
鸟类SLC2A4基因及其编码蛋白GLUT4分子进化分析
2020-04-22 09:38李清月刘晓颖
生物学杂志 2020年2期
刘 刚, 李清月, 汪 冲, 刘晓颖
(安徽医科大学 生命科学学院, 合肥 230032)
动物的生命活动都需要大量的能量,不同动物虽然食物来源不同,但在体内都需要先转化为葡萄糖来提供能量[1-2]。细胞对葡萄糖的摄入需要借助细胞膜上的葡萄糖转运蛋白(Glucose transporters,GLUT)才能得以实现[3-5]。迄今,被鉴定出来的GLUT共6种,即GLUT1~5和GLUT7,葡萄糖转运蛋白-4(glucose transporter type 4,GLUT4)是骨骼肌吸收和利用葡萄糖的关键蛋白,也是目前研究较多的葡萄糖转运体蛋白家族成员之一。GLUT4 主要分布于骨骼肌细胞中,是骨骼肌吸收和利用葡萄糖的关键蛋白,与骨骼肌的收缩密切相关,该蛋白能够在信号通路的调控下转移到细胞膜表面,参与骨骼肌细胞跨膜主动吸收葡萄糖[5-10]。GLUT4是由包含11个外显子的SLC2A4基因进行编码,研究表明,SLC2A4基因敲除的小鼠会出现严重的胰岛素抵抗以及葡萄糖耐受等不良症状,表明GLUT4在动物的骨骼肌葡萄糖吸收、血糖稳态维持以及骨骼肌运动等方面发挥极其重要的作用[11-14]。
鸟类飞翔时需要消耗骨骼肌产生的大量能量,因此,鸟类不但需要高效的血糖调节机制,而且还需要一个能够快速将葡萄糖转运到骨骼肌细胞内的重要介导蛋白[15-16]。大量的生理学研究表明,GLUT 4是鸟类骨骼肌细胞吸收葡萄糖的最主要转运蛋白,从而为鸟类的飞行活动提供能量[17-18]。因此,鸟类在飞翔中的高耗能活动,能够促使GLUT 4更加活跃的转运葡萄糖。研究表明,SLC2A4基因及其表达产物GLUT4在哺乳动物中发生了强烈的进化选择,与其他动物相比,鸟类飞翔需要高的能耗,我们推测该基因在鸟类中极可能发生了适应性进化,同时我们推测SLC2A4基因及其表达产物GLUT4在善于飞翔的鸟类和不善于飞翔的鸟类中可能发生不同的适应性进化[19-20]。本文选取了NCBI上已经公开发表且注释较完善的17种鸟类SL2A4基因及其编码的GLUT4作为研究对象,通过生物信息学的方法对该基因的结构、特征、基因进化速率及系统发育关系进行分析,探讨该基因在不同鸟类类群中是否发生了适应性进化,为鸟类SL2A4基因和GLUT4的进一步研究提供有用的信息。
