
海岛棉GbGPAT2基因的克隆与表达分析
2020-05-05 03:14甄军波刘琳琳杜海英张建宏迟吉娜
西南农业学报 2020年3期
甄军波,刘琳琳,杜海英,刘 迪,蔡 肖,张建宏,迟吉娜
(河北省农林科学院棉花研究所/农业部黄淮海半干旱区棉花生物学与遗传育种重点实验室,河北 石家庄 050051)
【研究意义】棉花是世界上重要的经济作物,棉籽油也是大豆、油菜、向日葵和花生之外重要的油料来源之一[1],利用棉籽油开发生物柴油,既能提高棉籽的综合利用率,又能实现能源的可再生,降低对化石能源的依赖,从而保护生态环境。甘油-3-磷酸酰基转移酶(GPAT)是植物三酰甘油类合成过程中的限速酶,相比于其他作物,棉花GPAT基因的研究相对较少,发掘棉花GPAT基因,对于深入解析棉籽油合成机理、提供棉籽含油量具有十分重要的意义[2-5]。【前人研究进展】甘油-3-磷酸酰基转移酶(GPAT)是植物生物膜中磷脂酰甘油(PG)生物合成过程中的第一个酰基酯化酶,是Kennedy途径的第一步,被认为是TAG生物合成的“阀门”,其对底物的选择性对决定PG分子中的不饱和程度起着关键性作用,与种子油份关系密切[6]。张楠[7]和朱双[8]等分别从小桐子中克隆了GPAT基因,分别命名为JcGPAT和JcGPAT2。拟南芥中鉴定了10个GPAT基因,包含AtGPAT1-9和AtATS1。AtATS1定位于质体,AtGPAT1-3定位于线粒体膜,AtGPAT4-9则定位于内质网膜[2]。AtGPAT1在拟南芥花粉管发育中发挥重要作用[9]。Men等[10]研究表明,OsGPAT3与水稻的花粉管发育和育性有关,并推测GPAT3基因在单子叶和双子叶植物中发挥的作用不尽相同。Sui等[11]将碱蓬的SsGPAT基因转化拟南芥,提高了转基因拟南芥的耐盐性,通过增加拟南芥不饱和脂肪酸含量,从而降低了高盐胁迫对于光合系统的影响。看见,GPAT家族基因在植物生长发育中发挥着重要的作用,有必要进一步深入研究。【本研究切入点】本研究从海岛棉中同源克隆获得一个新的GPAT基因,命名为GbGPAT2(登陆号为:KC292646),对该基因进行了生物信息学分析,通过实时定量PCR检测了该基因在幼胚发育不同时期的表达特异性,并在烟草中对其亚细胞定位进行了验证。【拟解决的关键问题】为进一步研究GbGPAT2在棉花中的功能提供理论基础。
1 材料与方法
1.1 试验材料
1.1.1 植物材料 用于目的基因克隆的海资8号(海岛棉)为河北省农林科学院棉花研究所分子设计育种研究室提供,种植于河北省农林科学院棉花研究所试验地。种子置于黑暗培养箱中,经12 ℃处理8 h,恢复25 ℃正常条件培养,取正在萌发的种子,立即放于液氮中,备用;分别取开花后16、25、30、35 d 的胚,液氮速冻,用于检测GbGPAT2的表达量。
1.1.2 质粒、菌种与试剂 克隆载体pT-Easy Vector、Premix ExTaq、DNA标准分子量DL2000、琼脂糖凝胶回收试剂盒、质粒小量提取试剂盒、反转录试剂盒均购自TransGen公司;植物总RNA提取试剂盒购自北京奥莱博生物有限公司;其他常规试剂均为国产分析纯。
