
基于最优解析的陀螺现场免转台标校方法
2020-04-16 01:39杨管金子李建辰黄海国琳娜
兵工学报 2020年3期
杨管金子,李建辰,黄海,国琳娜
(1.中国船舶集团有限公司第705研究所,陕西 西安 710075;2.水下信息与控制重点实验室,陕西 西安 710075)
0 引言
作为运动载体姿态测量的重要惯性器件,陀螺在使用过程中由于运输、振动及长时间存放等因素,在不同误差项上会积累一定的漂移误差,该误差的积累会直接导致载体姿态测量精度难以满足原定的指标需求,因此在一定时间后需对陀螺进行再次标校,以保证陀螺经过长时间运输及存放过程后的姿态测量精度要求。解决这类问题的传统方法是将陀螺返厂,在实验室条件下采用高精度转台设备,通过速率及位置法对陀螺刻度因数及漂移进行标校,这无形中增加了用户对陀螺的使用成本。而一般的现场标校方法对位置有着一定限制,且需要辅助设备或工装的支持,增加了用户的现场操作难度及限制[1-2]。
三轴陀螺的现场标定由于缺乏高精度设备支持,无法获取准确的输入基准,因此拟根据陀螺的静态及动态输出特性,参考以下原理进行陀螺刻度因数及漂移的现场标校:1)静态条件下,陀螺三轴输出模值应与地球自转角速率模值相等;2)动态旋转后,动作前的三轴加速度计输出,经过陀螺动态解析出的姿态阵转换后的计算值,应与动作后的三轴加速度计输出相等。根据上述两个相等原理可以构造关系等式。由于器件及现场环境的干扰,等式无法进行有效求解,而将陀螺的现场标校问题转化为最优解析问题进行最优解析,即可得到需标校误差参数值[3-5]。
陀螺标校过程中,由于动态旋转采样的关系,刻度因数及漂移对陀螺输出精度的影响存在耦合关系,需要分别对刻度因数及常值漂移进行动态及静态数据采集并进行标校;另外,陀螺动态实时姿态解析,加上优化解析过程中过于宽泛的初始范围,极易导致求解过程中的盲目进化及过早陷入局部最优[6-7]。
为满足陀螺长期的现场工作精度需求,贴合及方便用户的使用,本文拟对中等精度陀螺的现场无设备及工装支持的标校方法进行研究。研究过程中针对陀螺动态解析带来的耦合及局部最优问题,将进一步针对刻度因数与漂移的隔离标校方法、改进遗传算法最优解析、基于陀螺输出特性的初始搜索范围预先逼近方法等进行优化改进。
1 改进的遗传算法最优解析