1 材料与方法
1.1 研究对象的选取及数据来源
为了研究SLC2A4基因及其表达产物GLUT4在鸟类中的适应性进化,同时结合GenBank上已经释放的注释完整的数据,选取了今鸟亚纲(Neornithes)中的突胸总目(Carinatae)和平胸总目(Ratitae)鸟类为研究对象(表 1)。研究中的突胸总目主要包括Anseriformes、Columbiformes、Passeriformes和Galliformes等4目14种鸟类,推测该类群鸟类具有较为高效的GLUT4转运效率。该类鸟类的主要特征为善于飞翔,具有与飞行相关的生理结构特征:具充气性骨骼(气质骨),最后4~6枚尾椎骨愈合成一块尾综骨,胸骨具有龙骨突起结构,具有适宜于飞行的发达的骨骼肌,肋骨上有钩状突起;翼发达、正羽发达、羽小枝上具小钩,构成羽片。本研究中的突胸总目主要包括Apterygiformes、Struthioniformes和Tinamiformes等3目3种鸟类,推测该类群鸟类的GLUT4转运效率较低。该类鸟类主要特征不善于飞翔,生理结构特征主要为翼退化、胸骨不具龙骨突起,不具尾综骨,羽毛均匀分布、羽枝不具羽小钩,不形成羽片。平胸总目的著名代表为Apteryxrowi、Dromaiusnovaehollandiae和Nothoproctaperdicaria。本研究中所有物种拉丁名参照整合分类信息系统 ITIS(Integrated Taxonomic Information System,www.itis.gov)。下载GenBank(www.ncbi.nlm.nih.gov)迄今为止已释放且注释较完善的17种鸟类SLC2A4基因序列及其编码的GLUT4(表 1)。由于SLC2A4基因及其表达产物GLUT4在模式生物小鼠(Musmusculus)内研究较为透彻,另下载小鼠相应序列(NM_009204)作为参考进行比较研究。
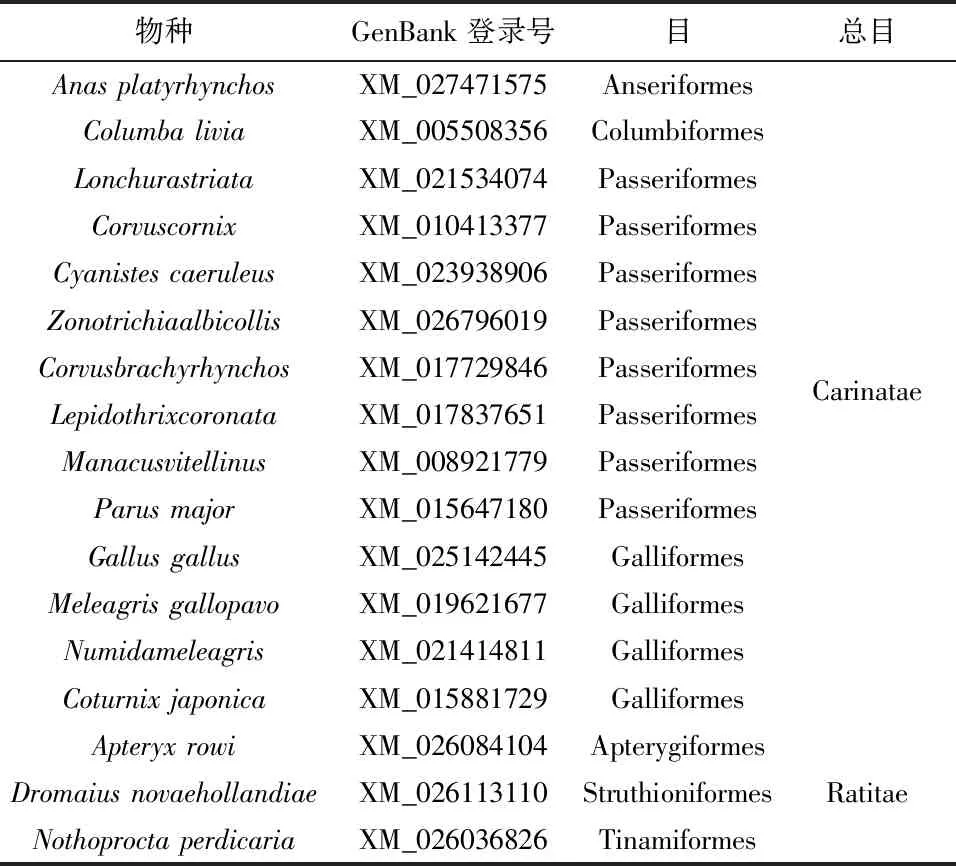
表1本研究物种来源及SLC2A4基因序列的GenBank登录号Table 1 Species examined and GenBank Accession No. of SLC2A4 in this study