1.2 总mRNA 提取和反转录
采用北京奥莱博生物有限公司Plant RNA Kit 提取棉花种子和未成熟胚的总RNA,具体方法参照试剂盒说明书。采用TransGen反转录试剂盒,进行反转录。
1.3 GbGPAT2基因的克隆
以蓖麻ACB30456.1氨基酸序列提交NCBI,获得棉花EST,通过DNAMAN 软件拼接序列,获得电子克隆片段。利用Primer5设计引物扩增基因全长,引物序列为:GPAT-S: ATGAACAGTAGTGAAGGGAAGTTGAAATCA,GPAT-A: TCATTTTTCTTCCAGTTCCAGTCCCCG,引物由Invitrogen公司合成。以cDNA为模板进行PCR,PCR 程序为94 ℃预变性4 min, 94 ℃ 1 min, 55 ℃ 1 min, 72 ℃ 1 min 30 s, 共30 个循环,72 ℃ 10 min,4 ℃ 保温。将PCR产物在含Goldview的1.0 %琼脂糖凝胶上进行电泳分离,电泳缓冲液为1×TAE,电泳30 min后,置于Biored凝胶成像系统观察照相。用DNA分子量Marker DL2000判断扩增产物的分子量大小。以琼脂糖凝胶分离纯化试剂盒回收PCR扩增产物,克隆到pT-EASY载体上,然后转化感受态大肠杆菌TransT3。经菌落培养和蓝白斑筛选随机挑取8个克隆进行PCR 阳性验证,将获得的3个阳性单克隆送Invitrogen公司测序。单克隆序列通过NCBI比对,确定克隆序列为目的基因,提交NCBI的ORF finder预测其开放阅读框,并经BLASTX同源比对验证。
1.4 序列的生物信息学分析
用NCBI (http://ncbi.nlm.ncih.gov/)中的BLAST、ORF及ExPASY (http://expasy.org/tools/)中的ComputepI/Mw tool 等软件进行生物信息学分析;通过NCBI CDD数据库(Conserved Domain Databases), 进行保守结构域的分析;采用ProtParam (http://expasy.org/tools /protparam.html) 进行氨基酸基本理化性质分析;利用Predictprotein (http://www.predictprotein. org/)和SignalP (http://www.cbs.dtu.dk/services/SignalP/)分别预测跨膜区、功能位点和信号肽;蛋白质的疏水性由ProtScale tool (http://expasy.org/tools/pscale/Hphob.Woods.html)预测。以GbGPAT2作为查询母序列,在NCBI的GenBank数据库中搜索与该基因同源的其他物种的氨基酸序列,通过ClustalX软件进行氨基酸序列比对,采用MEGA4软件构建系统进化树。
1.5 qRT-PCR检测胚发育过程中的表达特异性分析
