
星鸦(Nucifraga caryocatactes)对集中放置红松球果的分散贮藏与临时贮点管理
2020-04-30 04:32:00郑骄阳吕鸿达
野生动物学报 2020年2期
郑骄阳 吕鸿达 程 鲲 宗 诚
(东北林业大学野生动物与自然保护地学院,哈尔滨,150040)
食物贮藏是动物重要的适应性行为,也是许多植物尤其是大种子植物传播种子的主要机制[1]。分散贮藏是贮食动物将贮藏物存放在离散且间隔较远的贮点,将潜在的食物损失率降到最低[2-5],分散贮食研究主要包括行为特点[6]、形成原因[7]、影响因素[8]以及对植物天然更新的影响等[9-11]。关于其行为动机,学者们提出了快速隔离、缺乏贮藏空间和避免盗窃等假说[12]。其中快速隔离假说认为,分散贮藏行为是对丰富却短暂的食物资源的一种竞争性策略,当食物资源丰富时,动物的贮食活动加剧,在食源附近迅速地分散贮藏食物,其搬运距离小,有效地缩短了动物单次贮食时间,以最短的时间获取更多的食物,从而提高贮藏效率[13]。
分散贮藏动物会通过移动、取食、重取、二次贮藏以及盗取等行为持续对种子进行存储管理[14-15]。二次贮藏是指贮食动物将贮点内种子取出、搬运至新贮点,这种行为在啮齿动物[14-17]和鸟类[18-21]中均有发现。大量研究表明,当种子被二次贮藏时,与食物源的距离会增加[21-23],二次贮藏可能通过改变种子与母树的距离来影响幼苗建立的概率[24-26]。在动物贮藏管理的过程中,贮点中的种子都有可能会由于偷盗、种子易腐性、病虫感染或被贮食者遗忘而丢失,而未被取食的完好种子则对植物更新起到很大作用[27]。
在黑龙江凉水自然保护区,红松(Pinuskoraiensis)与松鼠(Sciurusvulgaris)、星鸦(Nucifragacaryocatactes)等贮食动物之间通过协同进化形成了互惠共生网络[28-31]。其中星鸦作为红松种子的主要传播者,其分散贮藏行为对原始红松林生态系统的维持有着重要作用。每年8月末凉水保护区人工采摘的红松球果会短期集中堆放,直到10月进行去壳处理,这为贮食动物提供了集中的食物源。2017年秋季,我们观察到大群星鸦啄取松塔堆的松子并在附近区域分散贮藏,而且对贮点不断更新,重取后又埋藏,具有2次贮藏和临时贮点管理行为。本研究通过探究星鸦对集中红松种子源的分散贮藏和重取行为,分析星鸦贮点特征、贮点生境选择及食物密集贮藏期的临时贮点管理过程,旨在进一步丰富动物的分散贮藏理论,探讨星鸦对红松种子传播和天然林更新的作用。
1 研究地区与研究方法
1.1 研究地概况
黑龙江凉水国家级自然保护区位于中国小兴安岭东南部,地理坐标为128°48′08″—128°55′46″E,47°07′15″—47°14′38″N。保护区总面积为6 394 hm2,南北长11 km,东西宽6.25 km。保护区核心区3 740 hm2,其中原始红松林面积2 375 hm2。本研究地在凉水自然保护区管理局附近的第19林班。