根据三轴陀螺的输出特性分析,由标校问题转化而成的最优求解问题存在多参量及强非线性特性,通过对不同数学方法的比对,本文选取遗传算法作为该最优解析的数学方法。遗传算法通过模拟生物的进化及交叉变异过程实现对最优数值的递推逼近。作为一种通用性好、鲁棒性强的启发式随机搜索优化算法,遗传算法在保证局部搜索精度的同时兼顾全局搜索能力,并能较好地根据对象的不同使用特性来完成对不同功能项的适应性改进。
图1所示为遗传算法的一般流程,大致分为初始化阶段、适应度计算阶段、判定阶段、新种群形成阶段(即交叉变异阶段)。具体步骤如下:1)确定待求解参数的大致范围,形成初始搜索范围,在该范围中建立初始种群;2)构建一种最优函数形式,对种群中的个体进行适应值计算描述,以对个体的优劣进行判断;3)根据适应值找出该代种群中的最优个体,将该最优个体与事先设计的收敛判据进行比对,若符合则可宣布获得待求解的参数;4)若不符合判据,则对余下的个体进行交叉变异操作,交叉变异后的个体与该最优个体共同形成新一代的种群继续迭代步骤2~步骤4,直至某一代最优个体符合收敛判据。
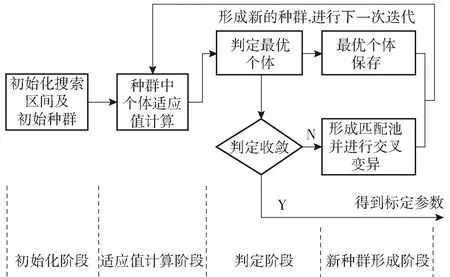
图1 遗传算法一般流程图Fig.1 General flow chart of genetic algorithm
其中,交叉变异为遗传算法中的关键环节之一,它关系着遗传算法的进化方向及样本多样性,并影响着全局及局部搜索精度。以下主要对交叉变异阶段进行介绍。
针对陀螺标校计算过程中需要实时进行姿态转换的问题,为保证陀螺实时更新的数据精度,本文对遗传算法采用实数编码模式,以减少转码带来的计算及精度损失。在此前提条件下,交叉变异过程采用随机加权的形式进行替代;同时,考虑到陀螺输出自身存在着随机噪声,相比于传统遗传算法,对其交叉变异过程应存在一定的束缚限制,以免生成超过范围的异常个体,导致迭代过程中出现收敛较慢甚至发散的现象。
交叉过程公式如下:
(1)
式中:c(t,i)、c(t,j)为交叉过程选中的父代种群中的第i、j个待交叉个体;t为迭代次数;c(t+1,i)、c(t+1,j)为生成的子代种群中用于替代父代种群的第i、j个新个体;α1为期望0.5的正态分布。
(1)式保证了生成的子代种群在理论上覆盖了父代种群的所有特征区域,且大概率继承父代优势特征,同时也可能跳出父代特征限制,生成新的种群个体。该设计在保证种群多样性的同时提高了种群的鲁棒性,保持了整体的进化方向。
同样,变异过程公式如下:
(2)
