
不同温度条件对3种车前属植物种子萌发的影响
2020-04-20 11:59:14贾风勤12
种子 2020年3期
贾风勤12
(1.乌鲁木齐职业大学旅游学院,乌鲁木齐 830002;2.乌鲁木齐职业大学旅游发展研究所,乌鲁木齐 830002)
在种子植物的生活史中,种子萌发是植物生长的基础,也是植物繁殖过程中的一个重要环节,萌发特征影响着幼苗的存活能力和个体适合度[1]。作为幼苗建成的前提,种子萌发甚至决定某些植物的稀有性和分布范围,深入分析种子萌发特点及其调控因子,不但可以明确其自然有性更新的原因,而且可以使人工控制种群更新成为现实[2]。
种子萌发受自身基因、生理和形态等属性影响[3-5],还与自然环境中非生物因子密切相关。温度是影响种子萌发的重要因素之一,适宜的温度可增强酶活性,促进物质和能量转化,种皮透性改变,利于种子吸水和气体交换。变温有利于种子萌发,如李雄等研究发现,紫花针茅(Stipapurpurea)种子适宜的萌发温度是变温15 ℃/25 ℃[6]。张颖娟等认为,变温15 ℃/25 ℃也同样有利于长叶红砂(Reaumuriatrigyna)和红砂(Reaumuriasoongorica)种子萌发[7]。而鱼小军等对条叶车前种子(Plantagolessingii)萌发研究认为恒温20 ℃最适宜该种子萌发[8];黄振英等也发现梭梭(Haloxylonammodendron)种子最适萌发温度为10 ℃[9]。刘有军等研究发现,恒温20 ℃和变温为15/25 ℃均为碟果虫实(Corispermumpatelliforme)萌发的最佳温度[10]。刘自刚等[11]在对桔梗的研究中也认为适宜水分条件下,变温10 ℃/20 ℃和恒温15~25 ℃条件下桔梗种子能够良好萌发。事实上,植物都以个性化的萌发模式适应着变化的环境,即使同一物种,产自不同地区的种群种子萌发特征也可能截然不同,如车前(Plantagoasiatica)[12]、紫花针茅[6]。所以,深入分析植物种子萌发特性有助于评估植物种群有性更新的适宜生态位,促进植物资源管理[13],有助于充分应用植物具备的生态功能。
我国有车前科植物1属约18种,新疆产11种,分布于全疆各地,也是天山天池景区常见植物。车前属植物多为理想的药用植物,或是优良的防风固沙植物。对车前属植物的研究集中在药用植物资源调查方法[14],关于车前属植物种子萌发的研究较少[6,15]。因此,本研究以分布在天山天池景区的3种车前属植物包括小车前(plantagominuta)、车前(Plantagoasiatica)和大车前(Plantagomajor)为材料,通过温度对其种子萌发影响的研究,旨在探索车前属种子萌发对策和生态适应性,为充实车前属植物对高山草原和荒漠环境的适应机制及对景区植被保持和恢复提供科学依据。
1 材料与方法
1.1 供试种子