保护区全境属于低山地貌,地形北高南低,相对高差一般为100—200 m,平均坡度为10—15°,最大坡度为40°。属温带大陆性季风气候,冬长夏短。保护区地带性植被是以红松占优势的针阔混交林,分为山地和谷地植被两大类。上世纪前半叶对红松的拔大毛式采伐使原始阔叶红松林被分割成大小不一的斑块,夹杂不同演替阶段的次生林、人工林和林间空地[32]。
保护区主要乔木树种有红松、红皮云杉(Piceakoraiensis)、鱼鳞云杉(Piceajezoensis)、臭冷杉(Abiesnephrolepis)、兴安落叶松(Larixgemliniivar.gmelinii)、胡桃楸(Juglansmandshurica)、黄檗(Phellodendronamurense)、水曲柳(Fraxinusmandshurica)、春榆(Ulmusdavidianavar.japonica)、硕桦(Betulacostata)、白桦(Betulaplatyphylla)等。林下灌木主要有金花忍冬(Lonicerachrysantha)、毛榛(Corylusmandshurica)、珍珠梅(Sorbariasorbifolia)、绣线菊(Spiraeasalicifolia)、刺五加(Eleutherococcussenticosus)等。林下草本种类繁多,主要有羊须草(Carexcallitrichos)、毛缘薹草(Carexpilosa)、问荆(Equisetumarvense)及各种蕨类植物等。
保护区内动物区系组成丰富,共有鸟类17目47科254种,兽类6目16科51种,其中啮齿类17种。其中松鼠、花鼠(Tamiassibiricus)、星鸦和普通(Sittaeuropaea)与红松种子之间形成了稳定的贮食传播网络[28-31]。
1.2 研究方法
1.2.1 星鸦贮食行为监测
2017年10—11月,对星鸦贮食行为进行红外相机监测。根据红松球果堆放时间,将监测分为2个时间段,10月1—15日为球果堆放期,此阶段松塔被集中堆成大堆,星鸦啄食松子,在附近埋藏;10月16日之后为贮点重取期,这期间松塔堆已经移走,星鸦陆续将埋藏松子重取。将相机捆绑于距地面10—20 cm的树干,设置为拍照2张后,录制15 s视频。记录每张照片的星鸦数量、贮食行为持续时间、温度(℃)以及干扰因素。第一阶段设置11台相机(型号:H6W),第二阶段设置6台相机。
1.2.2 贮食行为谱
根据对星鸦贮食行为的观察,将红外相机照片和视频中的行为类型进行分类定义,确定贮食行为谱,包括贮藏、重取、移动、争斗、警戒等行为(表1)。
1.2.3 贮点调查
贮点调查时间为10月1—7日。在距松塔堆1 km内选取5块样地,其生境类型分别为人工针叶林、林间空地、次生针阔混交林、云冷杉林、原始红松林,记录样地距食物源距离、距道路距离(表2)。每块样地选取10个样方(由于人工针叶林样地贮点数量多且集中,选取了22个样方),样方大小为2 m×2 m。
