
小麦叠氮化钠诱变群体产量及相关性状遗传特性分析
2020-04-10 06:57王伟王斌曹平平
江苏农业科学 2020年3期
王伟 王斌 曹平平

摘要:以含有72个品系的沧麦6005叠氮化钠诱变群体为试验材料,调查该群体的产量、千粒质量、穗粒数等产量相关农艺性状,分析该群体农艺性状的遗传多样性及相关性,并估计其遗传参数。结果表明,每穗小穗数的遗传多样性指数最高,为1.444 1,其后依次是穗粒数>亩穗数>千粒质量>产量>穗长,穗粒数的变异系数最大,为16.08%,每穗小穗数变异系数最小,为6.54%。简单相关分析结果表明,穗粒数、千粒质量、产量与株高均极显著正相关;产量和穗粒数、千粒质量之间极显著正相关。偏相关分析结果表明,产量和株高、亩穗数、千粒质量之间显著或极显著正相关。穗粒数、株高和产量的选择潜力较大,更有可能从该群体中选出偏离平均数大的家系类型。
关键词:小麦;叠氮化钠诱变;产量性状;遗传特性
中图分类号: S512.103.52 文献标志码: A 文章编号:1002-1302(2020)03-0102-04
小麦在全球的种植面积广泛,在我国农业生产中占重要地位。小麦品种的遗传脆弱性是小麦育种不易突破的瓶颈之一。诱变育种是小麦新品种选育和种质资源创新的重要手段。化学方法诱发基因突变是小麦遗传改良的重要途径之一[1]。叠氮化钠(NaN3)是应用于植物化学诱变的高效低毒的化学诱变剂之一。叠氮化钠已在小麦[2-3]、玉米[4]、水稻[5]、大麦[6-7],大豆[8]等农作物上加以应用。另外,张希太等研究小麦叠氮化钠诱变后代在株高、芒型、穗型等的变异特征,并从分子水平证明了叠氮化钠对小麦的诱变效果[9-11]。但还鲜有应用叠氮化钠构建小麦突变体库的报道。
小麦是国际主要粮食作物之一。小麦产量性状为数量性状,受多基因和多种环境因素影响,而且性状间也存在互相联系和制约。小麦的产量取决于很多内、外环境因素,从农艺学的角度来讲,主要与株高、小穗数、穗粒数、千粒质量和小区产量[12-13]等农艺性状有关。小麦产量构成三因素中,亩穗数对产量的影响最大,调节能力最强,其次对产量贡献较大的是穗粒数[14]。