
青藏高原冻融-水蚀凹陷对高寒沼泽草甸土壤呼吸的影响
2020-04-15 02:35丁俊霞周贵尧陈克龙陈生云
水土保持通报 2020年1期
丁俊霞, 周贵尧, 陈克龙, 陈生云, 崔 航
(1.青海师范大学 地理科学学院,青海 西宁810008; 2.华东师范大学 生态与环境科学学院 上海 200241; 3.青海师范大学 青藏高原地表过程与生态保育教育部重点实验室,青海 西宁 810008; 4.青海师范大学 科技处 青海 西宁 810008; 3.高原科学与可持续发展研究院,青海 西宁 810008; 5.中国科学院 西北生态环境资源研究所 冰冻圈科学国家重点实验室, 甘肃 兰州 730000)
作为陆地生态系统对气候变化的重要敏感区和生态脆弱区,青藏高原在全球土壤碳循环中发挥着重要的作用[1]。青藏高原土壤有机碳储量达到3.35×1010t 碳,占全国土壤有机碳的23.44%[2]。近年来,温度升高、放牧、氮沉降、冻融及高原鼠兔等生物和非生物因素正深刻影响着青藏高原的生物地理过程,对青藏高原碳循环格局产生了重要影响[1]。作为陆地生态系统和大气圈之间的第二大碳通量,土壤以土壤呼吸的形式每年向大气中排放6.00×1015~9.80 ×1015g的碳,土壤呼吸的微小变化会对大气碳浓度造成重要影响,进而改变碳循环—气候变化间的反馈过程[3]。目前在全球气候变化和人类活动等背景下,青藏高原上开展了大量的不同时空尺度的陆地生态系统土壤呼吸研究,但由于青藏高原自然生态过程的复杂性,青藏高原土壤呼吸过程及其调控机制仍存在广泛争议[4]。尤其是一些外力因素过于独特和复杂,比如在高寒沼泽草甸生态系统中冻融作用与水力侵蚀作用一起塑造形成了较为常见的凹陷微地貌。但目前尚未评估冻融—水蚀作用对高寒沼泽草甸生态系统的影响程度,这些认知局限在很大程度上将严重制约我们未来科学评估和高效预测青藏高原土壤呼吸动态特征。
冻融—水蚀凹陷是青藏高原常见的一种凹陷微地貌,经过漫长的冻融循环和水蚀作用,土壤表层下陷形成的大小不等、形态各异的冻融水蚀凹陷口[5]。冻融交替过程夹杂水力侵蚀作用形成了冻融—水蚀凹陷。在形成的冻融—水蚀凹陷区,土壤含水量、土壤透气性、土壤微生物对养分的活化、地下植物根系及微生物结构和组成发生变化,土壤呼吸及其组分也会发生变化[6-7]。在冻融的最初循环阶段土壤呼吸会增强(秋冬冻融交替期),最后下降至较为稳定的水平(冬季冻结期),随后土壤呼吸再度增强(冬春冻融交替期)[8]。冻融期土壤排放的CO2通量占全年土壤呼吸总额的3%~50%[9]。但当前关于高寒沼泽草甸冻融—水蚀凹陷土壤呼吸过程的研究仍然十分匮乏。自20世纪60年代以来,全球气候变暖,冻融和水蚀作用加剧,已造成青藏高原高寒沼泽草甸大面积退化[10]。在高寒沼泽草甸退化过程中,土壤微环境、有机碳、微生物及植物地下根系等因素均受到一定程度的影响,进而改变了土壤呼吸,导致青藏高原的碳平衡格局发生变化。因此,加强高寒沼泽草甸冻融—水蚀凹陷中的土壤呼吸组分变化和调控机制研究,将有助于精确估算高寒沼泽草甸土壤与大气的碳交换量,完善当前陆地生态系统碳循环过程的理论体系,同时也有利于提高对高寒沼泽草甸长期性冻融—水蚀自然过程与全球气候变化之间的耦合关系的认识。