首次贮点调查时,拨开样方中的枯枝落叶层,在找到的贮点旁插上标记,记录贮点特征:贮点大小、贮点深度、基质类型、基质硬度。其中基质类型分为草地、苔藓、枯草地以及裸地。基质硬度分为软、较软、较硬、硬。贮点重取调查时,贮点为空时记为重取贮点,贮点数量保持原样则记为留存贮点,如发现新的贮点记为新建贮点。重取率指重取贮点占所有贮点的比例;更新率指新增贮点占所有贮点的比例。
表1 星鸦贮食行为谱
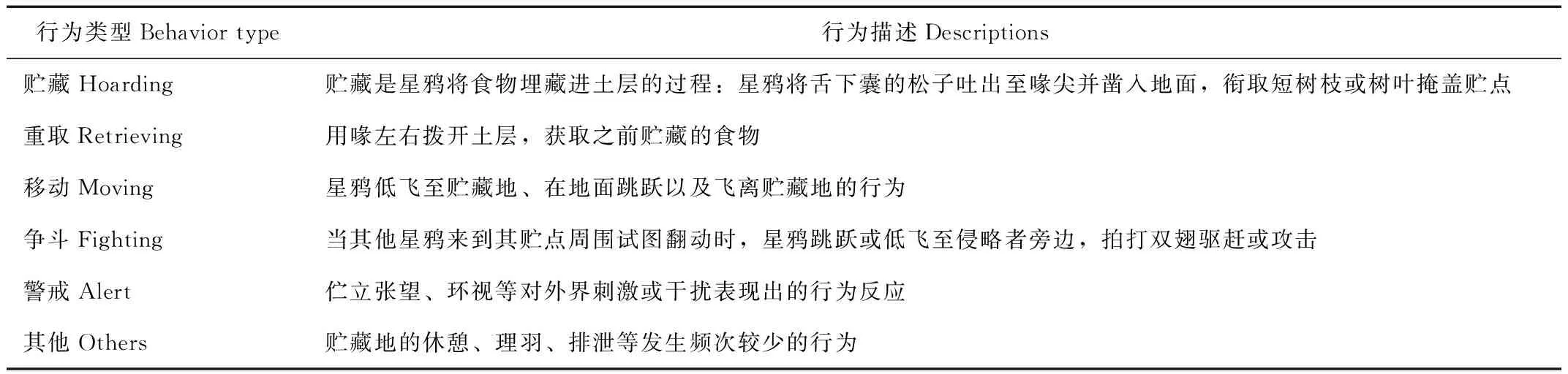
Tab.1 The hoarding behavioral spectrum of spotted nutcrackers
表2 星鸦贮点调查样地描述
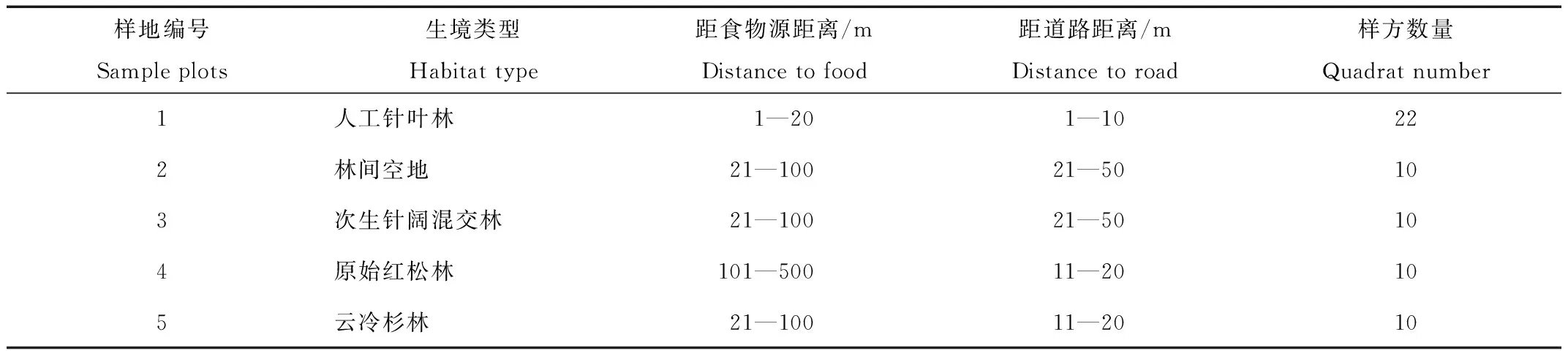
Tab.2 Description of sample plots for spotted nutcracker cache investigation
1.2.4 数据统计分析
数据描述用算术平均值±标准误表示,置信度95%。用单因素方差分析不同生境类型及基质特征中贮点特征的差异。数据处理、统计分析及作图在Excel 11.0、R 3.5.2、SPSS Statistics 22及Sigmaplot 10.0中完成。
2 结果与分析
2.1 贮食行为特征
2.1.1 贮食行为时间分配及节律
获得星鸦红外相机照片1 864张,记录到的星鸦行为频次共2 409次,主要行为类型有贮藏、重取、移动、争斗和警戒行为(图1)。其中,重取行为是最主要的行为类型,占(55.42±1.34)%;移动行为占(25.74±6.24)%;贮藏行为占(11.08±2.69)%;警戒行为和争斗行为分别占(3.28±0.79)%和(2.16±0.52)%;其他行为占(2.33±0.57)%。
星鸦重取行为的时间比例从早晨、中午到下午逐渐增加,8:00—9:00、10:00—12:00和16:00—17:00有2个活动高峰。贮藏行为与重取行为的活动高峰相邻,贮藏高峰出现在重取行为高峰之前,主要集中在早晨,5:00—6:00、7:00—8:00和9:00—10:00有3个活动高峰,中午和下午贮藏行为逐渐减少。移动行为仅在7:00—8:00出现1个峰值,其他时间段的时间比例较为平均。警戒行为5:00—6:00和15:00—17:00有2个峰值,争斗行为6:00—7:00和15:00—16:00有2个峰值,警戒行为活动高峰在争斗行为高峰之前(图2)。
2.1.2 贮藏地星鸦活动频次
球果堆放期的10月4—15日,监测样地的星鸦活动非常频繁,每台相机记录到星鸦的平均活动频次为每天(21.43±5.73)次。堆放球果被移走后,10月16日直至11月初,星鸦一直有持续的重取行为,但贮点重取期活动频次下降为(3.30±0.68)次/d/相机(图3)。
