲区和实验区的杨树林清除工作仍在进行. 近年来由于三峡水库蓄水以及气候变化等原因,西洞庭湖水文条件发生了一定程度上的改变. 综上所述,在进行湿地恢复的过程中研究西洞庭湖湿地在水文条件变化条件下温室气体排放通量的规律,对于西洞庭湖湿地恢复特别是植被恢复具有重要意义.
1 研究区概况与研究方法
1.1 研究区概况
实验地点位于湖南省汉寿县西洞庭湖国家级自然保护区,属于中亚热带季风气候,气候温和,日照充足,雨量充沛,年均气温16.7℃,降水量1200~1350 mm,无霜期274天. 保护区内主要分布有禾本科、莎草科等多种植物群落. 除此之外,由于人为影响,西洞庭湖国家级自然保护区内曾分布有杨树人工林. 截至目前,核心区内的杨树人工林已经被全部移除,正处于湿地恢复阶段.
1.2 研究方法
1.2.1 野外样品采集 通过前期文献阅读和实地考察,选取了芦苇(28°51′07.64″N,112°12′44.92″E)、苔草(28°51′22.04″N,112°13′50.26″E)、刚砍伐的杨树林(28°55′19.37″N,112°11′58.75″E)3个样点. 2018年1月,在以样点为中心的群落内,随机选择3个点,分别采集0~10、10~20、20~30 cm的沉积物样品,装入自封袋里保存,带回实验室分析其理化性质. 用直径为31.5 cm,高为30 cm的PVC管,在每种植被类型的样地上各取16个原状土带回实验站用于模拟实验.
1.2.2 实验设计 将采回的原状土柱静置至植被萌发,其中芦苇样点取回的土柱主要萌发的植被为芦苇(Phragmitesaustralis),苔草和杨树林样点取回的土柱主要萌发的植被为莎草科的灰化苔草(Carexcinerascens) (在后文描述中简称为芦苇湿地、苔草湿地和杨树林湿地),每种植被类型中选取生物量相近的3个原状土柱进行试验,每组3个重复. 将原状土柱上接0.5 m的PVC管后,按照洞庭湖水文变化节律进行淹水(7月5日)和退水(10月1日)处理. 淹水高度设置为0.4 m,淹水时,苔草平均高度约0.6 m,芦苇平均高度约为1.1 m. 2018年5月-2019年1月对原状土柱温室气体排放通量进行监测,频率为1次/月,4~7天/次.
1.2.3 气体采集及分析 温室气体监测采用静态暗箱-气相色谱法. 为了方便淹水和采样,在淹水前将原状土柱上接0.5 m PVC材质的延长箱,淹水高度为0.4 m,留下高度为0.1 m的气室用于采集气体样品. 每次采样前盖上PVC材质的盖子,并用封口膜密封;盖子内部装有一个小风扇,保证在采样时,箱内气体混合均匀;并且盖子上装有橡胶管连接的出气口. 釆集气体样品时,用带有三通阀的医用100 mL注射器连接采样箱出气口和气袋,把盖子盖上后立刻采集第一袋气体样品,取箱内气体后把气体样品注入到容积为100 mL的气袋中储存. 之后每隔10 min完成一袋气体样品的采样,每个采样点共采集4袋气体样品,运到实验室进行分析. 采集样品的同时,记录箱内温度. 采样后,记录每个采样箱的气室高度,用于温室气体排放通量的计算.
1.3 数据计算及统计分析
1.3.1 数据计算 采用公式(1)计算温室气体排放通量:
(1)
式中,F为温室气体(CO2、CH4和N2O)排放通量(mg/(m2·h)),M为气体摩尔质量(g/mol),V0为标准状态下气体的摩尔体积(V0=22.4 L/mol),P为采样时刻大气压(kPa),P0为标准大气压(P0=101.325 kPa),T0为标准状态下的绝对温度(T0=237.15 K),T为采样时刻温度(K),dCt/dt为箱内气体浓度变化速率,H为气室高度(m).
采用公式(2)计算气体累积排放量:
(2)
式中,CE(cumulative emission)表示温室气体(CO2、CH4和N2O)的累积排放量(g/m2);F为温室气体排放通量(mg/(m2·h));i表示第i次气体采样;ti+1-ti表示两个相邻测定日期的间隔(d);n为通量观测次数.