青海湖湿地位于青藏高原东北部,是维系该地区生态安全的重要屏障,包括湖滨湿地和河源沼泽,总面积有2 800 km2,占流域陆地面积的9.3%,平均海拔为3 800 m[11]。由于特殊的地理位置和气候条件,青海湖湿地对气候变化十分敏感,它是研究土壤碳循环对气候变化响应的理想场所。且由于海拔高、气候差异大,流域内冻融和水蚀作用明显,冻融—水蚀凹陷广泛发育,有利于研究冻融—水蚀凹陷对高寒沼泽草甸生态系统的影响。但目前关于青海湖湿地冻融—水蚀凹陷对高寒沼泽草甸的影响研究较少,基于此,本文以青海湖北部的瓦颜山高寒沼泽草甸冻融—水蚀凹陷微生态系统为研究对象,结合野外观测和室内试验,探究高寒沼泽草甸冻融—水蚀凹陷区2019年5月的土壤呼吸动态特征,分析高寒沼泽草甸冻融—水蚀凹陷中的空气温度、空气相对湿度、5 cm土壤温度、5 cm土壤含水量与植被群落(优势种、地上生物量、植物高度、群落盖度)的变化特征,及以上环境要素的变化与土壤呼吸之间的相互关系,以期为探讨和评估高寒沼泽草甸碳循环过程提供一定的科学依据。
1 研究区概况
研究样地位于青海湖北部海北藏族自治州刚察县的伊克乌兰乡(37°44′34″N,100°5′41″E),北靠祁连山,南临青海湖。气候类型为高寒大陆季风气候,寒冷期长,温暖期短,光照充足,太阳辐射强,干湿季分明,雨热同季。年平均气温为-3.31~1.4 ℃,最高气温为33.3 ℃,最低气温为-36.3 ℃。年平均降水量为426.8 mm,最高降水量为479.4 mm,最低降水量为341.1 mm。年日照时数为2 517.6~2 995.3 h,年辐射量5 210.2~6 568.3 MJ/m2[11]。一般在5月中旬进入生长季,8月为生长季旺期,9月底生长季结束。研究区域内主要为高寒沼泽草甸,小嵩草(Kobresiahumilis)为主要优势种,苔草(Carextristachya)、火绒草(Lobulariamaritima)、鹅绒委陵菜(Potentillaanserina)为主要伴生种。多年冻土最大深度约为1.7 m[12]。
2 材料与方法
(1) 样区选择与设计。瓦颜山研究样区土壤主要为沼泽土和草甸土,土壤厚度约为1.2 m,表层土壤下为洪积堆积物。季节性冻土层约1.7 m,深层为永久性冻土。地表5 cm土壤在11月中旬左右开始冻结,来年4月初开始消融,冻结期达145 d左右[11]。由此可见样区内冻融循环过程时有发生。在此种长期性的冻融循环过程中,土壤表层致密层破裂,在水蚀的作用下土壤表层凹陷、植被和土壤环境系统发生变化。在监测进行前依据发生了较大变化的植被特征(主要依据地上生物量及优势种—小嵩草的高度和群落盖度),在50 m×50 m的高寒沼泽草甸围封样地内,随机选取3处冻融—水蚀凹陷作为研究样区(每个凹陷的深度在9 cm左右,植被主要为小嵩草,凹陷中的小嵩草的高度和群落盖度较非凹陷区低,每个凹陷之间的距离大约在2~5 m)。在选取的3处冻融—水蚀凹陷内外,分别置入3个直径为20 cm,高14 cm的土壤呼吸环(为确保样点之间有较高的相似性,每个重复之间的距离≤0.5 m)。