2.2 贮点特征
2.2.1 贮点大小
共调查了62个样方,记录了451个贮点、1 079粒红松种子。贮点平均大小为(2.39±0.09)粒,多为1粒或2粒松子,分别占35.48%和32.15%,最大贮点为20粒松子。原始红松林贮点的平均松子粒数相对较多,为(3.50±0.84)粒;其次为林间空地(2.65±0.34)粒、次生林(2.44±0.20)粒、人工针叶林(2.41±0.11)粒,云冷杉林贮点相对较小,为(1.87±0.19)粒。但不同生境类型的星鸦贮点大小没有显著差异(F=1.679,P>0.05)。
2.2.2 贮点深度
星鸦贮点的贮藏深度为(1.55±0.03)cm。不同生境类型的贮藏深度有显著差异(F=17.171,P<0.05)。贮点埋藏最深的生境类型为原始红松林,为(2.50±0.71)cm;其次为次生林(1.78±0.06)cm、人工针叶林(1.59±0.03)cm、云冷杉林(1.35±0.09)cm,林间空地的贮点埋藏最浅,为(0.95±0.11)cm。
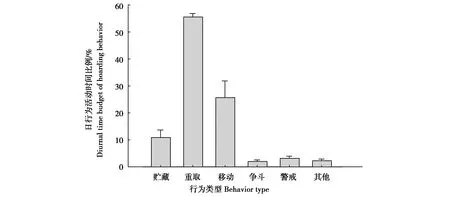
图1 星鸦贮食行为日活动时间分配Fig.1 Diurnal time budget of hoarding behavior of spotted nutcrackers
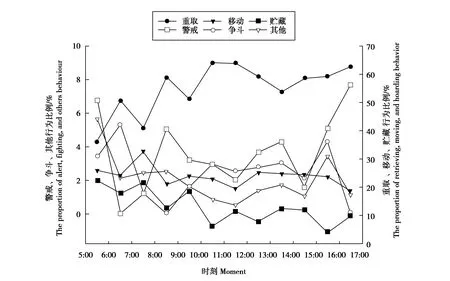
图2 星鸦贮食行为节律Fig.2 Hoarding behavior rhythm of spotted nutcrackers

图3 球果堆放期和贮点重取期星鸦的活动频次Fig.3 The activity frequency of the spotted nutcracker during cone piling period and retrieving period
2.2.3 贮点密度
各样方贮点的平均密度为(1.82±0.11)个/m2。不同生境类型的贮点密度差异显著(F=7.471,P<0.05)。星鸦贮藏最密集的生境为人工针叶林,平均每个样方有14个贮点,贮藏密度为(3.48±0.62)个/m2;其次为次生林(1.35±0.30)个/m2;云冷杉林(1.20±0.22)个/m2;林间空地(0.93±0.20)个/m2;原始红松林样地内贮点最少,平均每个样方仅有0.6个贮点,贮点密度为(0.15±0.07)个/m2(图4a)。
不同基质类型的贮点密度有显著差异(F=3.205,P<0.05),其中基质为草地的贮点密度最高,为(3.94±1.37)个/m2,共发现212个贮点,占所有贮点的46.09%;基质为苔藓的次之,贮点密度为(1.74±0.44)个/m2;裸地的贮点密度为(1.63±0.34)个/m2;枯草地的贮点密度最低,为(0.93±0.20)个/m2(图4b)。