有关小麦产量构成因素对产量影响大小,前人虽有研究,但因材料或地区不同,分析结果有所不同[15-16]。黑龙港流域是国家重要的冬小麦种植区之一。沧麦6005是沧州市农林科学院育成的抗旱耐盐碱能力突出的一个囯审小麦品种。本研究以72个沧麦6005叠氮化钠诱变的突变体作为研究对象,对小麦产量构成因素相关性状的遗传多样性及与产量的相关性进行分析,并对有关性状作为遗传潜力进行探讨,旨在为黑龙港流域盐碱地小麦高产育种提供参考。
1 材料与方法
1.1 试验材料
试验材料为沧麦6005叠氮化钠诱变突变体,共72个,2017年10月种于沧州市农林科学院前营试验站,2018年6月收获。同期调查株高、穗长、每穗小穗数、亩(1亩=667 m2)穗数、穗粒数、千粒质量和产量。本试验采用完全随机试验设计,重复2次。小麦种植期间,其他栽培管理措施同于大田。
1.2 试验方法
NaN3由美国西格玛(Sigma)公司生产。参照NaN3在大麦中的处理方法[6]处理小麦种子:待处理沧麦6005种子浸入自来水,先后在4 ℃浸泡16 h,20 ℃浸泡 4 h,然后加入适量1 mol/L KH2PO4(pH值=3),摇匀。于通风橱中加入5.0 mL 1 mol/L NaN3,混合充分。20 ℃轻柔振动2 h,处理结束后在自来水下彻底冲洗1 h,于通风橱中用纸巾晾干种子,过夜。废液用过量次氯酸钠处理后倾倒于水槽。
1.3 分析方法
数据采用Excel和SPSS软件进行统计分析,不同品系间性状的差异采用变异系数表示,遗传多样性指数采用Shannon-Weaver信息指数,计算公式:H′=-∑PilnPi,其中Pi为某一性状第i个级别出现的概率[17-18]。为了便于统计分析,将小麦产量相关性状进行分级,进而计算遗传多样性指数,同时进行相关分析及诱变群体遗传参数估计[19]。
2 结果与分析
2.1 沧麦6005叠氮化钠诱变群体产量及相关性状的描述性统计分析