选取基因特异性较高的序列,设计qRT-PCR引物,qG1:GTGTTTTAAGTCGAATCCCCCG;qG2:CCCTCTCCATCTTTTTCCGCAA。选取开花后16、25、30、35 d的胚,提取RNA,反转录获得cDNA,在德国耶拿荧光定量PCR仪中检测该基因的表达模式,以组成型表达的泛素基因Ubiquitin7作为内参,PCR程序为: 95 ℃ 2 min; 95 ℃ 15 s, 60 ℃ 15 s, 72 ℃ 30 s, 40个循环,实验重复3次。

表1 GbGPAT2氨基酸组成分析Table 1 Composition of the amino acids in GbGPAT2
1.6 GbGPAT2基因的亚细胞定位
设计引物GbGPAT2-F和GbGPAT2-R,将GhbHLH开放阅读框连接至亚细胞定位载体pCAMBIA1302,挑取单克隆测序验证后转化农杆菌GV3101,空载体pCAMBIA1302作为对照,农杆菌菌液注射烟草叶肉细胞,黑暗过夜培养后,在激光共聚焦显微镜下观察亚细胞定位结果。
2 结果与分析
2.1 棉花GbGPAT2 全长序列克隆
根据特异引物在cDNA中的扩增,获得约1300 bp左右片段(图1),经克隆后测序,发现GbGPAT2基因序列全长为1294 bp,包含 1 个1113 bp 的开放阅读框,编码370 个氨基酸的多肽,预测分子量为42.4 kD,等电点为8.72,5′端非翻译区长40 bp,3′端非翻译区长141 bp。氨基酸组成成分中,极性氨基酸占24.05 %,疏水性氨基酸占37.57 %(表1),在GenBank中登录号为KC292646。
2.2 GbGPAT2的功能结构域分析
用NCBI的Conserved Domains程序预测蛋白的保守区,发现本研究克隆的基因编码的蛋白是溶血磷脂酰基转移酶 (LPLAT-LPCAT1-like) 超家族成员(图2),该基因编码的蛋白具有一个典型的溶血磷脂酰基转移酶相似家族保守区(LPLAT-LPCAT1-like)和酰基转移酶超家族保守结构域(NAT-SF superfamily)。

M: D2000; 1: GbGPAT2图1 海岛棉GbGPAT2同源基因的PCR扩增Fig.1 PCR amplification of GbGPAT2 homology in Gossypium barbadense
2.3 GbGPAT2与其他植物GPAT氨基酸序列多重比对
用BLASTX同源分析显示,海岛棉GbGPAT2氨基酸序列与其他植物GPAT具有较高的同源性(图3)。GbGPAT2与小桐子、拟南芥GPAT氨基酸序列同源性在87 %~92 %,而与已公开的陆地棉GhGPAT氨基酸序列同源性只有8 %。GbGPAT2编码的氨基酸有4个序列保守区域,其中motif-1的组氨酸(H)和天冬氨酸(D)残基、motif-3的甘氨酸(G)残基、motif-4的脯氨酸(P)残基都高度保守,被认为是酰基转移酶家族中重要的催化位点[12]。motif-2的苯丙氨酸(F)和精氨酸(R)以及motif-3中的丝氨酸(S),在甘油-3-磷酸酰基转移酶的底物结合中起重要作用[13]。
2.4 GbGPAT2编码蛋白的跨膜区分析