此外,基质硬度对贮点密度也有显著影响,基质较软时,星鸦贮藏密度更大(F=31.446,P<0.05)。
2.3 临时贮点管理
在球果堆放期间,星鸦对贮藏地的贮点更新比较频繁,每天都有新增贮点出现,也有贮点被重取。临时贮点的更新率很高,为48.34%;贮点重取率也很高,为45.23%,仅有6.43%的贮点没有发生变化。
人工针叶林的贮点更新率最高,为58.17%;林间空地、次生林的贮点重取率较高;云冷杉林的贮点留存率相对较高。原始红松林发现的贮点很少,且没有新增贮点(表3)。
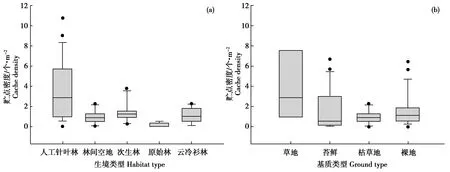
图4 不同生境类型和基质类型的贮点密度比较Fig.4 The cache densities in different habitat types and ground types
表3 不同生境类型中星鸦临时贮点的管理
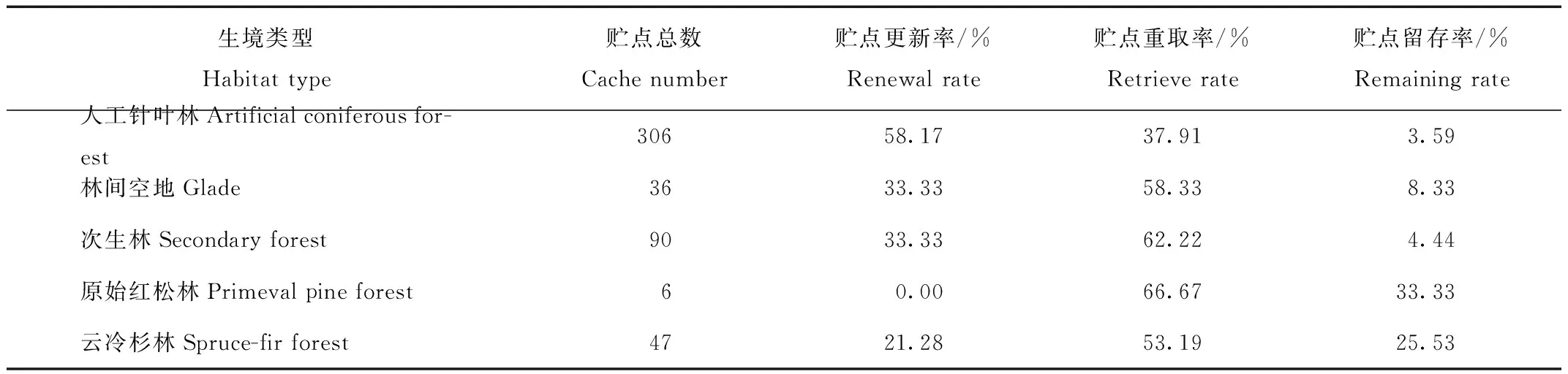
Tab.3 Temporary cache management of spotted nutcrackers in different habitats
星鸦重取贮点大小为(2.69±0.15)粒,留存贮点大小为(2.63±0.09)粒,临时新增贮点大小为(2.13±0.09)粒。留存贮点与新增贮点大小差异显著,新增贮点较小(F=1.471,P<0.05)。贮点重取与贮点大小显著相关(F=5.492,P<0.05),贮点越大,越易被重取。
3 讨论
3.1 星鸦贮食行为对集中食物源的适应
贮食行为是一种特化的取食行为。动物通常在食物资源丰富的时候贮藏食物,这种行为保证了动物在食物缺乏或在强烈的竞争条件下能够获得足够的食物,减缓食物缺乏季节所带来的困难。凉水自然保护区的红松球果集中堆放,成为星鸦的集中食物来源,本研究中星鸦对集中食物源的适应性行为主要包括在球果堆附近大量聚集、快速分散食物、频繁重取、更新贮点等。贮藏地星鸦的贮藏和重取行为时间比例为66.5%,其中重取时间比例高于贮藏行为。星鸦的贮食行为节律中,上午的贮藏行为时间比例较高,下午的重取行为时间比例较高。贮藏和重取行为有一定连贯性,贮藏在前,重取在后。
星鸦在食物源附近形成了大量临时贮点,并将临时贮点的种子重新取出,进行二次贮藏。因此在食物源附近适宜的贮藏地,星鸦对临时贮点重复利用,活动频次急剧增加,红外相机拍摄到多只星鸦在同一个样方内贮藏和翻取红松松子,这也导致贮食行为中出现了争斗行为,此行为在其他研究中很少有报道。出现在星鸦临时贮点的其他动物物种有松鼠、花鼠、普通,其中松鼠会盗取星鸦的贮点。
