
长白山阔叶红松林乔木幼苗组成及多度格局的影响因素
2020-04-20 13:22吴兆飞李明鲁张春雨赵秀海
生态学报 2020年6期
邱 华,舒 皓,吴兆飞,李明鲁,殷 正,张春雨,赵秀海,*
1 北京林业大学国家林业和草原局森林经营工程技术研究中心,北京 100083 2 北京林业大学理学院,北京 100083
幼苗阶段是植物生活史中生长最脆弱、对环境反映最敏感的时期[1]。在森林群落中,无论是种群水平还是群落水平,林下层幼苗的动态特征随时间、空间的变异性非常大,幼苗更新状况最终会决定森林生态系统的群落组成结构以及演替进程。影响幼苗更新的因素主要包括草本植物、林分条件、物种组成等生物因子[2-4],以及林冠盖度、土壤水分、坡度、坡向等环境因子[5- 6]。
随着森林固定监测大样地的发展,幼苗监测作为森林样地监测的一个重要组成部分,越来越被重视[7-9],结合固定监测样地,国内外学者对幼苗更新的研究主要包括幼苗物种组成及其空间分布[10- 13]、短期动态变化[14- 15],以及幼苗存活的影响因素[16- 18]等。
阔叶红松林作为长白山地带性顶级植被群落,林分内物种丰富度高、建群种独特、林下植被资源丰富、草本多样性高,一直是我国温带森林生态系统研究的重点[6,19]。对群落乔木幼苗组成及多度格局影响因素的研究将进一步完善我们对温带森林群落维持物种多样性机制的认识。有关阔叶红松林的研究已有一些[4,20- 23],而综合探讨生物和非生物等多种因素对幼苗多度格局影响的研究还相对较少。
综上所述,本研究通过分析影响长白山原始阔叶红松林乔木幼苗多度格局的生物邻体和生境因子,以期解决以下问题:1)乔木幼苗的物种组成及分布特征;2)生物因子(生物邻体及草本效应)和环境因子(土壤含水量及林冠开阔度)如何影响幼苗多度?3)在群落水平和物种水平下,生物及环境因子对幼苗多度的影响有何不同?
1 研究地区与研究方法
1.1 研究样地概况