1.2 SLC2A4基因序列和GLUT4分析
对从GenBank中下载的SLC2A4基因全长mRNA使用MEGA 7.0分析其碱基组成;GLUT4的理化性质主要利用ExPASy在线软件(http://web.expasy.org/protparam/)进行分析,通过SignalP4.1 Server软件(http://www.cbs.dtu.dk/services/SignalP/)预测信号肽位点;GLUT4的疏水性/亲水性通过ExPASy服务器的ProtScale程序中的K-D法(http://web.expasy.org/protscale)进行分析;通过GORIV在线分析平台(https://npsa-prabi.ibcp.fr)预测其二级结构,蛋白质糖基化位点利用在线分析平台(http://www.cbs.dtu.dk/services/NetNGlyc/)进行分析。
1.3 进化速率分析及系统发育树的构建
采用PAML软件包对SLC2A4基因进行选择压力分析。使用PAML4软件中的CODEML程序,采用基于密码子(codon-based)的分析方法计算非同义替换率(nonsynonymous substitution,dN)和同义替换率(synonymous substitution,dS)的比率(ω,dN/dS)。ω值是检测自然选择的重要参数,如果ω=1,基因则受中性进化(neutral selection),ω<1时基因受纯化选择purifying selection),ω>1则认为基因受正选择(positive selection)。模型的估算输入的物种树参考相关鸟类系统关系资料,我们选用了以下几种模型进行选择压力分析:1)用free-ratio模型估算进化树上每一枝的ω值;2)用two-ratio模型估算前景枝Gallusgallus的ω值(ω1)和背景枝除Gallusgallus外的所有枝的ω值(ω0)。最后对两种模型进行似然率(Likelihood Ratio Test,LRT)检验用以判定该模型的统计方法是否具有显著性。LRT需要用free-ratio与one-ratio模型、two-ratio与one-ratio模型的似然值之差的二倍去查询卡平方表来判定是否推翻零假设(null)而接受备则假设(alternative)。