采用公式(3)计算综合增温潜势:
GWP=CECO2+28CECH4+265CEN2O
(3)
式中,GWP为全球增温潜势(kg/hm2). IPCC指出最近100年的时间尺度内,同质量的CH4和N2O的增温效应分别是CO2的28倍和265倍(IPCC, 2013).
1.3.2 数据分析 本研究数据使用IBM SPSS Statistics 23进行单因素方差分析ANOVA,采用林肯检验比较不同淹水时期[25]、不同植被类型下温室气体排放通量的差异,采用SigmaPlot 12.5软件完成绘图.
2 结果分析
2.1 不同植被类型湿地沉积物中主要环境因子
从表1可以看出,0~10 cm和10~20 cm的土壤含水率排序为:苔草>芦苇>杨树林,苔草湿地含水率显著高于芦苇和杨树林湿地(P<0.05);20~30 cm的土壤含水率无显著差异(P>0.05). 无论哪个土层,杨树林湿地的土壤含水率均为最低. 0~10 cm土层中,杨树林湿地土壤中全氮含量显著高于芦苇和苔草湿地(P<0.05),20~30 cm不同植被类型土层之间全氮含量差异不显著(P>0.05). 0~10 cm的土层中,杨树林的铵态氮和有机碳含量均显著高于芦苇和苔草湿地(P<0.05),在其他土层中差异不明显. 除此之外,各个土层下不同植被类型硝态氮含量差异不明显(P>0.05).
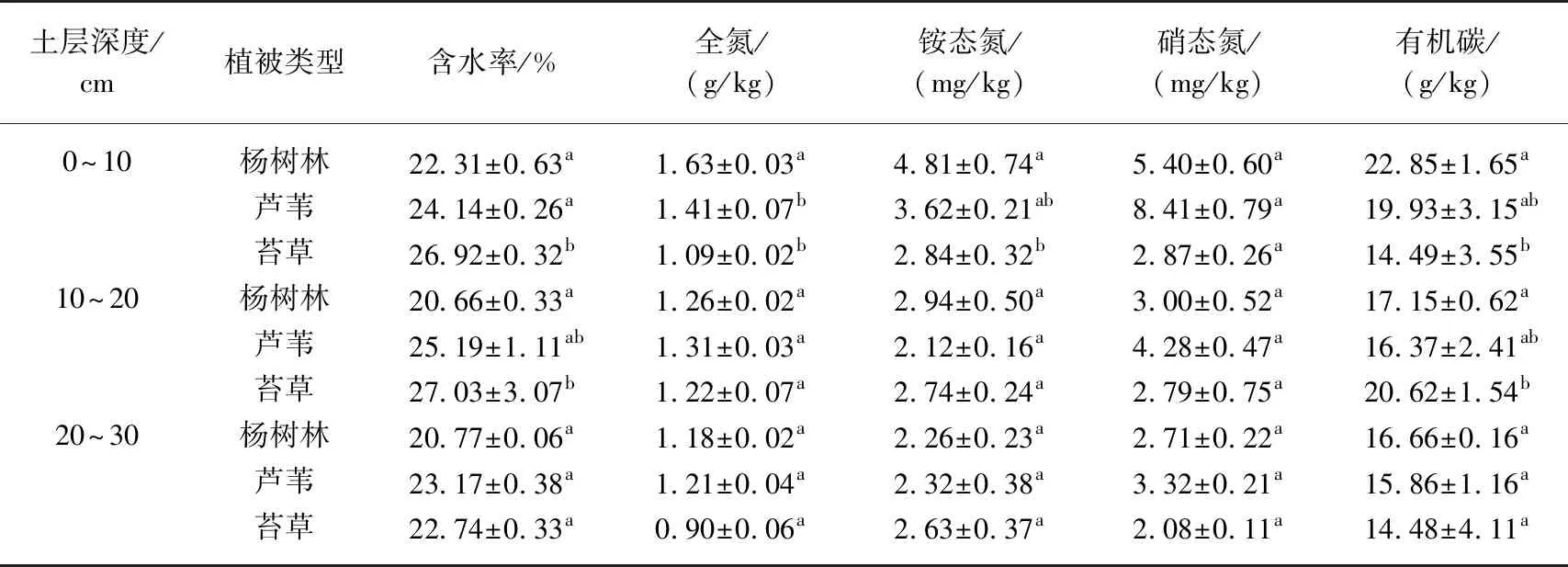
表1 不同植被类型湿地土壤理化性质*
*数据为平均值和标准误. 分别对3组数据(0~10、10~20、20~30)各项指标进行了多重比较,标记相同字母表示该组数据无显著性差异,标注不同字母则表示有显著性差异(P<0.05).