由沧麦6005叠氮化钠诱变群体产量相关性状的基本统计结果(表1)表明,每穗小穗数的遗传多样性指数最高,为1.444 1,其后依次是穗粒数>亩穗数>千粒质量>产量>穗长>株高。不同品系间的变异系数存在很大差异,穗粒数、产量和株高的变异系数较大,分别为16.08%、14.15%和 13.47%,变幅分别为16.50~39.20、187.66~418.79 和35.00~82.00;其次为亩穗数和千粒质量,变异系数为11.25%和11.39%,变幅分别为32.00~55.20和20.60~42.60;穗长和每穗小穗数的變异系数较小,分别为9.87%和6.54%,变幅分别为5.50~9.34和12.30~17.70。
2.2 沧麦6005叠氮化钠诱变群体产量及相关性状的简单相关分析
对沧麦6005叠氮化钠诱变群体产量相关性状进行简单相关分析,结果(表2)表明,穗粒数、千粒质量、产量与株高均极显著正相关;穗长和小穗数、穗粒数极显著正相关;穗粒数和小穗数极显著正相关,穗粒数和亩穗数之间极显著负相关;亩穗数和千粒质量之间极显著负相关;产量和穗粒数、千粒质量之间极显著正相关;其他性状两两间相关关系不显著。研究结果说明,小麦产量的3个构成因素亩穗数、穗粒数和千粒质量之间存在制约关系,另外,株高对产量构成因素的影响也不容忽视。
2.3 沧麦6005叠氮化钠诱变群体产量及相关性状的偏相关分析
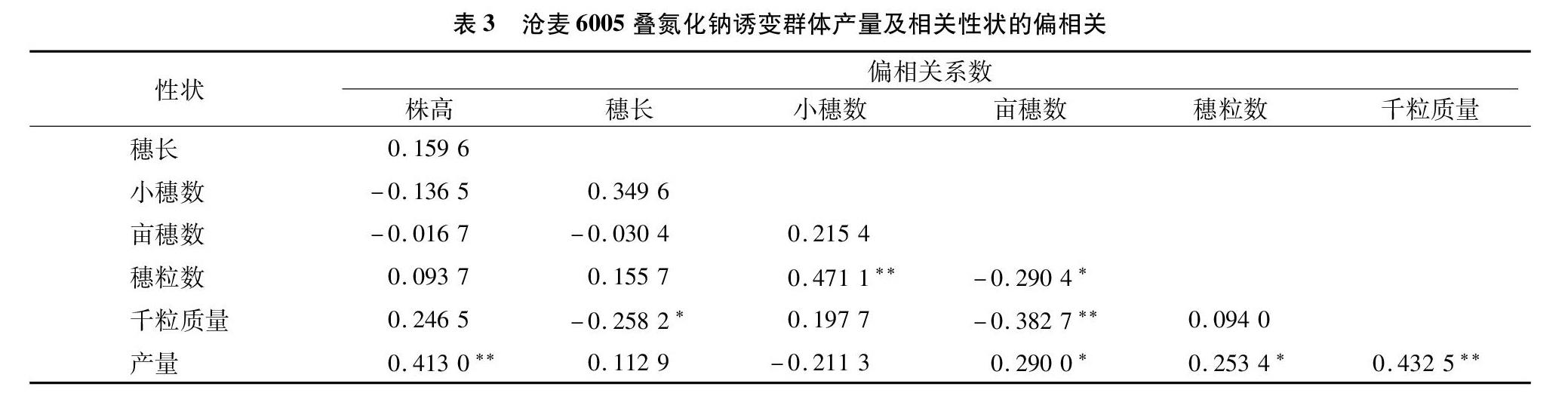
对沧麦6005叠氮化钠诱变群体产量相关性状进行偏相关分析,结果(表3)表明,产量和株高、亩穗数、千粒质量之间显著或极显著正相关;穗长和千粒质量,亩穗数和穗粒数、千粒质量之间显著或极显著负相关;小穗数和穗粒数、亩穗数和产量、穗粒数和产量、千粒质量和产量之间显著或极显著正相关;其他性状两两之间偏相关系数均不显著。
2.4 沧麦6005叠氮化钠诱变群体产量及相关性状的遗传参数
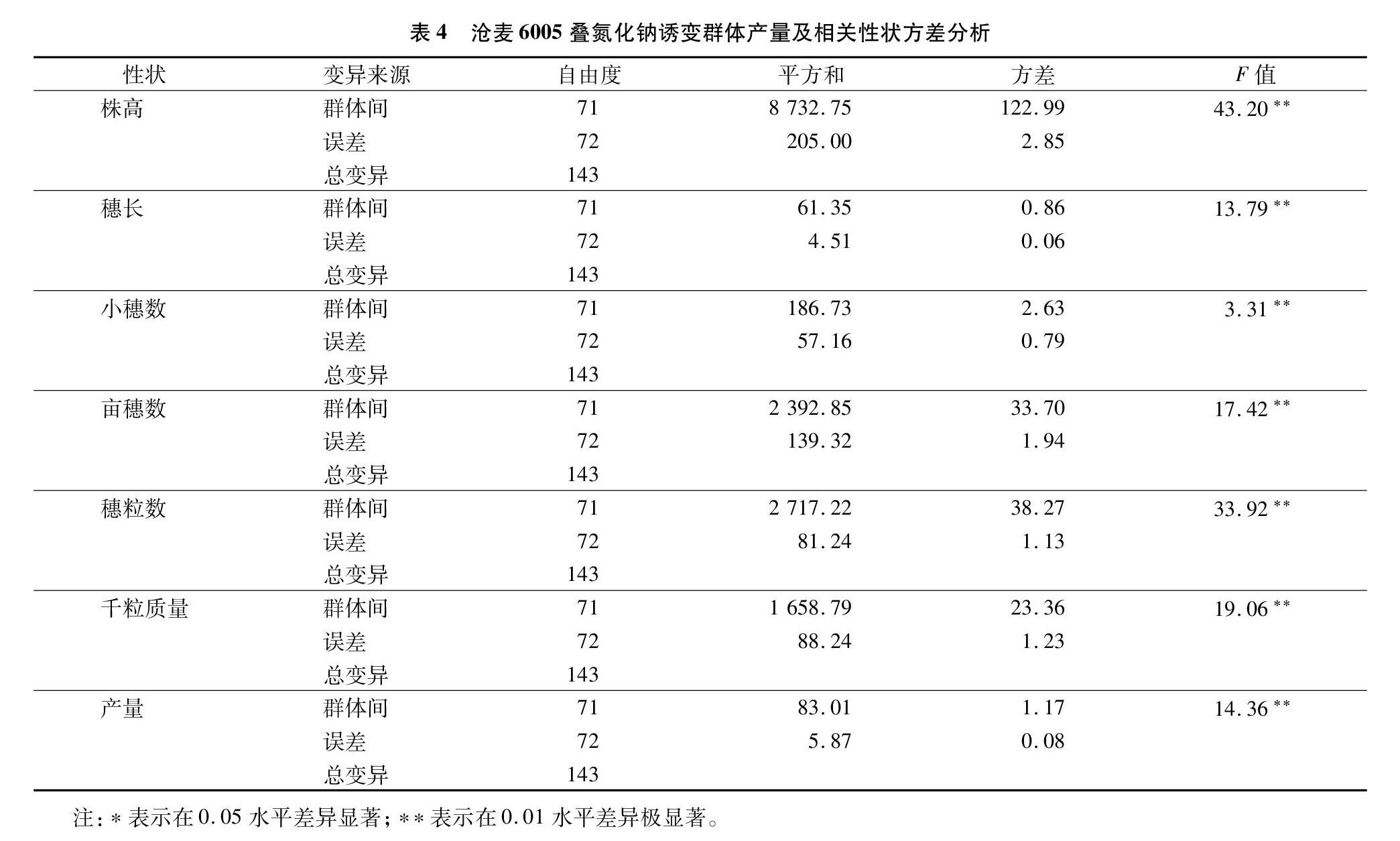
对72个诱变群体的产量及相关性状进行方差分析,结果(表4)表明,产量及其相关性状群体间均差异极显著。进而依据随机模型估计该群体的遗传参数(表5)。其中,株高、穗长、小穗数、亩穗数、穗粒数、千粒质量和产量的广义遗传力(h2b)分别为 0.95、0.86、0.54、0.89、0.94、0.90和0.87,除每穗小穗数外其他性状广义遗传力均比较高,说明除每穗小穗数外其他6个产量相关性状的变异绝大部分是由遗传原因造成的。