(2) 土壤呼吸和水热条件测定。为了消除地上植物呼吸的影响,测量的前一天剪掉地面植被,剪掉的植物移除土壤呼吸环。在2019年5月中旬(晴天),利用LI-COR公司的LI-8100A从早上6点到下午18点进行土壤呼吸监测,隔2 h监测一次,共获得7组数据,21个样本量。与此同时,利用LI-8100A的温度和湿度探头同步测定土壤5 cm的温度和土壤含水量,利用古德HTC-1空气温湿度计同步监测近地面空气温度与空气相对湿度。
(3) 植被特征测定。2018年8月17日,采用卷尺在随机选取的3个冻融—水蚀凹陷重复内,绕着凹陷口边缘均匀测量20次,得到冻融—水蚀凹陷程度。在冻融—水蚀凹陷样区和非冻融—水蚀凹陷样区中采用20 m×20 m的样方框采集地上生物,将采集的地上生物装入密封袋。在105 ℃下杀青15 min后,然后在65 ℃下烘干24 h至恒重,称重得到地上生物量。采用1 m×1 m的样方,用卷尺测量优势种—小嵩草的高度(20次测量)。群落盖度为每个种的垂直投影面积占样方总个体数的百分比。
(4) 数据处理与分析。在SPSS中采用配对样本T法分析检验冻融—水蚀凹陷对土壤呼吸、空气温度、空气相对湿度、5 cm土壤温度及5 cm土壤含水量的影响,采用Pearson相关分析法分析土壤呼吸与近地面大气气象因子(空气温度、空气相对湿度)与土壤环境因子(5cm 土壤温度、土壤含水量)的相关性,并通过逐步回归模型分析以上气象要素对土壤呼吸的影响。
3 结果与分析
3.1 冻融-水蚀凹陷中的环境要素特征
3.1.1 冻融—水蚀凹陷中的空气温度与相对湿度的变化 在本次野外监测中,试验地5月份的空气日间(6:00—18:00)气温在-4.7~32.8 ℃范围内,从早晨到中午再到下午表现为先上升再降低的趋势,其中6点左右的温度最低,12点左右的温度达到一天中最大。冻融—水蚀凹陷中的日间空气温度总体比非冻融—水蚀凹陷样区的空气温度低11.66%(p<0.05,n=21,表1),具体表现为在8点后冻融—水蚀凹陷样区的空气温度低于非冻融—水蚀凹陷样区的空气温度,12点左右冻融—水蚀样区和非冻融—水蚀样区的空气温度差异不大,气温在-28~32.8 ℃范围内(图1a)。相对于空气温度,日间(6:00—18:00)的空气相对湿度在25%~76 %范围内,且从早晨到中午再到下午表现为先下降再上升的趋势,其中6点左右的空气相对湿度值达到白天中最大,12点左右的空气相对湿度值达到一天中最小。冻融—水蚀凹陷中的日间空气相对湿度总体比非冻融—水蚀凹陷样区的空气相对湿度高8.69%(p<0.05,n=21)(图1b,表1)。
3.1.2 冻融—水蚀凹陷中的5 cm土壤温度与含水量及植被特征变化 瓦颜山高寒沼泽草甸5月份的5 cm土壤温度在-0.005~4.79 ℃范围内。从早晨6:00—18:00,5 cm土壤温度波动较大,但相较于冻融—水蚀凹陷来说,非冻融—水蚀凹陷区的5 cm土壤温度更稳定,且冻融—水蚀凹陷样区的5 cm土壤温度显著低于非冻融—水蚀凹陷样区(p<0.05,n=21)(图1c,表1)。5 cm的土壤水分含量从早晨—中午—下午表现为先降低再升高的趋势,其中早晨6点左右的土壤含水量是白天中最高,在10:00—14:00时5 cm土壤水分含量是一天中的最低值。而在14:00—18:00时冻融—水蚀凹陷中的5 cm土壤含水量总体高于非冻融—水蚀凹陷区,但二者之间的差别不明显(p>0.05,n=21)(图1d,表1)。