研究区位于吉林省长白山国家级自然保护区原始阔叶红松林内。该地区属于温带大陆性气候,冬季寒冷多风,夏季温暖湿润。年均温2.9℃,年均降水量在600—900 mm。土壤类型为山地暗棕色森林土。群落植被主要包括红松(Pinuskoraiensis)、紫椴(Tiliaamurensis)、水曲柳(Fraxinusmandschurica)、蒙古栎(Quercusmongolica)等[24]。
1.2 研究方法
1.2.1样地设置和调查
图1 幼苗样方示意图Fig.1 Sketch map of the seedling plots
2014年,根据美国热带森林科学中心(Center for Tropical Forest Science,CTFS)样地建设标准和操作规范,选择典型的原始阔叶红松林,应用全站仪,建立面积40 hm2(500 m × 800 m)的固定监测样地(图1)。样地坐标为42°20′ N、127°54′ E,海拔975—997 m。调查样地内所有胸径(diameter of breast height,DBH)≥1 cm的木本植物,记录其种类、胸径、树高、冠幅、枝下高和空间坐标等,并挂牌标记。
1.2.2乔木幼苗和草本调查
2018年,按机械布点的方式在样地内每隔40 m均匀设置228个幼苗样方(2 m×2 m),7—8月每月调查一次幼苗样方内所有胸径<1 cm的乔木个体,记录种类、数量及坐标等,并挂牌标记。同时,调查幼苗样方内所有的草本植物,记录样方内草本种类、数量、均高、盖度和总盖度。其中草本盖度的测量方法是找出该幼苗样方中能代表某种草本生长平均状态的一株草本,用卷尺测量出该草本平行于幼苗样方两边的最大边长,然后数出该种草本在该幼苗样方的株数,即可得到该种草本的草本盖度。
1.2.3生物邻体变量
生物邻体主要考虑大树邻体。大树邻体(A)是以目标幼苗为中心、半径15 m范围内的所有同种或异种大树的胸高断面积除以其与目标幼苗的距离之和[25]。
式中,BAi是第i棵大树的胸高断面积; DISTANCEi是目标幼苗与第i棵大树的距离;N是以目标幼苗为中心、半径15 m范围内的同种或异种大树个体数。
1.2.4生境变量