2.2 温室气体排放通量年内变化趋势
各个植被类型3种温室气体(CO2、CH4和N2O)排放通量在淹水条件发生改变后,表现出基本一致的变化趋势. 对于CO2而言,在7月淹水后其在3种植被-土壤类型下的排放通量均明显降低(图1a),在淹水期间,8、9月,芦苇植被-土壤体系的CO2排放通量有所上升,苔草和杨树林植被-土壤体系的CO2排放通量波动不大;对于CH4而言,在7月淹水后其在芦苇和杨树林植被-土壤体系的排放通量均明显增加(图1b),在9月达到最大值,芦苇湿地为144.59 mg/(m2·h),杨树林湿地为18.55 mg/(m2·h),而淹水后苔草湿地甲烷的排放通量有所波动,但变化趋势并不明显. 对于N2O而言,淹水后,在3种植被-土壤类型下的排放通量稍有降低,但变化趋势并不明显(图1c). 在排水后,3种温室气体(CO2、CH4和N2O)发生较大的波动. CO2和N2O的排放通量整体上呈现升高趋势,个别监测日其排放通量有所降低;芦苇湿地CH4的排放通量明显呈降低趋势,而苔草和杨树林湿地CH4的排放通量波动较大.
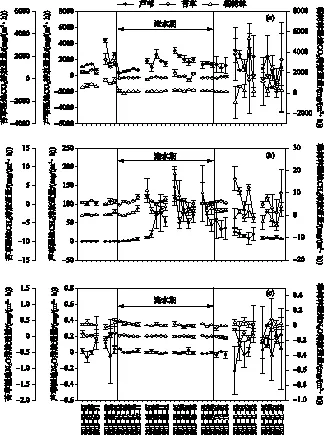
图1 不同植被覆盖下主要温室气体排放通量的季节变化Fig.1 The seasonal dynamics of main greenhouse gas fluxes under different vegetation covers
2.3 季节性淹水对不同类型植被-土壤体系温室气体排放通量的影响
将整个实验进程分为3个阶段,即淹水前(2018年5月6日-7月1日)、淹水期(2018年7月6日-9月30日)和排水后(2018年10月1日-2019年1月9日). 对不同植被类型在每个时期3种主要温室气体(CO2、CH4和N2O)排放通量的平均值进行分析,结果如图2所示. 水文条件及植被类型对CO2和CH4的排放通量均有影响,但对N2O的影响不显著(P>0.05).
对于CO2而言(图 2a),在淹水前和淹水期内,芦苇湿地的CO2排放通量均明显高于苔草和杨树林湿地(P<0.05),而苔草和杨树林湿地的CO2排放通量无显著差异(P>0.05);在排水后,芦苇湿地和杨树林湿地的CO2排放通量显著高于苔草湿地(P<0.05). 此外,淹水过程对CO2排放通量影响显著,相比于淹水前,淹水期内芦苇、苔草、杨树林3种植被类型的CO2排放通量均显著降低(P<0.05);而退水过程对CO2排放通量影响并不显著,在排水后,芦苇湿地CO2排放通量稍有降低,而苔草和杨树林湿地的CO2排放通量有所增大.
对于CH4而言(图 2b),在淹水前,各个植被类型湿地的CH4排放通量均处于较低的水平,并无显著差异(P>0.05);淹水期和排水后,芦苇湿地CH4的排放量均显著高于苔草和杨树林湿地的CH4排放量(P<0.05). 淹水过程对各植被类型CH4排放通量均有影响,淹水后CH4排放通量升高,排水后CH4排放通量降低;但淹水退水对芦苇湿地CH4的排放通量影响显著(P<0.05),对苔草和杨树林湿地CH4的排放通量影响并不显著(P>0.05).