式中:hi为变异过程选中的父代种群中第i个待变异个体;h′i为生成的子代种群中用于替代父代种群的第i个新个体;hmin、hmax分别为父代种群中适应度最小、最大的个体;ra为0和1之间的概率随机数;d(t,x)为[0,x]范围内符合非均匀分布的随机数,x表示任意实数,本文中主要指代(2)式中的hmax-hi和hi-hmin,
d(t,x)=x(1-r(1-t/T)b),
(3)
r为[0,1]区间的随机数,T为预设的最大迭代次数,b为系数参数。
(3)式使得变异过程的影响随着迭代次数的增长以某种速度减小,将迭代过程以该速度进行收敛,使得进化方向保证随着迭代次数的增长而逐步统一,减少盲目的进化方向及不必要的计算资源浪费。
交叉、变异并不是针对所有的父代个体,其目的是为了防止遗传算法陷入局部最优陷阱,同时加速收敛过程,因此需要对交叉和变异的个体进行有目的的选择。本文采取一种自适应的交叉概率和变异概率设定方式,概率公式如下:
(4)
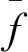
(4)式使得对遗传算法进化方向有用的个体更容易进行交叉和变异,从而激发出更加逼近真值的下一代个体。相对于固定的概率值,上述自适应概率公式的设计能够进一步加快遗传算法的收敛速度。
2 静态条件下的常值漂移现场标校方法
在动态条件下,陀螺的刻度因数和漂移对陀螺输出精度的影响存在耦合关系,无法对其进行辨识,因此,需要对陀螺的刻度因数和漂移进行隔离标校。根据第1节所述原理,静态条件下,陀螺三轴输出模值应与地球自转角速率模值相等,且在静态条件下刻度因数对陀螺输出的影响不会彰显。因此,可在静态条件下对陀螺的漂移进行预先标校,来消除在动态条件下漂移对刻度因数标校的耦合影响问题。根据上述原理,构建如下等式:
(5)
(5)式显示出即使在静态条件下,陀螺输出中仍包含着地球自转的影响,因此该项地球自转角速率不能看作常值,必须将其进行刻度因数的加权。根据刻度因数的初始范围,将该项地球自转角速度作为变量,与陀螺三轴漂移共同构成4变量参数的最优化问题:
(6)
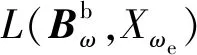
Xωe=[ak,bk]×ωe,
(7)
ak、bk为预估的刻度因数初始范围;按照静态条件,陀螺输出的最大值及最小值作为漂移的初始搜索范围的上下界即可。
通过(6)式即可完成对漂移项的最优解析,从而预先标校出漂移,消除动态条件下漂移对刻度因数的耦合影响。