实验所用的小车前种子于2017年6月采自天山天池景区的北部沙漠景区,车前和大车前种子于2017年8月采自天山天池景区天池核心区和灯杆山景区,于种子成熟尚未脱落或少量脱落时进行采集。将采集的种子自然风干后,清理干净,放置于实验室(平均温度为23 ℃)环境中保存备用。实验前对3种植物供试种子采用TTC法进行活力快速测定,经检测种子活力均为96%以上。
1.2 种子萌发
先将种子进行分拣,尽量去除空粒种子。设置3个变温(15 ℃/25 ℃、20 ℃/30 ℃、25 ℃/35 ℃)和3个恒温(20 ℃、25 ℃、30 ℃)条件,所有萌发在光照12 h/黑暗12 h的培养条件下进行。3种车前属植物每个温度条件设置5个重复,每个重复随机选取50~100粒种子(小车前种子为50粒),置于垫有2层滤纸的90 mm培养皿中。每皿加入5 mL蒸馏水,萌发期间及时补充蒸馏水,以保证种子萌发所需水分,实验持续7 d。每天统计萌发情况,以胚根长为2 mm作为标准判断种子是否萌发。计算公式为:
萌发率(%)=(7 d累计发芽种子数/供试种子数)×100%;
发芽势(%)=(规定4 d内发芽种子数/供试种子数)×100%;
发芽指数=∑(Gt/Dt);
式中:Gt为与Dt相对应的每天发芽种子数;Dt为发芽天数。
萌发时滞(d)为萌发开始时间,即第一粒种子萌发所需要的天数。
1.3 数据统计
用SPSS Statistics 19软件进行数据统计和差异性分析,用Tukey法进行多重比较。采用Graphpad Prims 5.0作图。
2 结果与分析
2.1 温度对3种车前属植物种子发芽起始时间的影响
统计分析结果显示,在不同培养温度条件下,3种车前属植物种子发芽起始时间有所不同(图1,表1),无明显规律。小车前种子在恒温条件下发芽起始时间较相对应的变温条件快,变温(15 ℃/25 ℃、25 ℃/35 ℃)显著延迟萌发起始时间,在恒温20 ℃和25 ℃培养条件下培养1 d即可开始萌发,但变温15 ℃/25 ℃和25 ℃/35 ℃时,培养2.8 d和2.6 d种子才萌发;变温培养条件也显著延迟了车前种子发芽时间,尤其在15 ℃/25 ℃培养条件下,培养4 d才可萌发;大车前种子发芽起始时间与小车前和车前略有不同,除变温15 ℃/25 ℃和25 ℃/35 ℃培养条件外,恒温20 ℃也显著延迟萌发起始时间,均达到30 ℃培养条件下发芽起始时间的3倍。
同一培养温度条件对3种车前属植物发芽起始时间的影响效应也有所不同,但总体情况为小车前种子萌发起动较快(除25 ℃/35 ℃),车前和大车前则明显延迟,且存在显著差异。
2.2 温度对3种车前属植物种子萌发进程的影响
6种培养温度下小车前、车前和大车前的种子萌发进程变化趋势相近,即均呈现随培养时间延长,种子萌发率逐渐增加。其中,大车前在20 ℃/30 ℃和25 ℃的培养温度下种子萌发呈快速且集中的特点,在20 ℃/30 ℃培养条件下所有种子在第4天即全部萌发,在25 ℃则提前至第3天;车前在20 ℃/30 ℃培养条件萌发更为集中(萌发率为100%),在25 ℃时萌发趋势与大车前相近,但萌发率约为85%。