对于N2O而言(图 2c),其排放通量极小,最大排放通量出现在排水后的芦苇湿地,为0.06 mg/(m2·h),苔草湿地对于N2O表现为弱吸收,芦苇和杨树林湿地对N2O表现为弱排放. 但各个水文时期和植被类型下,N2O的排放通量无显著差异(P>0.05).
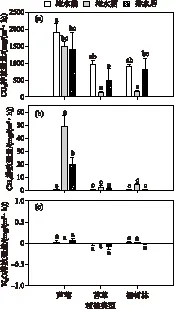
图2 不同植被类型淹水和非淹水阶段温室气体平均排放通量变化(图中的数据为平均值和标准误,方柱标注不同字母则表示有显著性差异(P<0.05))Fig.2 The effect of flooding and non-flooding on main greenhouse gas fluxes under different vegetation covers
2.4 综合增温趋势
不同植被类型湿地的3种温室气体对GWP的贡献表现出一致性(图3),贡献程度为:CO2>CH4> N2O. 整个观测期内,芦苇、苔草、杨树林3种植被类型湿地中,GWP分别为16191.3、3405.6和1883.1 kg/hm2,CO2对GWP的贡献率分别为:66.99%、101.16%和83.29%, CH4对GWP的贡献率分别为: 32.74%、4.32%和17.08%,N2O对GWP的贡献率分别为:0.27%、-5.48%和-0.37%. 由此可以看出,不同植被类型湿地主要通过增加CO2的排放通量提高了其GWP. 对于苔草和杨树林湿地而言,通过对N2O的吸收一定程度上削弱了苔草和杨树林湿地的GWP,但由于N2O的排放通量极少,影响并不明显. 通过单因素方差分析ANOVA发现,芦苇湿地对GWP的贡献显著大于苔草和杨树林湿地(P<0.05).
3 讨论
3.1 水文过程及植被对温室气体的影响
本研究发现水文过程和植被类型都会对3种主要温室气体(CO2、CH4和N2O)排放通量产生影响. 各植被类型湿地在淹水后CO2排放通量减少,且芦苇湿地CO2排放通量显著高于苔草和杨树林湿地,这与盛宣才等的研究结果基本一致[26]. 由于淹水后,沉积物氧化还原环境发生了改变[27],氧浓度降低,造成了CO2排放通量降低. 以往研究表明: 不同类型潮滩土壤呼吸之间的差异不大,植物地上部分的呼吸是引起生态系统 CO2排放通量差异的主要原因[28-29]. 本研究采用的是静态-暗箱法,测得的CO2排放通量主要包括土壤呼吸和植物暗呼吸两部分. 由于芦苇地上生物量明显高于苔草和杨树林,其通过暗呼吸所产生的CO2高于其余两种植被,从而导致芦苇湿地CO2排放通量显著高于苔草和杨树林湿地. 而在淹水后芦苇湿地CO2排放通量降低幅度小于苔草和杨树林湿地,其原因可能是芦苇具有发达的通气组织,可以向根系输送一定量的氧气以改变区域的缺氧环境[30],并且根部产生的CO2可以通过通气组织上传到空气中. 排水后,芦苇湿地CO2排放通量比淹水期小,这可能是由于排水后期到了秋冬季,温度降低、芦苇开始枯萎死亡,许鑫王豪等对苏北潮滩的研究也表明CO2的最低排放通量出现在冬季[29]. 而苔草和杨树林湿地在排水后温室气体排放通量升高,则可能是由于氧化还原环境的瞬间改变提供了足够的氧气,导致CO2的排放量升高.