为了更好地分析SLC2A4基因在鸟类中的分子进化,以小鼠作为外群,以SLC2A4基因编码的GLUT4作为分子标记构建17种鸟类的系统发育树。具体方法如下:把所有GLUT4序列用CLUSTALX 2.1进行比对,用MEGA 7.0软件转变成进行修剪并转换成“.nex”格式文件进行数据模型选择,最终选取AIC模式下的模型应用于系统树的构建。采用PAUP 4.10软件构建最大似然法(ML树)系统树,建树前先使用Modeltest v 3.7选择最优模型,然后将所得最优模型写入PAUP 4.10软件命令行中,建树执行快速支持度分析并在一次单独的程序运行中搜索最佳分值的ML树的方法,自举检验100次。贝叶斯推论法构建系统树(BI树)用MrBayes 3.1.2软件建立,建树前先使用Modeltest v3.7选择最优模型,运行4个马尔可夫链,每100代抽样1次,运行100万代,去除老化样本。
2 结果与分析
2.1 SLC2A4基因的核苷酸序列分析
17种鸟类SLC2A4基因序列长度在990~1719 bp之间,平均长度为1446 bp,其中最短的为Parusmajor,最长的为Coturnixjaponica(表2)。SLC2A4基因的AT含量在26.0%~53.3%之间,GC 含量在46.7%~74.0%之间(表2)。
2.2 GLUT4分析
17种鸟类的SLC2A4基因编码的GLUT4进行分析,其长度在329~572 AA之间,平均长度为481 AA,其中最短的是Parusmajor,最长的为Coturnixjaponica。GLUT4序列信号肽剪切位点在氨基酸序列的第11位与21位之间,Gallusgallus位于第48位,而Parusmajor位于第57位。在17种鸟类中,GLUT4分子质量在 35 424.4~59 310.9之间,平均分子量为52 550.89;等电点在5.09~9.58之间,平均等电点为6.53;不稳定指数在38.52~79.99之间。在鸟类中所有GLUT4分子为亲水性蛋白,总平均疏水性-0.54~-0.285之间(表3)。通过对GLUT4蛋白分子糖基化位点分析表明,N-糖基化个数在1~2之间,而O-糖基化个数在29~50之间(表4)。通过对GLUT4二级结构预测,发现17种鸟类蛋白质分子均不存在跨膜结构(图1)。
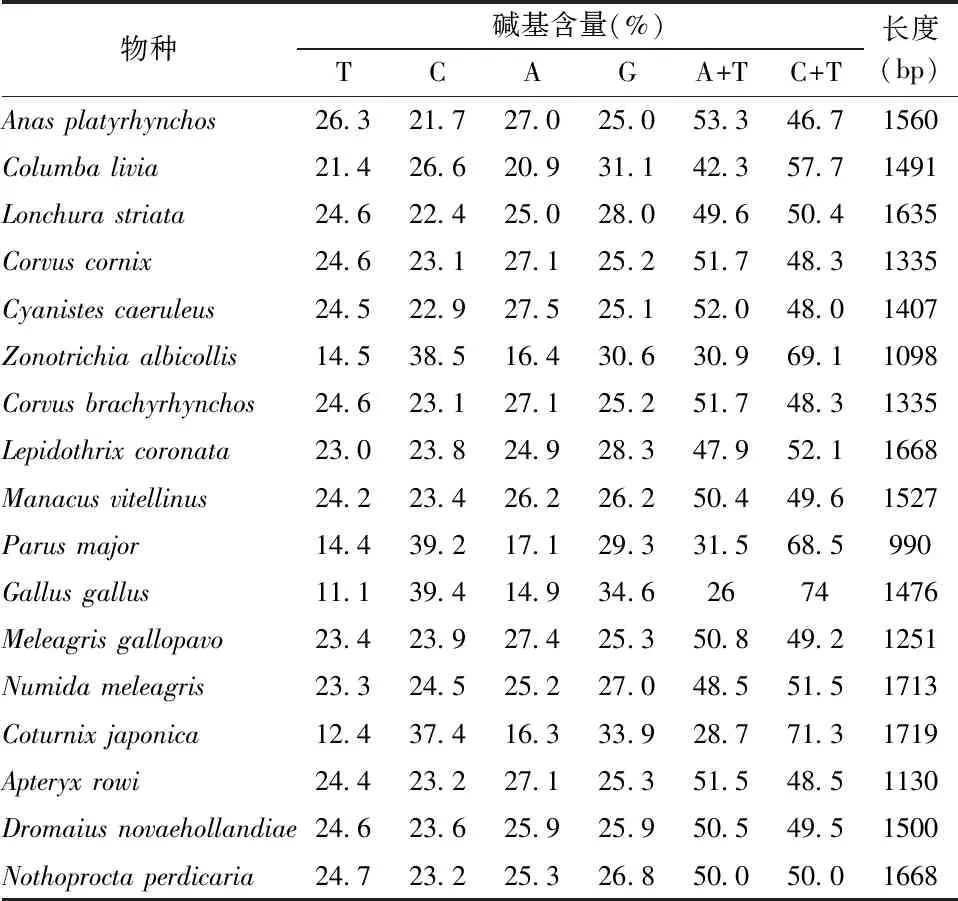