生境变量主要包括林冠开阔度和土壤含水量。研究样地地势平坦,没有沟渠、山脊、山谷等特殊地形,因此本研究没有考虑地形对幼苗的影响。林冠开阔度用叶面积指数(leaf area index, LAI)表示,LAI指单位土地面积上植物叶片总面积占土地面积的比例,LAI值越大,林冠开阔度越低[26- 27]。2018年7月份阴天条件下用冠层分析仪(WinSCANOPY, Quebec, Canada)在每个幼苗样方内距离地面1.3 m处各进行一次测量,用FV2200软件处理数据,得出每个幼苗样方的LAI值。同时用手持土壤水分测定仪(HH2Delta-T Devices Moisture Meter, Delta-T, Cambridge, UK)在每个幼苗样方中随机选取3个点,测定表层土壤水分含量,取平均值作为该幼苗样方的土壤含水量。
1.3 数据处理
采用样地胸径≥1 cm木本植物的调查数据、2018年7和8月份228个4 m2样方的幼苗及草本调查数据,以及林冠开阔度指标和土壤含水量指标进行数据分析。
运用广义线性混合效应模型(GLMMs),以7月和8月份两次调查中每个幼苗样方所有胸径≤1 cm的存活幼苗的幼苗多度作为因变量,分析其与生物和生境变量之间的关系,在模型中,将生物因子和环境因子作为固定效应,样方号及物种作为随机效应。所有连续型自变量在进入模型之前都进行标准化处理,即该值减去平均值后除以标准偏差。
运用模型对群落水平(群落中所有物种)和物种水平(两次调查中物种多度排名前5的物种,包括水曲柳、紫椴、假色槭、红松和簇毛槭)上影响幼苗多度格局的因素进行了分析。本研究中所有分析与绘图采用Microsoft Excel 2016和Visio 2013及国际通用软件R 3.5.3(http://www.r-project.org), 其中‘lme4′软件包完成GLMM的分析。
2 结果与分析
2.1 物种组成及数量特征