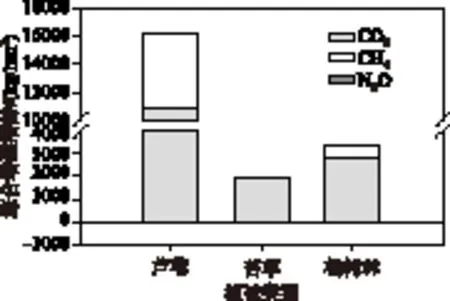
图3 监测期间不同植被类型综合增温潜势Fig.3 Global warming potential of different vegetation types during the monitoring period
淹水期间,各植被类型的CH4排放通量均有所增加,该结果与前人的研究结果具有一致性[31-32]. 可能是由于淹水造成了严格的厌氧环境,厌氧呼吸成为了绝大多数湿地碳降解的主要途径,最终产物一般为CH4[33]. 此外,很多研究表明植被种类对于CH4排放通量有着重要的影响[5, 34]. 本研究结果显示在淹水后芦苇湿地CH4排放通量增加显著(P<0.05),苔草和杨树林湿地CH4排放通量也有所增加,但不显著. 该结果与他人的研究结果有所差异,Ding等[35]和Maucieri等[36]的研究均表明莎草科植物对甲烷的传输能力大于禾本科. 虽然在本研究中苔草和杨树林湿地中生长的植被主要为莎草科植物苔草和藨草,芦苇湿地主要生长的植被为芦苇,但芦苇湿地甲烷排放通量在淹水后显著增高的可能原因如下:在淹水0.4 m后苔草和杨树林湿地植被几乎被淹没到水面以下,甚至一段时间后根部发生枯萎腐烂,降低了其运输温室气体的能力,相对比而言,芦苇作为挺水植物,大部分茎叶仍处于水面之上,发达的通气组织为运输CH4提供了通道. 排水后,各个类型湿地的CH4排放通量均有所降低. 这是由于在排水后,环境由无氧状态变为有氧状态,氧化反应增强,因此CH4排放通量降低.
在整个监测期内,不同植被类型在不同水文条件下N2O的排放通量无显著差异(P>0.05). 多项研究表明,N2O的产生与土壤性质、含水率、电导率等环境因素及微生物种群等生物因子均存在密切关系[37-38],但N2O 的产生机制很复杂,目前仍未得出统一结论. 但可以明确的是N2O主要来源于微生物参与下的硝化反硝化反应,土壤含水量低和土壤长期持续淹水都不利于硝化和反硝化细菌的生长[33];也有研究表明植被类型会对N2O的排放通量产生一定的影响[39-40].
3.2 全球增温潜势
本研究发现,不同植被类型湿地对于GWP的贡献不同,由大到小排序为:芦苇>杨树林>苔草. 许鑫王豪也曾通过研究指出,植被对温室气体通量的排放起着至关重要的作用,植被存在与否以及植被类型差异均会影响温室气体排放,进而导致GWP存在一定的差异[29]. Kandel等在监测了干旱的河流再被重新淹水恢复后的温室气体排放情况后发现,在重新被淹水恢复的湿地中CO2及N2O的排放通量较低,但CH4排放通量持续处于较高水平[41]. 西洞庭湖湿地自2013年开展了“退林还湿”工程,清除了保护区核心区内的杨树林,开展湿地恢复,其主要目的是恢复候鸟栖息地及恢复湿地生态功能. 湿地作为一个重要的碳库,合理的恢复及管理有望恢复天然湿地的碳汇功能,因此在恢复过程中温室气体排放量的变化应该得到重视. 而本研究结果,在西洞庭湖目前的恢复阶段,其碳汇功能可能尚未实现,但我们不难发现芦苇湿地的GWP明显高于其余两种植被类型,因此我们建议,在可能的情况下,不再人为地种植芦苇,尽可能地将杨树林恢复为苔草湿地,更有利于减少温室效应.
猜你喜欢
小学生学习指导(低年级)(2021年10期)2021-11-01 08:22:58
鸭绿江(2020年15期)2020-11-12 02:07:28
金桥(2020年8期)2020-05-22 06:22:54
前卫文学(2017年5期)2017-11-09 17:27:39
环境科技(2016年5期)2016-11-10 02:42:04
音乐天地(音乐创作版)(2016年4期)2016-07-28 09:39:37
现代农村科技(2016年20期)2016-03-28 23:34:54
林业与生态(2016年3期)2016-02-27 14:24:22
江西理工大学学报(2015年3期)2015-12-22 05:26:17
现代农业科技(2014年7期)2014-07-07 10:23:54