穗粒数的遗传变异系数(GCV)最高,为15.03%,其后依次为株高>产量>千粒质量>亩穗数>穗长>每穗小穗数,每穗小穗数遗传变异系数最小,为6.11%。穗粒数、株高、产量的选择潜力相对较大,更有可能从该群体中选出偏离其平均数大的家系类型。
3 结论与讨论
在小麦的产量构成三要素中,亩穗数和穗粒数在一定范围内相互协调,小麦亩穗数和千粒质量也只在一定范围内有联系,穗粒数和千粒质量之间相对独立。在本研究中,亩穗数、穗粒数和千粒质量之间存在制约关系,与近年相关研究结论[20-21]一致。另外, 穗粒数与产量的简单相关系数和偏相关系数显著或极显著存在,说明穗粒数是影响小麦产量的主要因素,穗粒数对产量有很大的直接作用,这与姚国才等研究结果[22]一致。但是本研究亦表明,株高对产量构成因素的影响也不容忽视。
叠氮化钠是一种高效且无毒害作用的植物化学诱变剂。利用叠氮化钠进行化学诱变的方法是创造小麦新种质、选育小麦新品种的有效途径之一,在扩大遗传变异、加速提高育种效率、改进育种方法等方面有着很大发展潜力。本研究提及的72个叠氮化钠诱变群体的株高、穗长、亩穗数、穗粒数、千粒质量和产量性状均具有较高的广义遗传力,说明除每穗小穗数外其他6个产量相关性状的变异绝大部分是由遗传原因造成的。穗粒数的遗传变异系数最高,其后依次为株高>产量>千粒质量>亩穗数>穗长>每穗小穗数,这与前面计算的变异系数基本一致。说明穗粒数、株高、产量的选择潜力相对较大,更有可能从该群体中选出偏离其平均数大的家系类型。
参考文献:
[1]王琳清,陈秀兰,柳学余. 小麦突变育种学[M]. 北京:中国农业科学技术出版社,2004:35-88.
[2]Cheng X Y,Gao M W,Liang Z Q. Effect of mutagenic treatments on somaclonal variation in wheat (Triticum aestivum L.)[J]. Plant Breeding,1990,105(1):47-52.