下面采用数学仿真的形式对上述漂移标校问题进行验证。数学仿真模拟参数指标如表1所示。
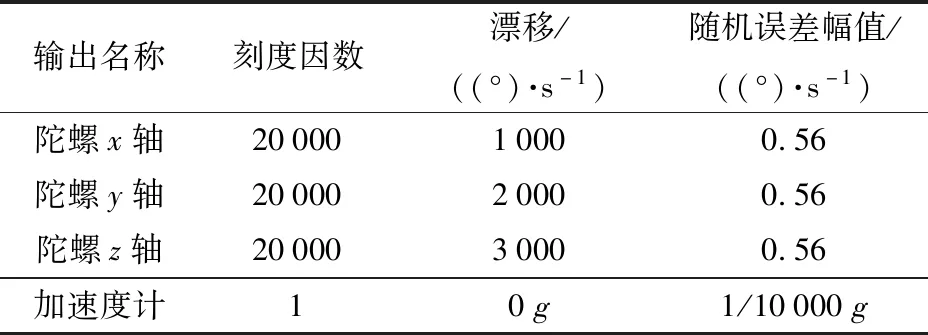
表1 仿真参数Tab.1 Simulation parameters
对陀螺分别进行不同姿态下的静态放置,陀螺及加速度计的连续采样原始数据如图2所示。
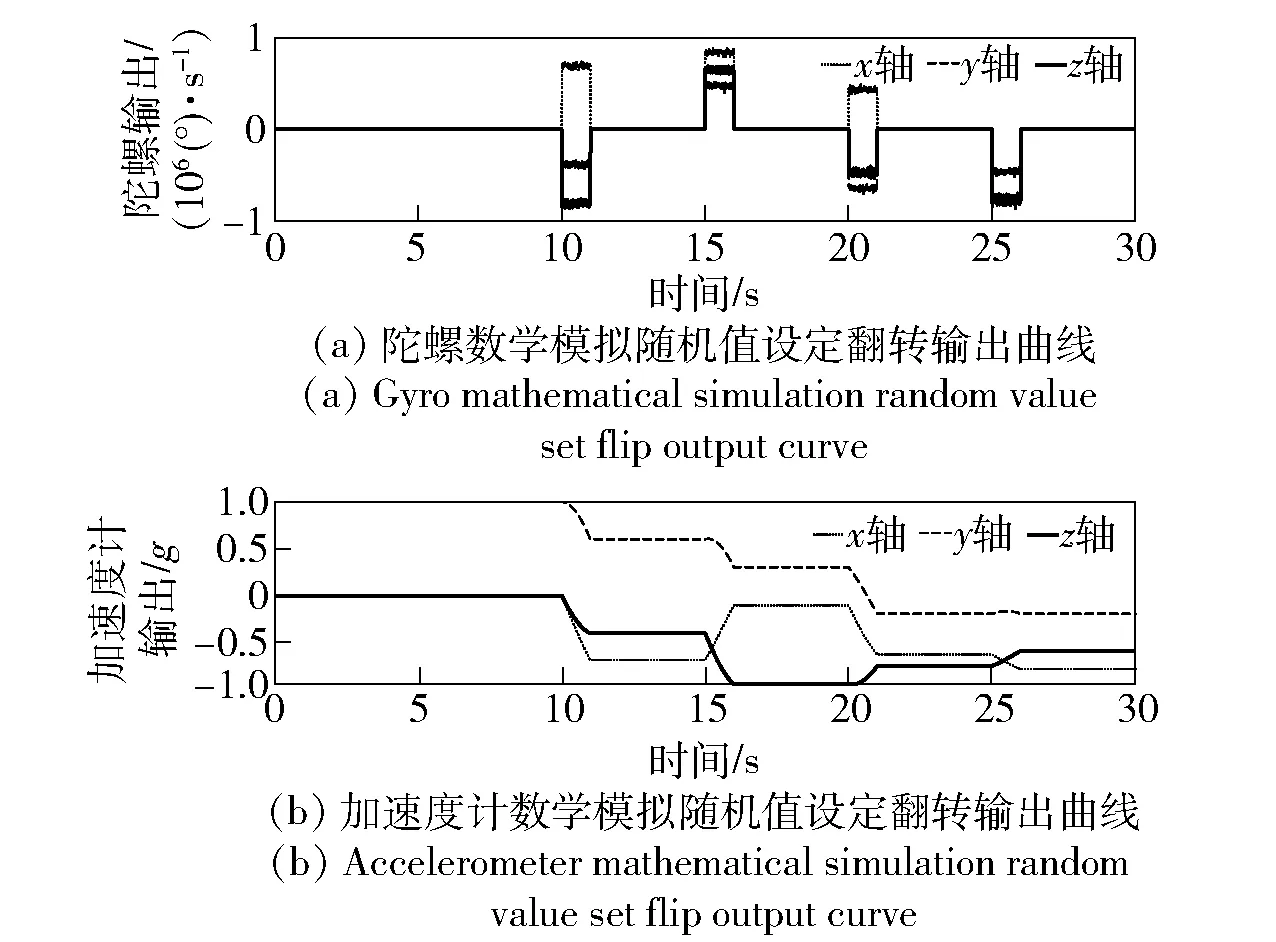
图2 数学模拟条件下的惯性测量组合输出曲线Fig.2 Output curves of inertial measurement unit in mathematical simulation
对上述数据,截取陀螺静态数据进行优化函数构建,采用遗传算法进行最优解析。遗传算法迭代目标函数价值变化曲线及三轴漂移参数解析迭代过程如图3所示。
由图3可见,漂移间参数相差较大,无法准确观察到漂移的收敛情况,最终具体标校参数如表2所示。

图3 遗传迭代过程中的参数收敛曲线Fig.3 Convergence curves of parameters in process of genetic iteration

表2 现场标校结果Tab.2 Field calibration results
由表2可见,通过设定的刻度因数加权补偿后,标校得到的漂移参数与设定值间的误差为[0.003 °/h,0.053 °/h,0.060 °/h],该标校误差与设定不可补偿常值漂移误差0.1 °/h相符。该精度验证了本文所提出方法的有效性,即在数学仿真基础上能够达到理论的标校精度。
3 动态条件下的刻度因数现场标校方法
完成陀螺的静态漂移标校后,即可根据引言中的原理2,以加速度计输出作为姿态转换的方向基准,构造以陀螺刻度因素为解析变量的目标函数,对陀螺刻度因数值进行标校。
将上述原理过程进行以下公式的推导:
(8)
(9)
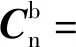
(10)
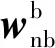
通过(8)式~(10)式,可根据陀螺的采样数据对姿态阵进行实时更新。
测量出初始时刻的三轴加速度计输出,通过(11)式对每一静止时刻的加速度计输出值进行计算:
(11)