2018年7月份调查到乔木幼苗6377株,8月份调查到存活及新萌发幼苗3687株,两次累计调查到17个物种,分属9科9属(表1)。其中,蒙古栎、鱼鳞云杉和怀槐只在8月份调查到。两次调查中,水曲柳幼苗数量均在3000株以上,在4 m2的幼苗样方内,幼苗密度达到了(6.50±0.04)株/m2和(3.73±0.02)株/m2(n=228),在乔木树种幼苗组成中占据重要位置。个体数在两次调查中均小于10的物种有4种,占幼苗总数的0.114%。从幼苗的种频度来看,水曲柳、假色槭、紫椴、红松、小楷槭和簇毛槭幼苗分布相对广泛,其他树种幼苗分布相对较少。
2.2 幼苗多度的影响因素
2.2.1 群落水平
广义线性混合效应模型中用到的参数见表2。群落水平上,幼苗多度与异种大树效应、草本密度和林冠开阔度呈显著正相关,与草本盖度、土壤含水量呈显著负相关,同种大树效应对于幼苗多度的影响不显著。由于调查到的幼苗多度之间存在显著差异,把群落中多度较低的幼苗从模型中筛除后,幼苗多度与同种大树效应呈显著负相关(图2)。
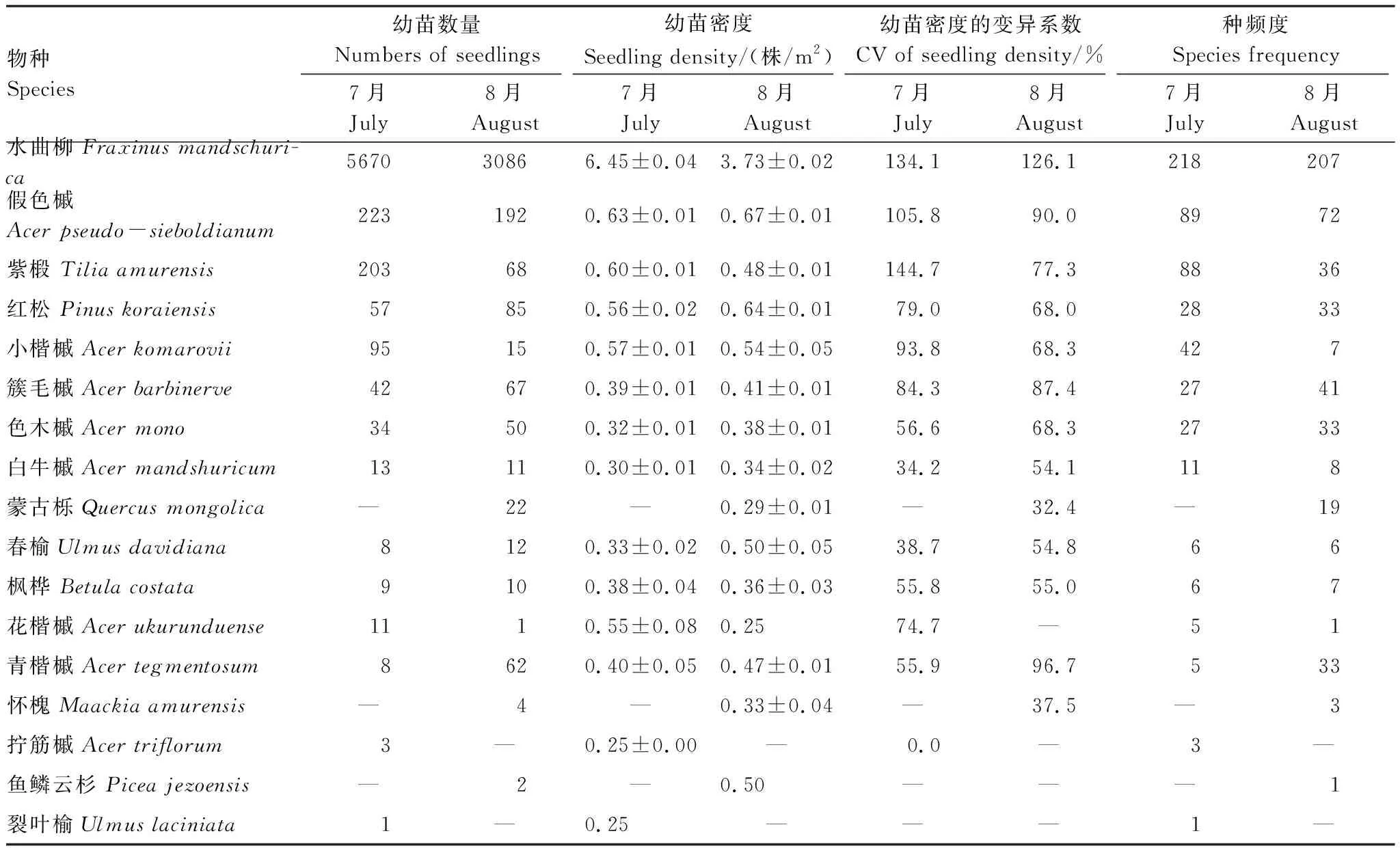
表1 幼苗物种数量组成
种频度指幼苗在幼苗样方中出现的次数
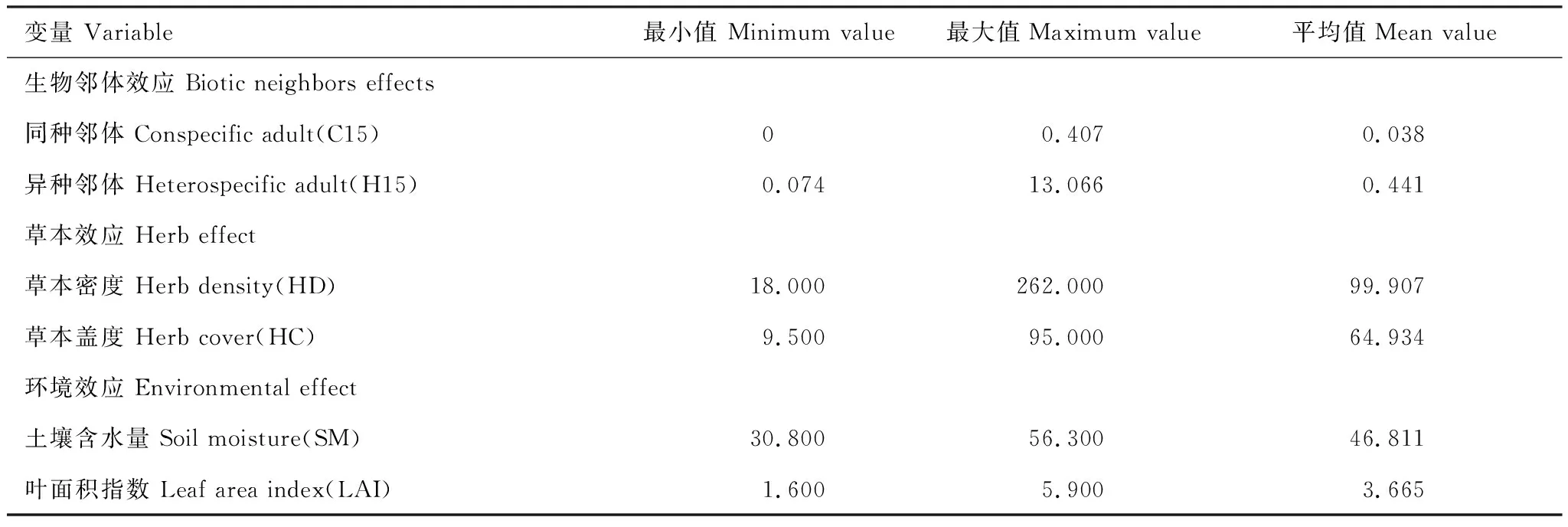
表2 广义线性混合效应模型中使用的参数
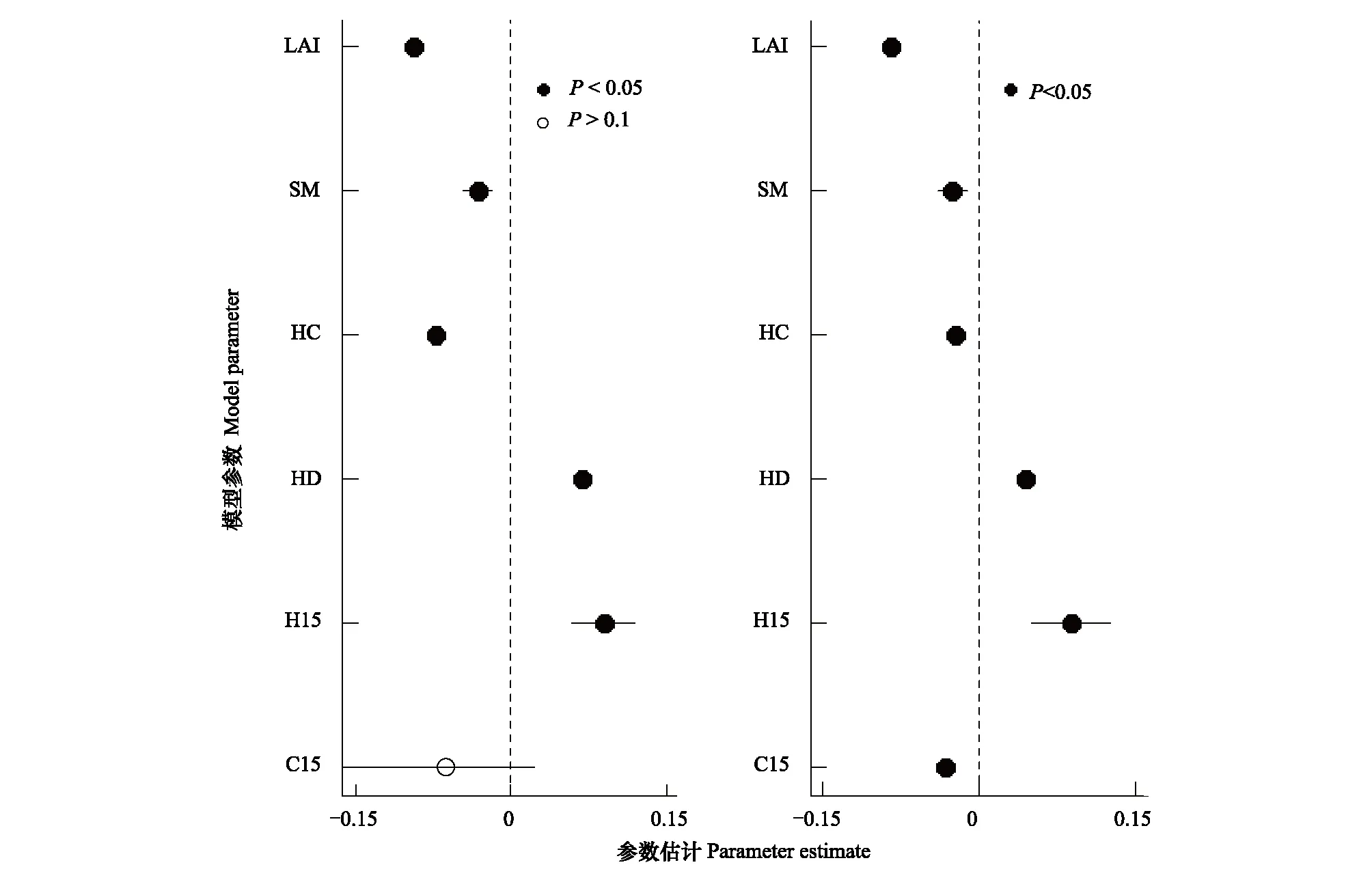
图2 群落水平上模型各参数估计值Fig.2 Estimates of the coefficients of the models at the community level (mean±2SE)左侧为所有种的结果, 右侧为筛选后的结果 LAI: 叶面积指数 Leaf area index; SM: 土壤含水量 Soil moisture; HC: 草本盖度 Herb cover; HD: Herb density; H15: 15 m范围内的异种大树效应 Heterospecfic adult effect within 15 m; C15: 15 m内的同种大树效应 Conspecific adult effect within 15 m
2.2.2物种水平