[3]张超美. 叠氮化钠对小麦的诱变效应[J]. 湖北农学院学报,1994,14(3):56-60.
[4]Kiruki S,Onek L A,Limo M. Azide-based mutagenesis suppresses Striga hermonthica seed germination and parasitism on maize varieties[J]. African Journal of Biotechnology,2006,5(10):866-870.
[5]Gao M W,Cai Q H,Liang Z Q. In vitro culture of hybrid indica rice combined with mutagenesis[J]. Plant Breeding,1992,108(2):104-110.
[6]Talame V,Bovina R,Sanguineti M C,et al. TILLMore,a resource for the discovery of chemically induced mutants in barley[J]. Plant Biotechnology Journal,2008,6(5):477-485.
[7]曹 欣,杨煜峰,钱强华. 叠氮化钠对不同大麦品种的诱变效应[J]. 浙江农业学报,1991,3(3):143-146.
[8]姜振峰,刘志华,李文滨,等. 疊氮化钠对大豆M1的生物学诱变效应[J]. 核农学报,2006,20(3):208-210.
[9]张希太. NaN3诱变小麦矮抗58后代变异的研究及SSR分析[J]. 作物研究,2011,25(3):189-192.
[10]张希太,谢淑芹,张彦波,等. 利用NaN3诱变中育5号选育变异种质系及SSR分析[J]. 西南农业学报,2011,24(4):1239-1242.
[11]张希太. NaN3诱变小麦山农8355后代变异的研究及SSR分析[J]. 农业与技术,2011,31(2):38-42.
[12]杨春玲,关 立,侯军红,等. 黄淮麦区小麦产量构成因素效用研究[J]. 山东农业科学,2007(4):19-23.
[13]刘兆晔,于经川,杨久凯,等. 小麦生物产量、收获指数与产量关系的研究[J]. 中国农学通报,2006,22(2):182-184.
[14]郭春强,黄 杰,曹燕燕,等. 小麦新品种漯麦6010稳产性、产量构成因素变异性及通径分析[J]. 湖北农业科学,2017,56(4):608-610.
[15]王继滨,李慧东,李凤云. 我国小麦区域试验中的问题与对策[J]. 作物杂志,2010(4):87-91.
[16]郭凌云. 限量灌溉下不同密度对冬小麦产量和水分利用效率的影响[J]. 山东农业科学,2010(7):51-52.
[17]张礼凤,李 伟,王彩洁,等. 山东大豆种质资源形态多样性分析[J]. 植物遗传资源学报,2006,6(4):450-454.
[18]郝黎仁,樊 元,郝哲欧,等. SPSS实用统计分析[M]. 北京:中国水利水电出版社,2002:280-285.
[19]莫惠栋. 农业试验统计[M]. 上海:上海科学技术出版社,1984.
[20]陈剑锋,陈登耀,张秋英. 小麦主要性状与产量的通径和灰色关联度分析[J]. 福建稻麦科技,2011,29(4):5-8.
[21]佟汉文,黄荣华,刘易科,等. 小麦新品种农艺性状与产量的相关及通径分析[J]. 湖北农业科学,2008,49(7):758-760.
[22]姚国才,姚金保,杨学明,等. 长江中下游小麦品种产量性状的遗传相关和通径分析[J]. 南京农专学报,2002,18(4):11-14.
猜你喜欢
今日农业(2022年16期)2022-11-09
今日农业(2021年12期)2021-11-28
今日农业(2021年8期)2021-11-28
金桥(2021年10期)2021-11-05
今日农业(2021年13期)2021-08-14
作文小学中年级(2020年4期)2020-06-11
农家科技下旬刊(2016年9期)2016-12-15
山东农业科学(2014年1期)2015-03-09
山东农业科学(2014年2期)2014-03-24