冻融凹陷下的地上生物量和自然条件的地下生物量,分别191.15,483.1 g/m2。相较于非冻融—水蚀凹陷区而言,冻融—水蚀凹陷下的优势种—小嵩草的高度和群落盖度分别降低了68.7%和16.9%。

图1 5月中旬冻融-水蚀凹陷与非冻融-水蚀凹陷样区空气温度,空气相对湿度,5 cm土壤温度,5 cm土壤水分含量的动态特征及差异
表1 冻融-水蚀凹陷区与非冻融-水蚀凹陷区的气候要素及土壤呼吸差异分析

气候要素及土壤呼吸均值标准差均值的标准误差分的 95%置信区间下限上限tdfsig.(双侧)土壤呼吸0.572 380.627 770.136 990.286 630.858 144.178200.000空气温度1.980 953.863 500.843 080.222 313.739 602.350200.029空气相对湿度-3.000 004.335 900.946 17-4.973 68-1.026 32-3.171200.0055 cm土壤温度-1.100 241.414 380.308 64-1.744 06-0.456 42-3.565200.0025 cm土壤含水量-0.719 052.598 390.567 01-1.901 820.463 72-1.268200.219
3.2 冻融-水蚀凹陷中的土壤呼吸日间变化
高寒沼泽草甸5月份的土壤呼吸值在0.01~0.38 μmol/(m2·s)范围内,一天中土壤呼吸最强的时段处于14:00左右。白天中6:00左右的土壤呼吸速率最弱,之后随着空气温度及土壤温度的升高而升高,在14:00时左右呼吸速率达到最高峰,之后呼吸速率开始变小。冻融—水蚀凹陷土壤呼吸速率总体比非冻融—水蚀凹陷中的土壤呼吸速率显著低于非冻融—水蚀凹陷的土壤呼吸(p<0.05,n=21)(见图2)。
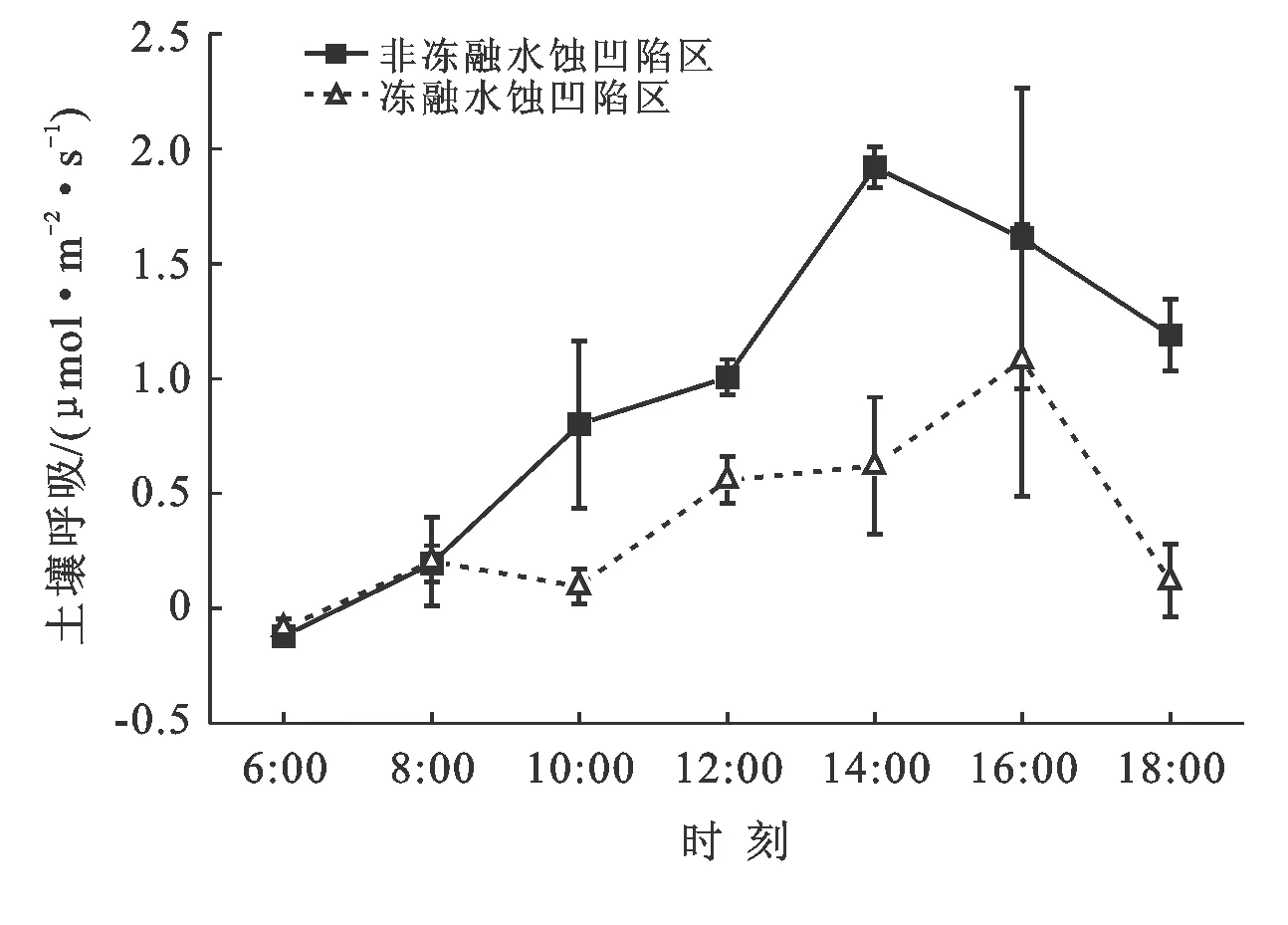
图2 冻融-水蚀凹陷与非冻融-水蚀凹陷样区的土壤呼吸在5月份白天的变化
3.3 冻融-水蚀凹陷环境要素变化与土壤呼吸
土壤呼吸的变化主要受制于土壤和大气的水热条件的变化。从表2可以看出空气温度、空气相对湿度、5 cm土壤含水量与土壤呼吸显著相关(p<0.01,n=42)。土壤呼吸的变化主要受土壤温度和土壤含水量的影响,但本次研究中5 cm土壤温度与土壤呼吸的相关性不显著(见表2)。
通过逐步回归模型分析空气温度,空气相对湿度,5 cm土壤温度及5 cm土壤含水量对土壤呼吸的影响,所构建的逐步回归模型能够解释27.5%的土壤呼吸的影响(R2=0.275)。
y=-0.028x+2.008 (R2=0.275)
(1)
式中:y——土壤呼吸速率;x——空气相对湿度。

表2 青藏高原冻融-水蚀凹陷气象要素及土壤呼吸相关性分析
注:**表示在0.01 水平(双侧)上显著相关。
逐步回归模型结果表明空气相对湿度显著影响土壤呼吸速率(λ=-0.028,p<0.01),但空气温度、5 cm土壤温度和5 cm土壤含水量对土壤呼吸速率没有显著的影响。