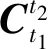
通过(12)式对加速度计实际输出和计算输出进行差值模计算:
(12)

通过(13)式、(14)式构造陀螺标校的目标函数:
(13)
(14)
式中:kax、kay、kaz分别为陀螺三轴的刻度因数;Nwx、Nwy、Nwz分别为陀螺三轴的原始输出;bwx、bwy、bwz分别为陀螺三轴的漂移。
目标函数构成后即可采用遗传算法,在搜索范围内对其进行参数最优解析,得到最终标校结果[8-11]。
在开始进行遗传算法迭代计算前,最优解析问题初始搜索范围的有效设定,一方面可以降低最优化过程过早陷入局部最优的风险,另一方面可以有效地抑制盲目的进化方向,减少无效计算过程;同时,本文所涉及的刻度因数及漂移标校过程皆建立在刻度因数初始范围基础上,因此需结合陀螺标校自身特性,对刻度因数初始范围进行有效地逼近缩小。
以往对惯性器件的使用经验及厂家提供的大量数据显示,同厂生产的同类型惯性器件,装配到同一产品后其刻度因数值大致相同。基于此,在遗传迭代前,可以采用对生成种群进行分割、并行计算、种群淘汰的方法,同一速率、有指向性地将陀螺三轴刻度因数的搜索范围逼近至一个有效的、较小的靠近真值范围内。该方法的大致流程如下:
1)预先按照所需标校的陀螺三轴正方向,旋转陀螺,判定刻度因数符号;
2)依据三轴模值粗略计算辅以加权,得到一个较大的初始搜索范围,生成初始种群;
3)对初始范围进行分段区间划分,将初始种群分割成H个子种群(H可取5~10),对每个子种群进行M次随机复制,并计算每个种群的价值(即目标函数值);
4)对比各价值均值,认为较小价值均值对应的子种群拥有进化优势,淘汰掉一部分具有明显劣势的子种群;
5)对生存的子种群再次进行H段分割,进行新一轮的生存、淘汰挑选;
6)直至第j子种群,其价值计算结果仅在小范围波动时,则认为无法继续进行分割淘汰,认定形成遗传算法的最小初始搜索范围[12-15]。
按照上述方法,在数学仿真过程中将初始搜索范围从0~0.000 264,缩小至0.000 048 56~0.000 050 68,区间长度缩短近125倍。相关过程如图4所示。图4中X表示无法再继续分割的初始范围集。

图4 并行计算种群淘汰过程Fig.4 Parallel computing of population selection