通过在线的亲/疏水性分析(http://cn.expasy.org/cgi-bin/protscale.pl),发现GbGPAT2的N端肽链存在一定范围的疏水区(图5,0以上的区段为疏水区),而其他部分则存在较大范围的亲水区,有较强的亲水性(图4A)。进而通过在线的TMHMM(http://www.cbs.dtu.dk/services/TMHMM/)对编码蛋白跨膜区进行了预测,发现GbGPAT2具有2个显著的跨膜区,分别在第92~144氨基酸处和第127~144氨基酸处(图4B)。
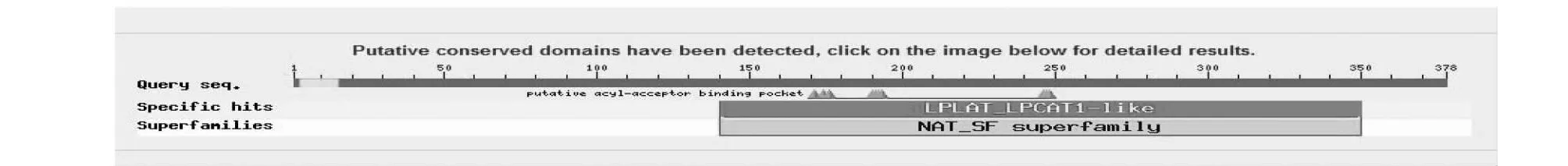
图2 GbGPAT2 氨基酸序列保守结构域Fig.2 Conserved domain of GbGPAT2

黑线标示区域依次为GPAT保守域1-4 图3 不同植物GPAT蛋白保守区的多序列分析Fig.3 Alignment of deduced amino acid of GPATs from different plants
2.5 GbGPAT2二级结构预测
利用DNAStar软件分析发现GbGPAT2编码的肽链含36.4 %的α-螺旋,38.3 %的β-折叠,15.1 %的转角构象,10.5 %的无规则卷曲(图5)。

图4 GbGPAT2疏水性和跨膜区预测Fig.4 Prediction of hydrophobicity and trans-membrane region of the GbGPAT2 amino acid sequence

图5 GbGPAT2氨基酸序列DNASTAR-Protein分析结果Fig.5 Result of GbGPAT2 amino acid sequence analyzed by DNAStar-Protein programe

图6 GbGPAT2与其他物种GPATs氨基酸系统进化树Fig.6 Phylogenetic tree analysis of GPATs amino acids
2.6 GbGPAT2同其他物种的进化关系分析
构建陆地棉、拟南芥和小桐子等植物的GPAT氨基酸系统进化树,发现GbGPAT2在分化时间上较晚,能与小桐子的JcGPAT和拟南芥的AtGPAT9亲缘关系较近,而与陆地棉已有的GPAT进化关系较远(图6)。
2.7 GbGPAT2在幼胚中的表达模式
在种子形成的胚发育的不同阶段,选取16 DPA、25 DPA、30 DPA、35 DPA胚,提取RNA,利用GbGPAT23′-UTR区特异引物,以组成型表达的泛素基因Ubiquitin7作为内参基因,通过qRT-PCR检测了其表达特异性(图7)。结果表明,GbGPAT2在胚发育的不同时期表达丰度不同,在开花后25 d 的胚中表达量最大,随着胚发育的逐渐成熟,GbGPAT2表达量逐渐下降。
2.8 GbGPAT2亚细胞定位
为了验证GbGPAT2的亚细胞定位信息,本研究构建了pCAMBIA1302-GbGPAT2-GFP融合表达载体,利用农杆菌将空载体和融合载体转化烟草叶肉细胞,暗培养后,利用激光共聚焦显微镜观察绿色荧光表达情况,结果表明,GbGPAT2定位于细胞质膜中(图8),与预测结果一致。
3 讨 论