物种水平上,通过对两次调查中多度排名前5的物种分析发现,各因子影响幼苗多度的相对重要性在物种间存在显著差异。水曲柳和紫椴幼苗多度的影响因素与群落水平一致。红松幼苗多度与异种大树效应呈正相关,与同种大树效应和林冠开阔度呈负相关(图3)。簇毛槭及假色槭与文中所测的所有生物因素和非生物因素均无显著相关性(图4)。
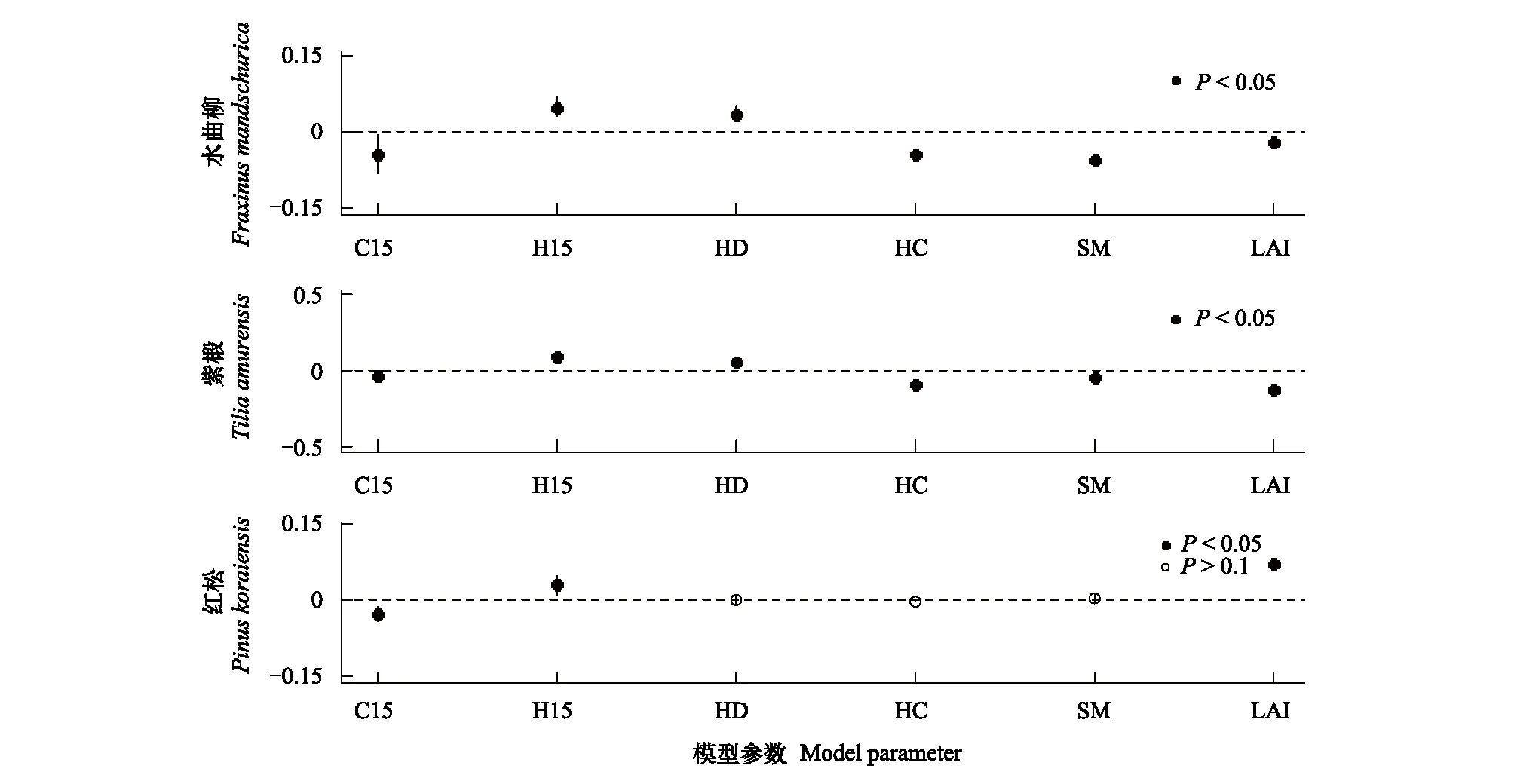
图3 显著物种各参数估计值Fig.3 Estimates of the coefficients of the significant species (mean±2SE)
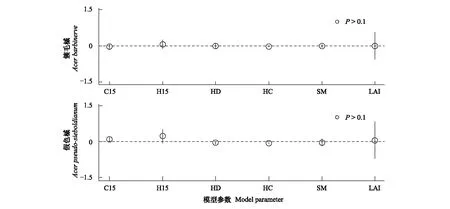
图4 不显著物种各参数估计值Fig.4 Estimates of the coefficients of the non-significant species (mean±2SE)
3 讨论
3.1 群落的物种组成及分布
2018年7和8月份对长白山40 hm2原始阔叶红松林样地内228个幼苗样方的幼苗调查,共调查到17个物种,这些物种都是群落乔木层的主要优势物种,幼苗物种组成与样地乔木大树物种组成一致[15,28]。但仍然有许多树种的幼苗没有调查到或者数量极少,如蒙古栎、枫桦等。宾粤等[12]认为这可能是由于样方周围许多树种个体胸径较小还未繁殖,或者有些树种虽胸径较大,但因种子扩散限制的作用,从母株掉落的种子未能达到幼苗样方内,从而导致调查到的幼苗个数极少。另外,某些树种是许多野生动物取食的主要对象,以及幼苗受自然天敌的影响等,均是导致幼苗较少的原因[29- 30]。
幼苗数量变化很大且在各样方表现出明显的数量差异,这可能是由于幼苗的大量生长及死亡导致的[31],相关原因仍需对种子雨动态及幼苗更新动态的影响因素做进一步调查研究。
3.2 不同水平上幼苗多度格局的影响因素
3.2.1群落水平