同理,在第2节的数学仿真条件下,对陀螺刻度因数标校问题进行验证。摘取陀螺动态数据进行姿态转换计算过程,加速度计静态数据进行转换前后对比基准,构建最优函数进行最优解析,遗传迭代目标函数变化及三轴刻度因数变化曲线如图5所示。具体标校参数如表3所示。
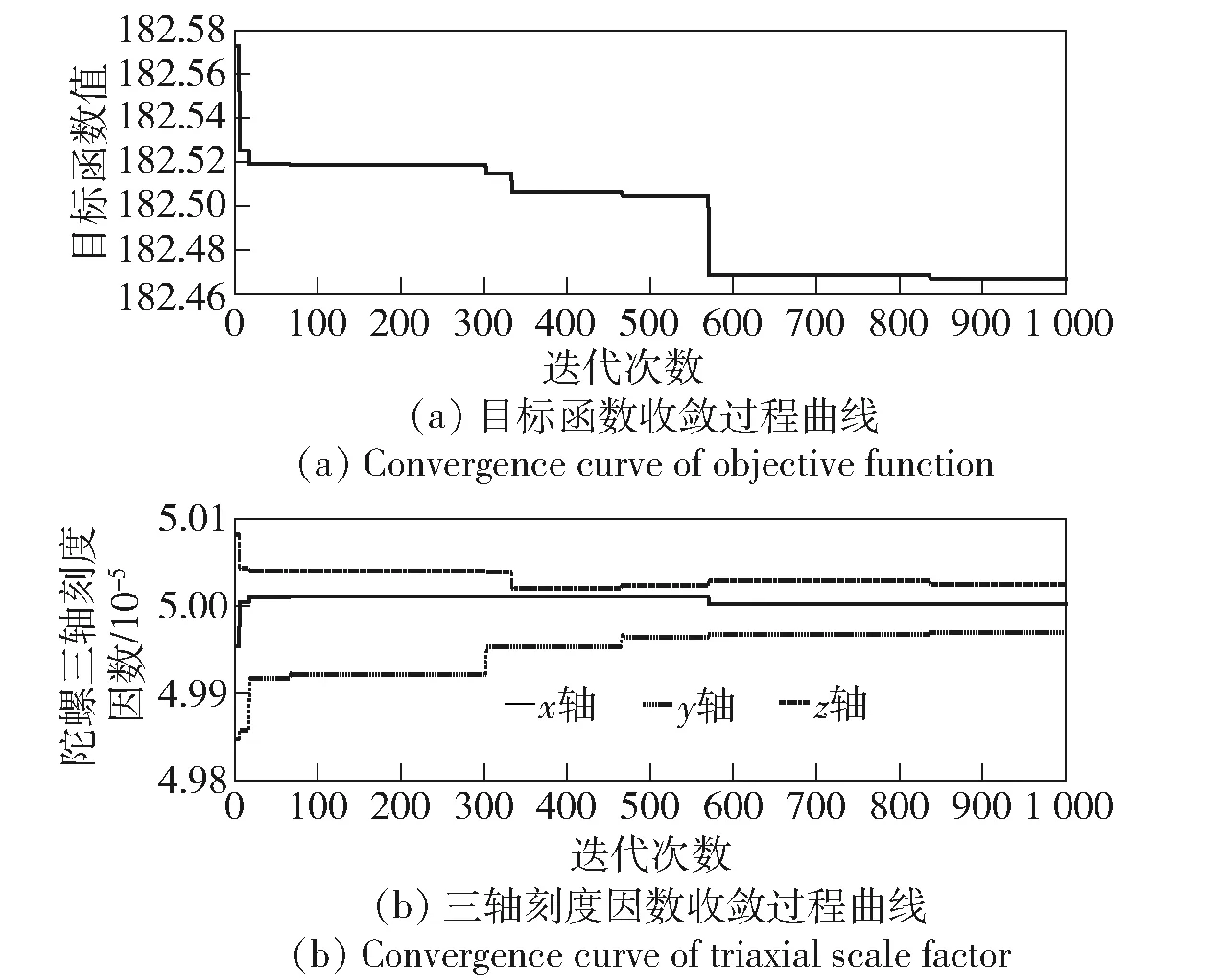
图5 遗传迭代过程中目标函数值及陀螺刻度因数收敛曲线Fig.5 Convergence curves of objective function value and gyro scale factor in genetic iteration
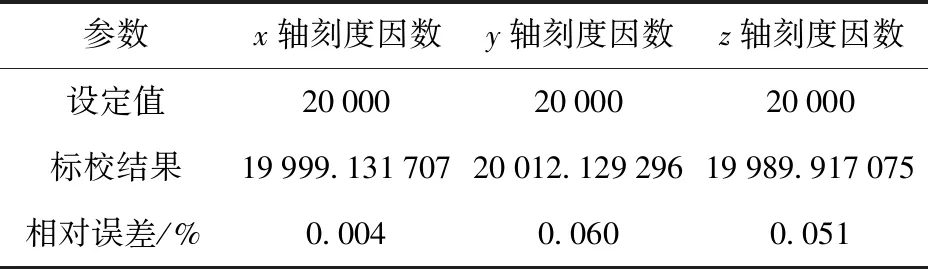
表3 现场标校结果Tab.3 Field calibration results
由表3可见,最终标校刻度因数与设定刻度因数间相对误差为[0.004%,0.060%,0.051%],验证了本文方法在刻度因数误差项上的现场标校精度。
4 方法综述及仿真验证结果
根据现场所具备的条件,本文所提新的陀螺现场标校方法大致流程如图6所示。图6中,b0表示标校得到的陀螺漂移参数。
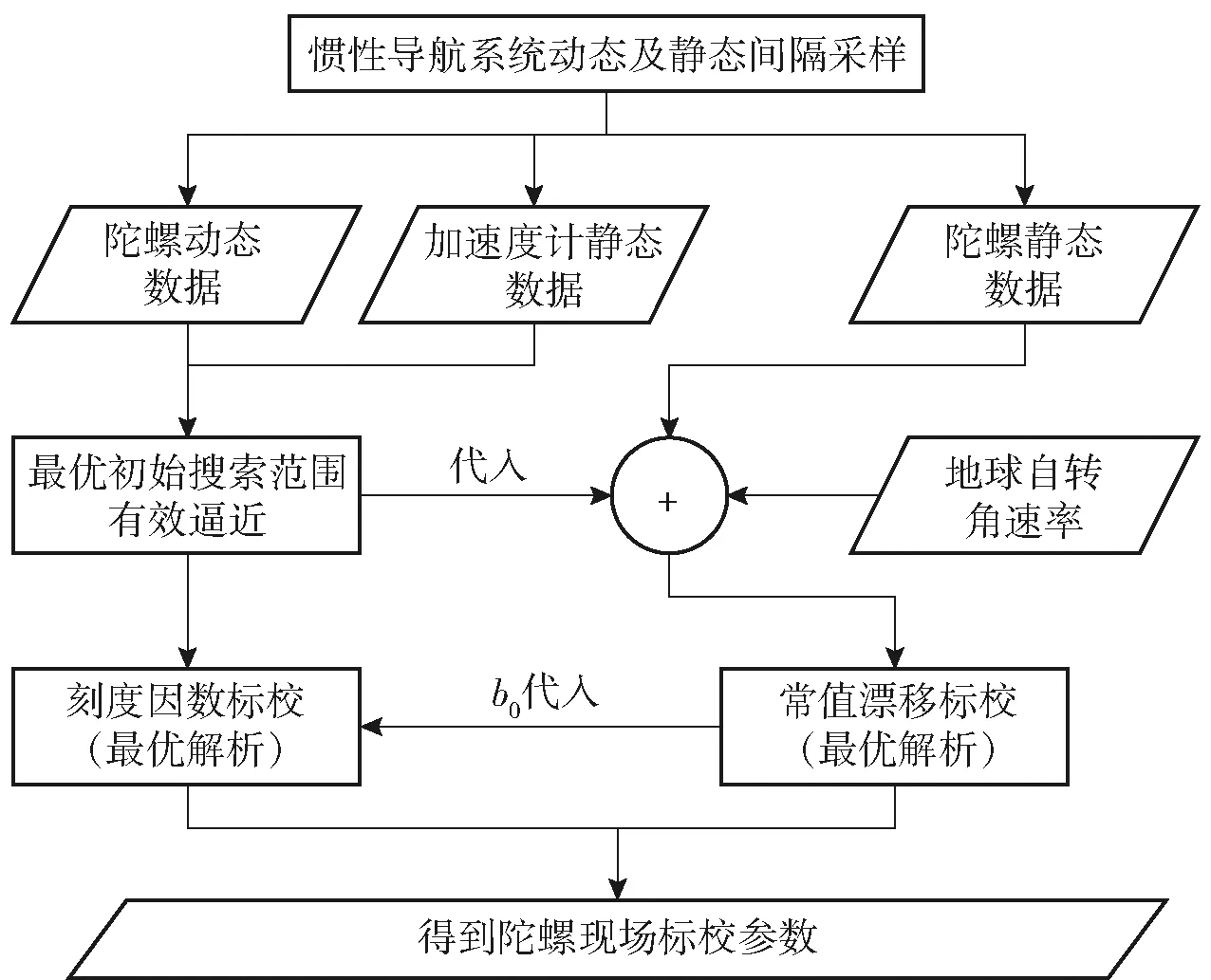
图6 三轴陀螺标校流程图Fig.6 Flowchart of three-axis gyroscope calibration
具体标校流程及操作方法如下:
1)以现场桌面或地面作为静态平面基准(无需工装),对陀螺3个敏感轴分别进行动态旋转及静态放置连续采样(徒手翻转即可,为保证陀螺输出精度,在静态放置及翻转过程中尽量保证时间在10 s以上),获得时序上连续的陀螺动态、静态数据和加速度计静态数据;
2)陀螺动态数据及加速度计静态数据用于进行刻度因数标校,预先采用初始分段并行种群淘汰机理,对刻度因数初始搜索范围进行方向性逼近;
3)地球自转角速率加权初始搜索范围,结合陀螺静态数据,参考引言中原理1构建漂移标校最优函数,漂移初始范围边界参考陀螺静态数据最大值及最小值加减地球自转角速率,进行漂移的最优解析;
4)原始数据经常值漂移补偿后,根据引言中原理2建立刻度因数标校最优函数,刻度因数初始搜索范围参考步骤2的逼近结果,进行刻度因数的最优解析;
5)结合步骤3、步骤4的最优解析结果,得到最终需标校参数。