表2 17种鸟类SLC2A4基因碱基含量分析Table 2 Nucleotide composition (%) of SLC2A4 among 17 birds
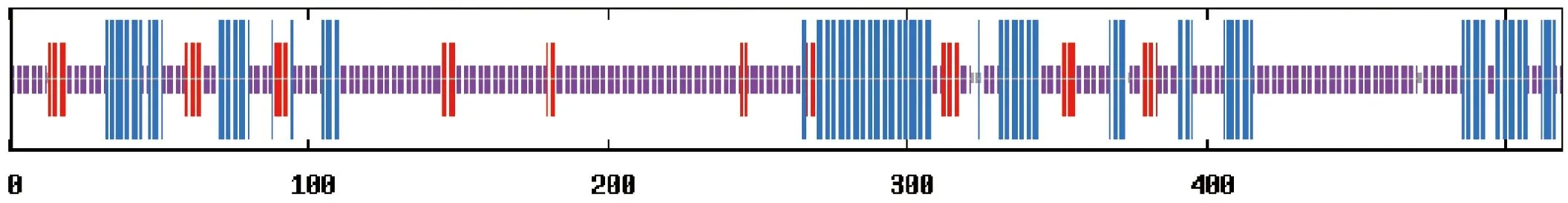
蓝色为α螺旋结构,红色为β片层结构,紫色为无规则卷曲结构(以Anasplatyrhynchos为例)
图1 GLUT4二级结构
Figure 1 Secondary structure of GLUT4
2.3 进化速率和系统发育分析
通过对编码GLUT4的SLC2A4基因的进化压力分析发现,14种突胸总目(Carinatae)鸟类ω值1.195,具有明显的正选择效应;而3种平胸总目(Ratitae)鸟类ω值0.433,有纯化选择作用;17种鸟类的ω值为0.685;使用PAML程序包对选择压力进行分析,均未检测到正选择位点。使用Datamonkey的SLAC算法,17种鸟类GLUT4找到4个正选择位点,46个负选择位点(P=0.01)。
研究中系统发育树图的枝长和拓扑结构源于贝叶斯推断法树图,通过构建GLUT4系统发育树。结果显示系统发育树分为两枝:Ratitae分为第一枝,包括Struthioniformes、Apterygiformes和Tinamiformes 3个目; Carinatae构成第二枝,即Galliformes、Anseriformes、Columbiformes和Passeriformes(图2)。
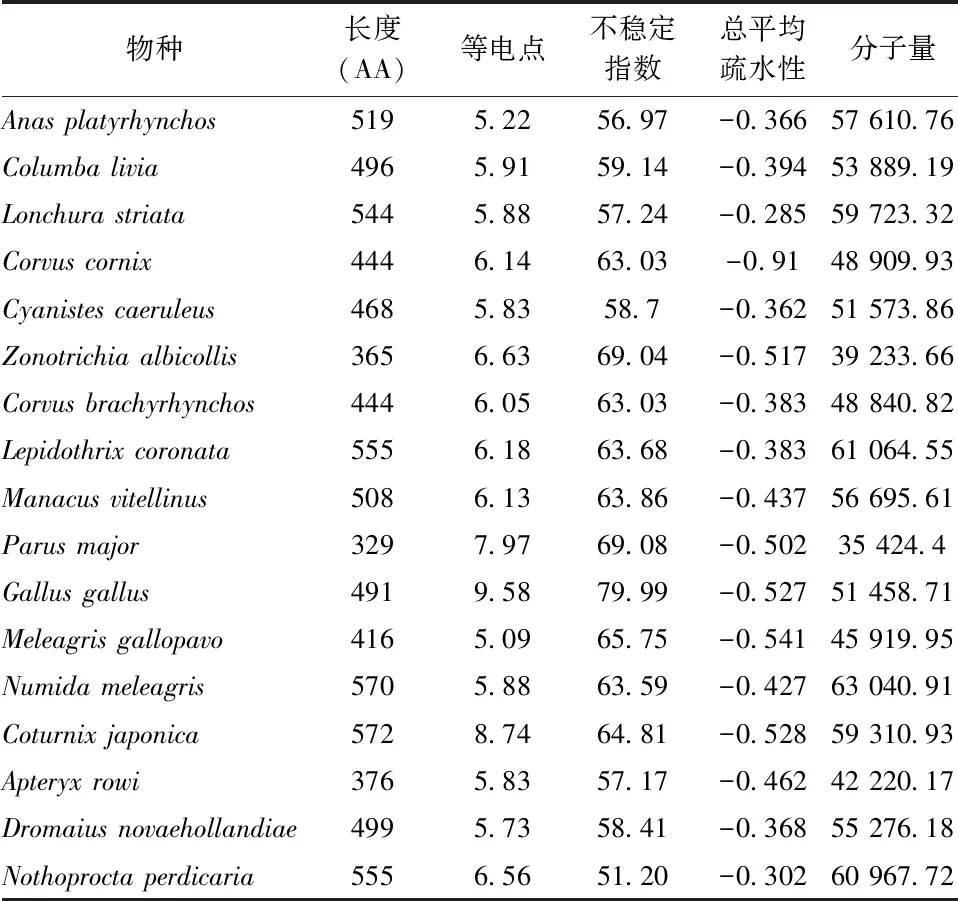
表3 GLUT4的氨基酸理化性质分析Table 3 Physicochemical property of GLUT4
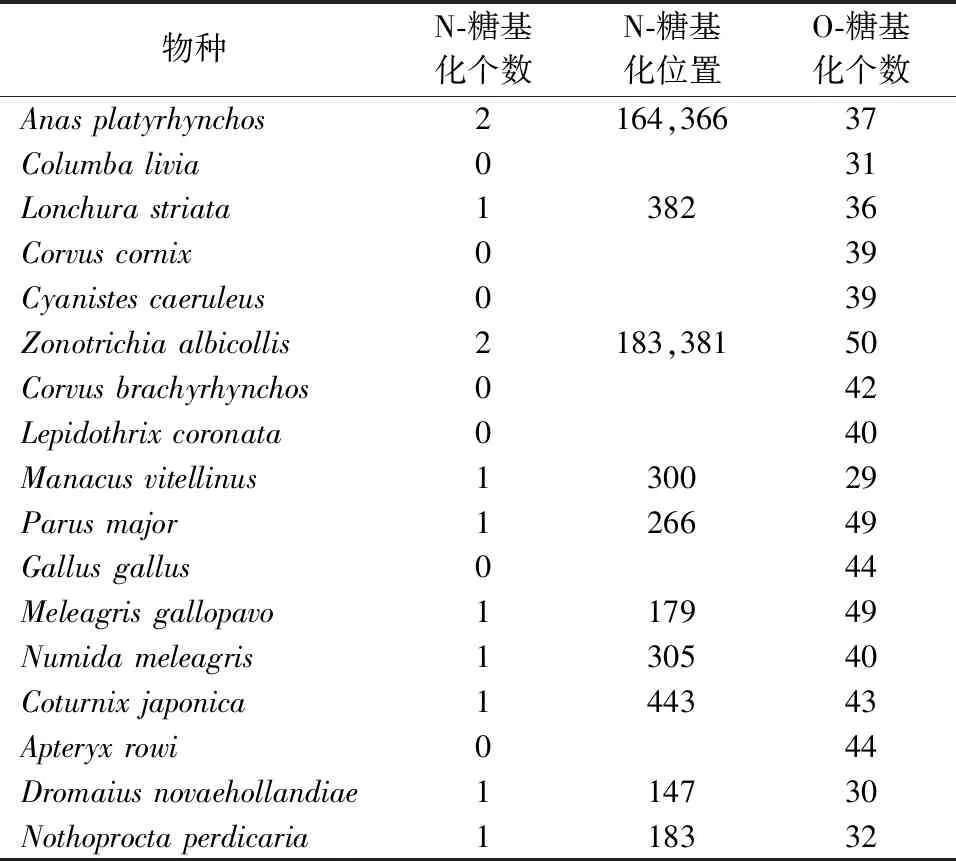
表4 GLUT4的糖基化分析Table 4 Glycosylation analysis of GLUT4
3 讨论