生物因素和环境因素共同影响幼苗的多度格局。在大树效应上,幼苗多度与异种大树效应呈显著正相关,与同种大树效应的关系不显著,当模型中去除多度较低的物种后,幼苗多度与同种大树效应呈显著负相关。这可能是由于样地中样本量较少幼苗的掩盖作用,使得整个群落中幼苗多度与同种大树效应关系不显著。Janzen-Connell假说认为种子扩散以母株为中心, 邻近母株的种子和幼苗存在较高的死亡率,本研究得出同种大树效应与幼苗多度呈显著负相关,即局域同种大树密度越大,由于宿主的专一性及捕食者的捕食,目标幼苗死亡率越高[32],从而导致幼苗多度的减少,这在一定程度上证明了Janzen-Connell假说。
在草本效应上,幼苗多度与草本盖度呈显著负相关,这是因为草本盖度在一定程度上限制了幼苗所需的光照条件和土壤养分,从而影响了幼苗更新。幼苗多度与草本密度呈显著正相关,可能因为在一定程度上,幼苗和草本对环境的需求相似,草本密度大的地方周围环境良好,从而也有利于幼苗多度的增加,这与Cao等[24]在长白山关于草本效应对幼苗存活影响的研究结果一致。另外,肖翠等[4]在长白山阔叶红松林幼苗存活因素分析的研究中没有发现草本效应的显著影响,可能是因为本研究草本量较大,使得群落水平上草本效应显著。
环境特性上,幼苗多度与土壤含水量呈显著负相关,表明土壤含水量在一定程度上可以限制幼苗的更新和生长。在2018年幼苗生长期的调查中,长白山雨多且雨期较长,土壤长期处于较湿的状态,使土壤含水量限制了幼苗的多度甚至存活。林冠开阔度与幼苗多度呈显著正相关,说明幼苗在林下光照条件较好的生境中多度较高,Montgomery等[33]得出过类似的结论。另外,光照条件在其他研究中同样被认为是一个显著限制植物更新的因子[5,34- 37]。
3.2.2物种水平
影响幼苗多度格局的生物和生境因素的相对重要性存在显著的种间差异。水曲柳和紫椴幼苗多度与异种大树效应、草本密度及林冠开阔度呈显著正相关,与同种大树效应、草本盖度和土壤含水量呈负相关,生物邻体及生境因素共同影响着幼苗多度。另外这两个物种幼苗多度的影响因素与群落水平一致,也证明了水曲柳和紫椴幼苗数量在所有调查的幼苗中占优势地位。红松幼苗多度与异种大树效应呈正相关,与同种大树效应及林冠开阔度呈负相关,这可能是因为红松幼苗及幼树耐阴性强,光照太强反而不利于幼苗生长。另外,簇毛槭和假色槭幼苗多度与任何因子均无显著关系,野外调查发现两种幼苗新生率较高且成年大树均为样地内的优势种,分布范围较广,这可能是两种幼苗多度和生物因子及环境因子关系都不显著的原因。