研究表明,在植物细胞中GPAT主要存在于质体、内质网和线粒体等部位,其中定位于质体中的GPAT是可溶性蛋白,以acyl-ACP为底物进行催化反应,而定位于线粒体和内质网膜上的则是结合蛋白,以acyl-CoA和acyl-ACP为底物[14]。本研究中从海岛棉中克隆了GbGPAT2基因,该基因开放阅读框全长1113 bp,在棉花胚发育25 d表达量最高,在烟草中的亚细胞定位表明,该基因定位于细胞质膜之中。
由于GPAT在sn-1和sn-2进行催化反应的偏好性不同,即使属于同一亚组,GPAT也可能发挥不完全相同的作用[10,15]。在拟南芥中已经发现了10个AtGPAT基因(AtGPAT1-9和ATS),其中AtGPAT8和AtGPAT9同源,定位于内质网膜上,有着相似的功能,在Kennedy途径中控制TAG的生物合成[2]。
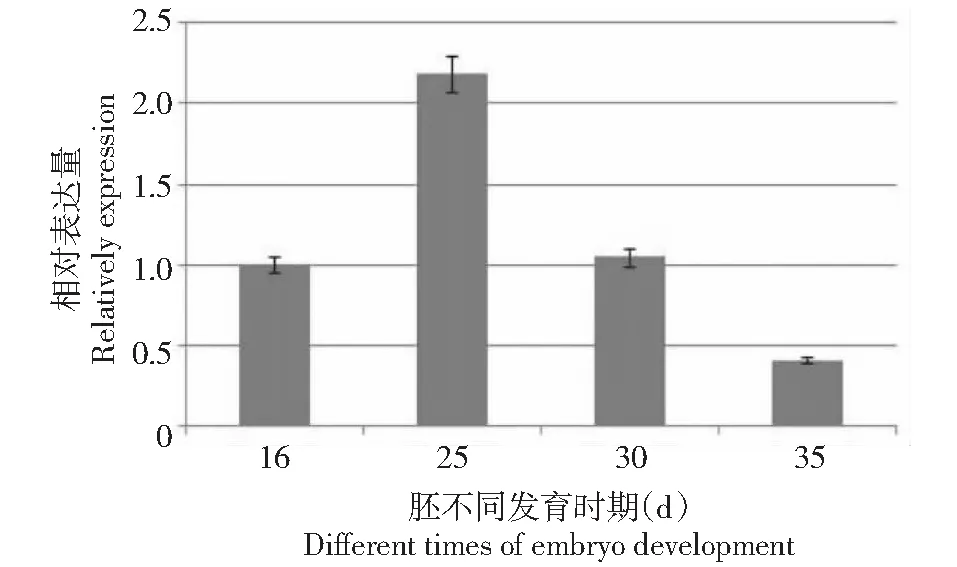
横坐标为胚不同的发育时期(16DPA、25DPA、30DPA、35DPA),纵坐标为相对表达量图7 GbGPAT2在胚发育不同时期的表达特异性分析Fig.7 Real-time PCR of GbGPAT2 in different times of embryo development
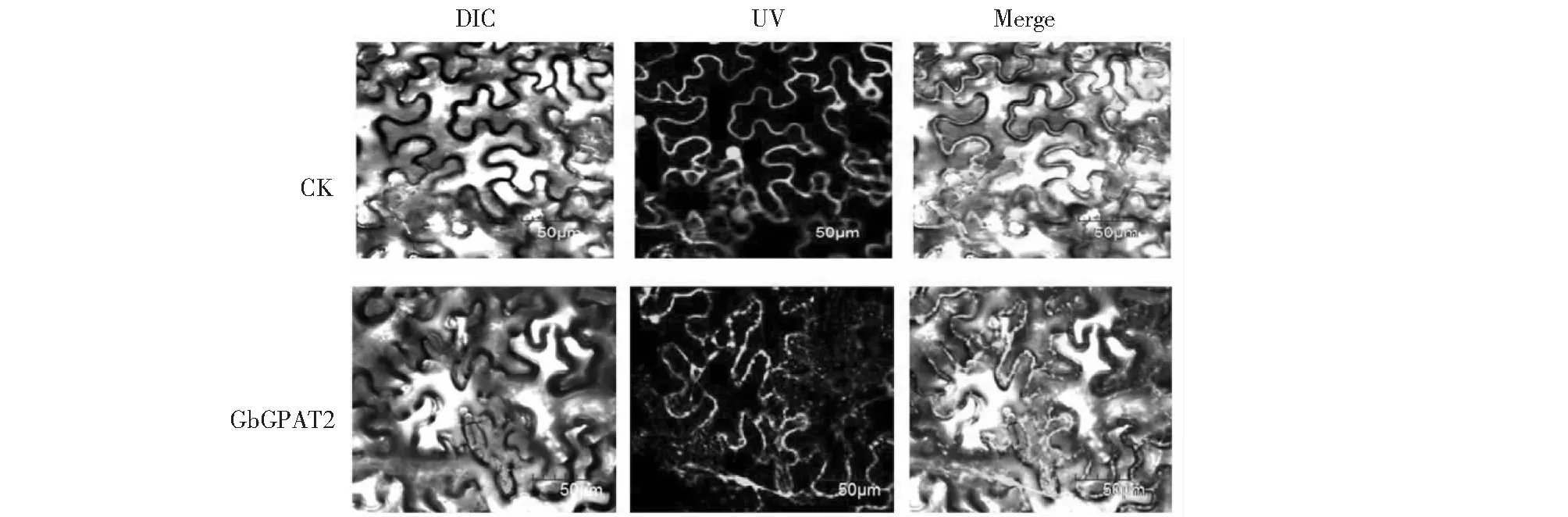
图8 GbGPAT2亚细胞定位Fig.8 Subcellular localization of GbGPAT2
GPAT在植物脂类代谢中功能多样,不但参与TAG的合成,而且涉及膜脂的生物合成和植物的抗性[12-14]。国内外研究者已经从已先后从小桐子[7-8]、拟南芥[9]、番茄[13]、甜辣椒[16]、豌豆[17]等多种植物中克隆到了GPAT基因。将拟南芥和菠菜的GPAT基因分别转入水稻中,使水稻叶片中磷酸甘油的顺式不饱和脂肪酸含量得到了显著提高[18]。在拟南芥中过表达GPAT基因能够显著提高种子含油量[14,19],拟南芥AtGPAT5在种皮和根中优势表达,对于拟南芥软木脂等聚酯类物质的合成以及脂类聚合物的结构都具有重要作用[20]。构建的氨基酸序列系统进化树表明,GbGPAT2与拟南芥AtGPAT9和小桐子JcGPAT(HQ395225)基因的亲缘关系较近,同样具有高度保守的motif 1-4,以及对应的氨基酸残基,推测GbGPAT2基因可能与拟南芥AtGPAT8、AtGPAT9和小桐子的JcGPAT具有类似的功能。本研究克隆的GbGPAT2可能参与脂类代谢,在提高种子油含量方面发挥着重要的作用,具体功能如何有待于进一步的研究。
猜你喜欢
热带作物学报(2022年7期)2022-08-06
中国农业科学(2022年14期)2022-07-26
保健与生活(2022年11期)2022-06-09
保健与生活(2021年5期)2021-04-12
园艺与种苗(2020年12期)2021-01-08
含笑花(2019年3期)2019-06-12
文苑(2018年20期)2018-11-09
新农村(2018年13期)2018-08-20
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