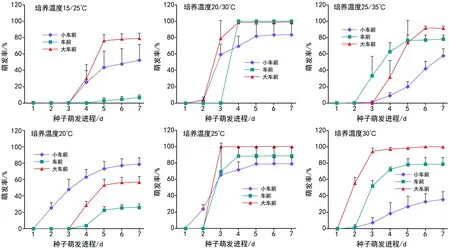
图1 3种车前属植物种子萌发进程受培养温度的影响效应
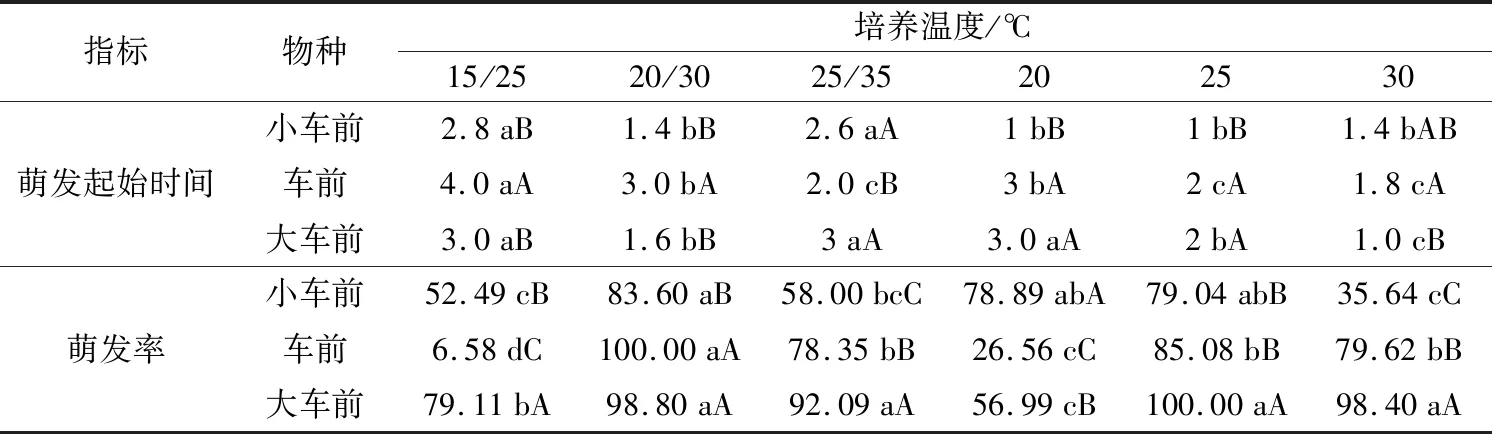
表1 3种车前属植物种子萌发率受培养温度的影响效应
注:不同小写字母表示同一植物种的萌发率在培养温度间差异显著;不同大写字母表示不同植物种在同一培养温度间差异显著。
2.3 温度对3种车前属植物种子萌发率的影响
由表1可知,3种车前属植物种子萌发率表现出相同规律,即种子萌发最适宜的变温环境为20 ℃/30 ℃,恒温培养条件为25 ℃;同时无论在变温培养条件下还是在恒温培养条件下,3种车前属植物种子萌发率均表现出随培养温度递增萌发率先增后降的变化趋势。
通过表1还可知,3种车前属植物种子萌发率受培养温度的影响效应也有所不同。小车前和车前种子在变温20 ℃/30 ℃培养条件下萌发率均为最高,且显著高于其他培养温度;大车前种子在变温20 ℃/30 ℃和25 ℃/35 ℃、恒温25 ℃和30 ℃培养条件下萌发率均超过90%,且以25 ℃时萌发率最高为100%,但4种培养温度间萌发率无显著差异。较低的变温(15 ℃/25 ℃)和较高的恒温(30 ℃)显著抑制小车前种子萌发,车前种子不适宜在变温15 ℃/25 ℃和恒温20 ℃条件下萌发,恒温20 ℃也不利于大车前种子萌发。
在同一培养温度条件下,3种车前属植物间萌发率对培养条件的响应有所不同,其中,大车前在变温(20 ℃/30 ℃培养条件下其萌发率与车前无显著差异)和部分恒温培养条件下的萌发率要显著高于小车前和车前,表明大车前种子萌发所需的温度条件比较宽泛。
2.4 温度对3种车前属植物种子萌发特征的影响
种子发芽势、发芽指数是评价一定条件下种子萌发优劣的常用指标之一。由图2可知,3种车前属植物种子发芽势、发芽指数表现出与萌发率相一致的变化规律,即变温环境20 ℃/30 ℃、恒温25 ℃时,种子的发芽势和发芽指数均为最高;并且在3个变温和3个恒温培养条件中,3种车前属植物种子发芽势和发芽指数均表现出随培养温度递增呈先增后降的趋势。
由图2可知,3种车前属植物种子发芽势和发芽指数受培养温度的影响效应也有所不同。变温(20 ℃/30 ℃)、恒温(20 ℃和25 ℃)培养条件使小车前种子的发芽势和发芽指数显著增加,车前种子也表现出相同的萌发特征;变温(20 ℃/30 ℃)培养条件下大车前种子的发芽势和发芽指数显著高于其他5个培养温度,这与其萌发率受培养温度影响的变化趋势不一致。变温(15 ℃/25 ℃和25 ℃/35 ℃)和恒温(30 ℃)显著降低了小车前种子发芽势和发芽指数;对于车前种子而言,在培养温度依次为20 ℃/30 ℃、25 ℃、30 ℃和25 ℃/35 ℃、20 ℃和15 ℃/25 ℃条件下,其发芽势呈显著下降趋势,但其发芽指数则无相似表现;20 ℃/30 ℃、25 ℃和30 ℃培养条件下大车前种子发芽势无显著差异,但要显著高于其他培养温度,其发芽指数则在25 ℃培养条件下为最高,与其他培养温度间的发芽指数存在显著差异。