下面采用光纤惯性测量组合实体样机对陀螺现场标校方法进行半实物仿真实验验证,该光纤惯性测量组合实体样机陀螺不可补偿漂移在0.1 °/h左右。对实体样机在工作桌面上徒手随意进行的翻转情况及采集的原始输出曲线如图7所示。
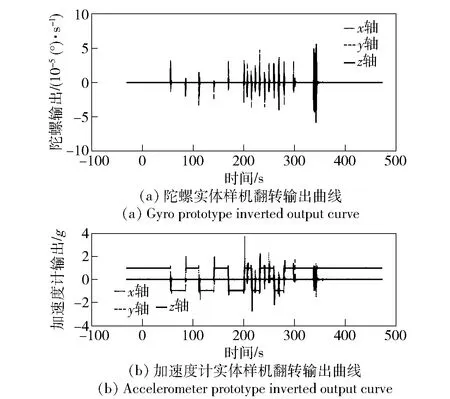
图7 惯性测量组合实体样机现场翻转过程中的输出曲线Fig.7 Output curves of inertial measurement unit prototype in the field turnover process
由图7可见,动态翻转过程中,角速率及姿态随机,且时间较短,整个过程约8 min30 s,截取其中209~258 s,即49 s的时间进行标校计算。陀螺标校过程中遗传迭代目标函数变化过程曲线及三轴刻度因数变化过程曲线如图8所示。标校最终结果如表4所示。
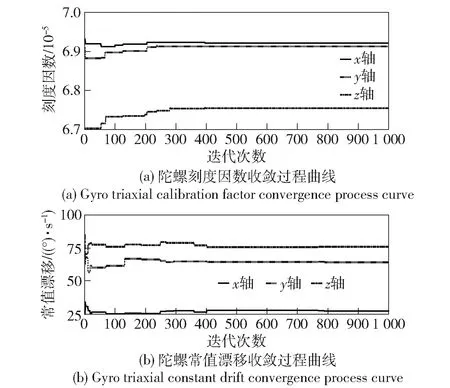
图8 遗传迭代过程中陀螺误差参数的收敛曲线Fig.8 Convergence curves of gyro error parameters in genetic iteration

表4 现场标校结果Tab.4 Field calibration results
由表4可见,与转台标定值相比,陀螺常值漂移现场标校误差为[4.74 °/s,6.92 °/s,6.44 °/s]×10-5,即[0.17 °/h,0.25 °/h,0.23 °/h],基本符合该实体样机的不可补偿漂移指标,刻度因数现场标校相对误差为[0.14%,0.04%,0.25%]。上述标校结果即来自现场徒手操作的标校过程,与该陀螺的转台标定结果相比,该验证实验结果表明,本文所设计方法的标校精度能够完全满足一般运动载体的姿态测量指标需求,且其所利用的数据长度不超过1 min,与传统标定时间(需要对各轴进行多次且保证一定时间的设定旋转)及对器件误差的估计时间(一般需要至少两个位置各100 s的估计时间)相比,本文所设计方法能够缩短标校时间。
通过上述标校值对该实体样机输出进行参数误差补偿,再通过补偿后姿态角解算值与转台标定补偿后输出值进行比较,以判定该标校方法的可行性及有效性。验证实验过程中陀螺及加速度计输出曲线如图9所示,通过本文设计方法补偿后姿态角解算值与转台标定补偿后姿态角解算值的对比误差曲线如图10所示。