土壤呼吸过程复杂,影响因子多样。主要的有地域气象要素(空气温度、相对湿度、降水)、土壤理化性质(土壤温度、土壤含水量、土壤有机质含量)、土壤微生物及人类活动。以上要素相互影响、相互作用,且随着时间的变化而变化。因此,无法定量评估单一要素对土壤呼吸的作用[8],冻融-水蚀凹陷中土壤呼吸的变化是多种影响因子共同作用的结果。
4 讨论与结论
4.1 讨 论
(1) 冻融—水蚀凹陷对土壤呼吸的影响。高寒沼泽草甸5月份的土壤呼吸随着空气温度及土壤温度的升高而升高,6:00左右的土壤呼吸速率最弱,在14:00左右呼吸速率达到最高峰,之后呼吸速率开始变小。冻融—水蚀凹陷土壤呼吸速率总体比非冻融—水蚀凹陷中的土壤呼吸速率显著低于非冻融—水蚀凹陷的土壤呼吸47.99%(p<0.05,n=21)。影响土壤呼吸的因子复杂多样,且不同因子之间存在相互作用[13]。例如土壤理化性质(温度、含水量、有机质含量、土壤孔隙度等)、土壤养分状况、植被类型与地表覆盖、地域气候(气温和降水)及人为活动造成的土地利用方式的改变等。多个因子之间对土壤呼吸的相互作用可能会随着时间的变化而变化[14]。
(2) 冻融—水蚀凹陷中环境要素变化对土壤呼吸的影响。在高寒沼泽草甸生态系统中土壤温度、水分是影响土壤呼吸的主要因子,空气温度和相对湿度通过影响土壤表层作用于土壤呼吸。冻融沼泽湿地土壤温度与土壤CO2排放通量呈显著指数相关[8]。土壤温度降低会导致某些微生物数量下降,影响微生物呼吸,从而导致土壤呼吸随之减弱[8]。在本文的研究中土壤呼吸和5 cm土壤温度的变化具有一致的单峰型变趋势,10:00以后的5 cm土壤温度上升快,土壤呼吸速率随之上升也快,12:00左右土壤温度和土壤呼吸均达到一天中最大,16:00后土壤温度减低,土壤呼吸减弱。利用相关分析得出土壤呼吸与5 cm土壤温度相关性不显著,可能是由于本次研究的数据量有限影响。已有研究表明在一定范围内土壤呼吸随着土壤水分含量的增加而降低[15-16]。土壤冻融交替增强了土壤的保水性[8]。在冻结过程中5—40 cm深度处土壤含水量会下降,融化过程中5—10 cm深度处土壤体积含水量呈现上升趋势[17]。在冻融循环过后,大的土壤团聚体被粉碎成小团聚体,小团聚体随土壤水流而运动,填埋原有及新形成的孔隙,土壤孔隙度变小[18]。土壤孔隙度变小导致土壤的下渗能力减弱,土壤含水量增大。本次测定的高寒沼泽草甸的冻融—水蚀凹陷样区的土壤含水量与非冻融—水蚀凹陷样区差别不大。主要原因是样地为季节性水淹湿地。然而土壤水分含量仍然对土壤呼吸有显著的影响,本研究中土壤呼吸与土壤含水量呈显著的负相关关系,与空气温度和相对湿度也极显著。
冻融—水蚀凹陷中的土壤呼吸低于非冻融—水蚀凹陷样区,除了空气温度、空气相对湿度、5 cm土壤温度和土壤含水量的变化以外可能还与冻融—水蚀凹陷造成的微生物变化、植被特征的变化有关。研究表明冻融循环能够使大量微生物死亡,死亡的微生物释放出营养物质和易降解有机物,短暂刺激参与微生物的活性,进而增强异养呼吸[19]。但长期性的冻融循环使土壤微生物数量持续减少,分解有机碳能力下降[20],也使土壤微生物生理活性和群落结构发生变化,进而抑制土壤微生物量对土壤有机碳的分解能力[21]。由此可见,不论是冻融循环过程还是长期性冻融—水蚀形成的凹陷,微生物群落结构及种群数量将会发生较大变化,土壤微生物控制下的土壤异养呼吸发生改变,进而会影响土壤呼吸。