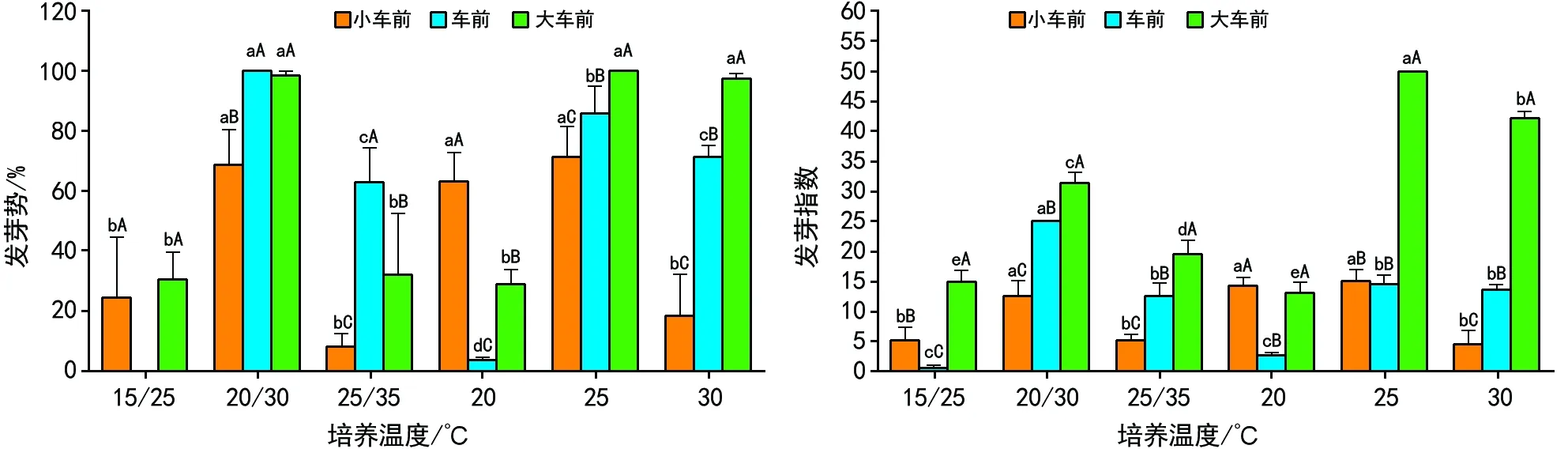
注:不同小写字母表示同一植物种的发芽势和发芽指数在培养温度间差异显著;不同大写字母表示不同植物种在同一培养温度间差异显著。 图2 3种车前属植物种子发芽势和发芽指数受培养温度的影响效应
3 讨 论
根据种子的发芽起始时间、萌发进程等可将植物种子萌发类型划分为爆发型、过渡型、缓萌型和低萌型[16-17],大量研究表明,爆发型的萌发过程是一种带有很大风险的萌发对策[8,10]。在本研究中大车前种子在20 ℃/30 ℃、25 ℃/35 ℃、25 ℃和30 ℃ 4个培养温度下均表现为爆发型,萌发率均超过90%;车前种子在20 ℃/30 ℃时大量且集中的萌发,这种带有很大风险的爆发型萌发其实也是植物对进化过程中的一种适应策略。在自然生长条件下,车前和大车前在长达10~20 cm穗状花序上会产生上千个小种子(每个车前果实中有种子5~6枚,大车前则有6~30枚)且种子表面光滑无任何附属结构,大量成熟后的种子便被密集贮藏在母株周围,形成有限空间内种子密度非常大的土壤种子库。这也意味着从种子萌发到幼苗建植过程中时刻存在空间、时间、资源上的剧烈竞争。这种爆发型萌发方式有可能是通过在一定程度上减少土壤种子库种子数量以保证未萌发种子有效萌发;也能通过种内竞争淘汰弱小幼苗,保证强壮个体良好生长,从而有利于种群的建立和延续,成为增大物种适合度的一种生态策略。
温度是影响植物种子萌发的最主要原因之一[18],综合其在种子萌发中发挥的作用,可概括为直接影响萌发和调控休眠[19,20]。本研究中的实验材料车前和大车前种子均具休眠特性[14],预实验表明在10 ℃/20 ℃变温环境下,车前和大车前种子萌发率为0;当温度升高5 ℃即15 ℃/25 ℃后,其种子萌发率分别为6.58%和79.11%;当温度继续升高5 ℃(20 ℃/30 ℃)后,其萌发率分别达到最高值100.00%和98.80%,表现出不同温度直接影响种子萌发效应的显著差异。小车前种子在变温培养条件(10 ℃/20 ℃、15 ℃/25 ℃、20 ℃/30 ℃、25 ℃/35 ℃)和恒温条件(20 ℃、25 ℃)下,其种子萌发率均超过50%;但当温度升高至30 ℃时其种子萌发率呈显著下降趋势,仅为35.64%,对未萌发种子进行活性检测显示超过90%的种子胚为红色,表明其依旧为有活力种子。研究表明,当温度达到35 ℃时,纳罗克非洲狗尾草种子会进入次生休眠[21],新鲜小车前种子具休眠特性[22],本研究的结果显示,随培养温度梯度增加小车前种子萌发经历了从萌发-再休眠的过程,这种萌发特征反映出温度调控休眠的特性。