图9 转台模拟惯性测量组合机动过程中的输出曲线Fig.9 Turntable simulated output curves of inevtial measurement unit during maneuvering
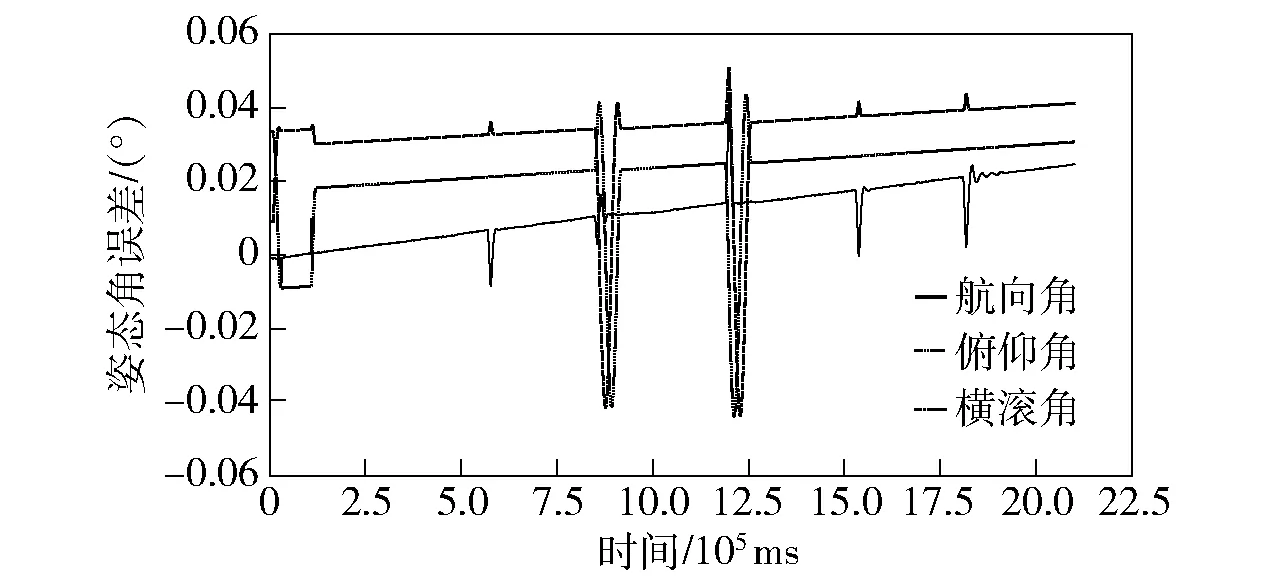
图10 转台及现场标校补偿后姿态角输出比较误差Fig.10 Errors of attitude angle outputs after turntable and field calibration compensations
由图10可见,在一定的时间内对姿态角进行解算分析,通过与转台标定实体样机姿态角输出进行比较后发现,现场标校补偿后,无论旋转及静态条件下对比误差不超过0.1°,对比结果充分证明了本文所提陀螺现场标校方法的可行性与有效性。
5 结论
本文针对陀螺长期使用误差积累的问题,同时为贴合用户实际现场使用需求,提出一种基于最优解析的陀螺现场标校方法。该方法利用陀螺使用过程中的动态及静态数据,对漂移及刻度因数分别进行标校计算,消除了漂移及刻度因数间的耦合影响问题;通过对刻度因数初始范围的逼近,解决了优化过程中的盲目进化及过早陷入局部最优问题。该方法在保证陀螺整体现场标校精度的同时,简易了操作过程,缩短了标校时间,摆脱了对实验条件的束缚,提高了标校现场操作的便利性,具有较强的实际应用价值。
数学仿真及半实物仿真实验结果显示,该方法在现场标校过程中的刻度因数标校精度能够达到0.3%,漂移标校精度能够达到0.25 °/h,外特性试验显示姿态角测量精度在动态及静态条件下均能够达到0.1°,完全能够达到目前中等精度陀螺的姿态测量精度需求,从而证明了该方法的有效性,满足了陀螺长期现场使用的精度指标需求。
猜你喜欢
小学生学习指导(中年级)(2021年3期)2021-04-06
学苑创造·A版(2019年9期)2019-11-07
小学生学习指导(低年级)(2019年10期)2019-10-16
学生天地(2019年6期)2019-03-07
小学生学习指导(高年级)(2018年3期)2018-11-29
小学生学习指导(中年级)(2018年9期)2018-11-29
快乐语文(2018年36期)2018-03-12
学苑创造·B版(2017年1期)2017-02-21
学苑创造·B版(2017年1期)2017-02-21
故事作文·高年级(2016年6期)2016-06-21