SLC2A4基因及其编码蛋白GLUT4是细胞转运葡萄糖的主要转运体,是维持机体葡萄糖稳态的关键因素,该蛋白质是细胞在胰岛素刺激的情况下发生的急性反应,主要表达在脂肪、心肌和骨骼肌等对胰岛素相对敏感的组织中,与骨骼肌细胞的能量产生具有重要作用[3,22]。研究表明,敲除小鼠的SLC2A4基因会出现严重的胰岛素抵抗和葡萄糖耐受不良等症状,表明该基因在动物血糖稳态维持等方面发挥极其重要的作用[11-12,21]。通过对SLC2A4基因的长度、碱基变异分析,发现在鸟类中该基因具有较高的变异率。本研究的结果发现该基因在平胸总目和突胸总目鸟类中发生了明显的适应性进化,平胸总目鸟类中该基因选择压力ω<1,表明该基因选择压力为纯化选择作用,可能是由于与该总目的鸟类不善于飞翔,骨骼肌耗能较小,因此只需要较少的葡萄糖相关。在对突胸总目鸟类中,该基因在进化过程中发生了显著的选择压力的变化,即ω>1,表明该基因经历明显的正选择作用,这可能与该总目鸟类普遍善于飞翔,需要大量的能量,因此骨骼肌细胞需要吸收大量的葡萄糖来维持能量的需求。研究结果表明,在不同的物种中GLUT4均具有明显的糖基化修饰,通过改变其空间结构增加蛋白质的稳定性[23-24],本研究对鸟类的GLUT4研究发现,尤其是在突胸总目鸟类中均具有大量的糖基化位点,这些糖基化位点为能够加速葡萄糖的转运,推测可能与鸟类飞翔时需要大量的能量有关。通过对GLUT4蛋白分子二级结构预测,发现17种鸟类蛋白质分子均不存在跨膜结构,与小鼠蛋白存在明显区别[25-26]。
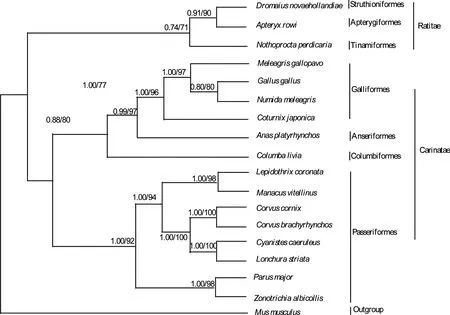
图2 基于GLUT4序列构建的系统发育树
ML和BI系统发生树分为Ratitae和 Carinatae两枝,且获得了较高的支持率,与形态学分类结果相吻合,表明该系统树均能够很好地反映物种进化的历史[27-28],这说明SLC2A4 基因编码的GLUT4蛋白具有较强的系统发育意义,可以作为研究鸟类进化的理想分子标记。
猜你喜欢
华东师范大学学报(自然科学版)(2021年6期)2021-01-01
能源研究与信息(2020年4期)2020-02-19
癌症进展(2018年11期)2018-12-30
天然产物研究与开发(2018年2期)2018-04-04
中国运动医学杂志(2016年3期)2016-07-10
中国运动医学杂志(2016年3期)2016-07-10
医学研究杂志(2015年7期)2015-06-22
医学研究杂志(2015年12期)2015-06-10
医学研究杂志(2015年5期)2015-06-10
中国中医药图书情报(2014年6期)2015-03-30