3.3 草本效应和环境因子
以往对于固定监测样地中草本的研究主要集中在草本的数量组成、年月际动态及与环境的关联性方面[38];也有文章在分析幼苗存活的影响因素中提到草本效应对于幼苗存活的影响[4]。而在大尺度的原始阔叶红松林中,分析草本植物对于乔木幼苗多度影响的研究相对较少。本研究将草本样方中的草本密度和草本盖度引入到幼苗多度的影响因素中,结果表明,草本植物对某些特定的幼苗多度有影响,比如草本盖度与水曲柳幼苗多度存在显著负相关关系(图3)。另外,环境因子中的林冠开阔度及土壤含水量很大程度地影响到幼苗的多度格局,从而制约幼苗存活。Cao等[24]在长白山原始阔叶红松林幼苗更新的研究中认为,土壤含水量对于幼苗的影响应该存在一个平衡点,平衡点的两侧显著度相反。因为本研究的取样时间以及某些物种的样本量有限,草本植物对某些幼苗多度的影响没有检测出来,同时,土壤含水量没有去找平衡点。以后将加大研究时间及采样次数,以期更准确地探讨草本植物和土壤含水量对于幼苗多度的影响。
4 小结
通过对长白山原始阔叶红松林木本植物幼苗2018年的两次调查发现,新生幼苗数量较多,但各物种间幼苗多度的差异较大。不论是群落水平还是物种水平,生物因子和环境因子共同影响幼苗的多度格局,并且两者间的相对重要性因物种而异。
猜你喜欢
林业资源管理(2022年6期)2023-01-18
林业勘查设计(2022年1期)2022-02-15
现代园艺(2021年23期)2021-12-01
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
青年文学家(2021年16期)2021-08-05
新农业(2020年18期)2021-01-07
花卉(2020年24期)2021-01-04
少儿美术(2020年8期)2020-11-06
现代园艺(2017年13期)2018-01-19