自然环境中星鸦的冬季重取期从11月初到次年1月[30]。而本研究中的贮点在11月初就全部被重取了,说明星鸦对集中食物源的重取行为与自然状态下完全不同,只是一种机会主义的分散贮藏行为,随着食物源消失,星鸦的活动频率、贮藏和重取强度均急剧下降,直到11月6日监测样地已无星鸦出现。
3.2 星鸦贮点生境选择
本次研究结果显示星鸦对贮点生境具有选择性,不同的生境类型中贮点的数量差异显著。凉水自然保护区的松塔堆位于管理局旁边的小路上,因此集中食物源周围生境较为复杂且人为干扰较大,但本研究结果表明星鸦的贮食行为受人为干扰影响很小,食物源距离是影响了星鸦对贮点生境的选择的最主要因素。在距离最远的原始红松林样方内,星鸦贮点最少,仅发现6个贮点、21粒红松种子,其中有2/3贮点被重取,且未发现新增贮点,所以可判定原始红松林不是星鸦倾向选择的临时贮藏地。
研究表明星鸦贮藏种子的传播距离较远,且随着种子大小而变化,种子越大传播距离越远[33-35]。加州星鸦(Nucifragacolumbiana)可将美洲矮松(Pinusedulis)种子传播至距母树远22 km的地点[36],星鸦搬运红松种子距离可达4 km[28]。但在本次调查中,发现星鸦的贮点距离人工采集红松球果堆较近,尤其是距离食物源不足5 m的人工针叶林林下,发现很多星鸦贮点,这符合短距离搬运、短时间分散的快速撤离假说,是对大量集中食物源的适应。人工针叶林地表植被为草地和苔藓,基质较软适合贮藏。鸦科(Corvidae)鸟类通常将单枚种子贮藏在开阔地或灌丛边,很少见其穿过浓密的灌丛,将种子贮于灌丛下[30,34]。本研究发现,星鸦多选择人工针叶林林下贮藏,树木分布疏松且林下无低矮灌丛,适合短距离低飞往返食物源处搬运种子。
星鸦优先选择在其家域范围内建立多个贮藏点,每个贮点内贮藏少量食物[33]。研究表明星鸦的贮点平均大小为(3.21±0.22)粒,深度(2.73±0.15)cm,密度(406±55)个/hm2[30]。本研究发现星鸦快速分散的临时贮点较小、密度高、聚集分布,临时贮点多为2—3个贮点一组,贮点之间距离较近,星鸦将一次携带的种子在相距不远处先后贮藏,方便记忆和取回。
猜你喜欢
格言·校园版(2022年17期)2022-07-06 22:01:48
小哥白尼(野生动物)(2021年9期)2022-01-17 02:31:00
中国林副特产(2020年5期)2020-10-09 08:24:14
广西林业科学(2019年2期)2019-07-25 02:28:46
中国林副特产(2019年3期)2019-01-10 17:17:11
福建林业(2018年3期)2018-11-07 05:34:36
防护林科技(2017年7期)2017-09-01 01:08:43
现代营销(创富信息版)(2016年6期)2016-11-21 06:00:03
现代营销(创富信息版)(2016年12期)2016-02-08 02:32:08
满族文学(2014年5期)2014-09-22 23:50:36