高寒沼泽草甸冻融—水蚀不仅改变了植物群落物种多样性,降低了植物群落盖度、高度及生物量,并且土壤有机质含量也会随着冻融—水蚀程度的加剧逐渐降低[5]。季节性冻融为高寒植物生长提供良好的土壤水分,对植物初期生长发育有利,可弥补春夏之交时降水不足引起的干旱胁迫影响;季节冻土的长时间维持,有利于植物残体和土壤有机质留存于土壤,并随土壤冻结和融化而迁移,进而提高土壤肥力[14]。而在季节性冻融作用和水蚀作用造成的凹陷微地貌中,植被变化可以在一定程度上直接反映微生态系统的变异过程。研究表明随着高寒草甸的冻融—水蚀作用的加剧,豆科和菊科等杂草植物入侵群落,降低了群落高度和盖度,莎草科和禾本科优良牧草逐渐减少,地上生物量降低[5]。在本次研究中冻融—水蚀造成地表下陷9.193 cm,地上生物量降低了60.4%,小嵩草的高度降低了68.7%,群落盖度降低了16.9%。主要的原因可能是因为高寒草甸冻融—水蚀作用会降低土壤有机质含量,导致植物养分供给量减少,影响地上生物量;且随着冻融—水蚀程度加剧,植物高度和植物群落盖度逐渐降低,地表裸露比例逐渐增加,地上生物量会持续降低[5]。因此,季节性冻融循环过程有利于植物生长,但冻融—水蚀后的微地貌环境由于土壤养分的流失、水热分配条件的变化等,可能对原生种群植物来说则不是最佳的生境。
4.2 结 论
高寒沼泽草甸长期性冻融—水蚀使地表凹陷9.193 cm,土壤呼吸速率减弱了47.99%(p<0.05,n=21)。冻融—水蚀凹陷中的空气温度、空气相对湿度、5 cm土壤温度、5 cm土壤含水量的变化影响土壤呼吸。高寒沼泽草甸长期性缓慢且持续不断的冻融—水蚀形成的凹陷中土壤呼吸及其影响因子发生明显变化,改变了原生高寒草甸生态系统的碳循环。在本次研究中尚未探测冻融—水蚀凹陷对原生生态系统的影响强度,但冻融—水蚀凹陷的强度、凹陷形成的时间及未来的发展方向不容忽视。不同深度的冻融—水蚀凹陷对土壤呼吸的影响不同,不同深度的冻融水蚀凹陷中土壤养分、土壤微生态群落及种群、植被特征等状况相差甚大。在未来的研究中深入开展不同凹陷程度中的土壤呼吸与之影响因子探究,不仅可准确估算高寒沼泽草甸冻融—水蚀对土壤碳循环的影响,而且可以预测冻融—水蚀凹陷造成的草地退化对未来气候的影响,为青藏高原生态系统碳循环模型研究提供科学合理的补充。
冻融—水蚀凹陷形成的微生态系统中土壤温度、水分发生较大变化,使冻融循环作用不同于原生生系统。冻融循环作用影响下的土壤化学循环、土壤微生物及其活性相继发生改变,进而影响土壤碳库。因此,对高寒草甸冻融—水蚀凹陷中的土壤冻融循环特征研究尤为关键。
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
海外文摘·文学版(2021年3期)2021-05-19
水土保持研究(2019年6期)2019-10-19
活力(2019年21期)2019-04-01
下一代英才(酷炫少年)(2017年9期)2017-11-27
中国水土保持科学(2017年3期)2017-07-05
剑南文学(2016年11期)2016-08-22
浙江大学学报(工学版)(2016年2期)2016-06-05
时代英语·高三(2014年5期)2014-08-26