另外,大量研究表明变温处理较恒温环境更有利于大多数种子萌发[6,23],还可促进未成熟或浅休眠种子萌发[8,24]。本研究中,20 ℃/30 ℃变温较其他培养温度条件更有利于车前属3种植物种子萌发,原因可能是变温激发了酶的活性,有利于贮藏物种的转化和胚的生长;同时变温还可加速种皮的机械变化,使水分和空气易于进入而促进种子萌发。这种效应通常被认为是具有较强休眠特性的野生物种的种子对自然环境的一种适应性,具有重要的生态学意义[25,26]。
植物种子萌发率的不同除了受自身种子特性影响外,物种的地理分布和生态分布也影响着种子萌发对温度的响应[27]。产于高海拔的紫花针茅种子在变温10 ℃/20 ℃时有较高萌发率(为74.0%),而产于低海拔的紫花针茅在15 ℃温度下发芽率较高[6]。内蒙古吉兰泰地区的梭梭种子最适萌发温度为25 ℃[28],而吐鲁番沙漠植物园的梭梭种子最适萌发温度为10 ℃[9]。在本研究的3种车前属植物中,大车前分布范围遍及平原绿洲,在水分充足条件下,温度对其种子萌发的限制不显著,在实验设置的6个温度条件下其萌发率最低为56.99%(20 ℃培养条件),最高为100%(25 ℃培养条件);车前则分布的海拔为1 000~2 000 m,种子在20 ℃/30 ℃、25 ℃/35 ℃的变温环境和25 ℃、30 ℃的恒温环境下萌发率为高,这2种生长在平原绿洲和山地草原的植物种,适宜其种子萌发的温度范围随着海拔升高相对缩小,萌发温度上的差异使得物种的生态位在空间上的分化变大,降低种间竞争,有利于提高物种多样性。小车前种子采自北部沙漠景区,其种子萌发率在高温环境下(30 ℃)显著下降,这与荒漠一年生植物碟果虫实种子萌发对温度的响应相一致[10],这也是小车前种子对荒漠地区不可预测的环境的一种重要的萌发策略,通过种子延迟萌发或保持休眠状态的缓萌策略以减少种子萌发和幼苗生长过程中遇到的死亡风险,确保物种的延续[23]。
猜你喜欢
快乐语文(2020年36期)2021-01-14 01:10:32
中国盐业(2018年16期)2018-12-23 02:08:28
中国特种设备安全(2018年10期)2018-12-18 02:17:18
小学生学习指导(低年级)(2018年9期)2018-09-26 05:59:28
读者·校园版(2018年1期)2017-12-16 18:19:11
启蒙(3-7岁)(2017年9期)2017-02-26 02:52:48
中国粮油学报(2016年5期)2016-01-23 02:44:44
中国惯性技术学报(2015年1期)2015-12-19 13:12:12
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15 12:47:44
中国光学(2015年5期)2015-12-09 09:00:43
