
高强度间歇训练通过支配内脏脂肪的交感神经活动促进白色脂肪组织棕色化
2020-04-06 01:03董高芳赵晓博何玉秀张海峰
中国体育科技 2020年2期
刘 阳 ,张 赛 ,董高芳 ,赵晓博 ,何玉秀 ,张海峰 *
肥胖是现代人健康的主要威胁之一。体内约80%的甘油三酯储存于皮下脂肪组织,仅有约20%储存于内脏脂肪组织,但内脏脂肪堆积是心血管及代谢类疾病的独立风险因素(Shafqat et al.,2018)。运动是减肥的有效方法之一,高强度间歇训练(high intensity interval training,HIIT)与传统的中等强度持续训练(moderate intensity continuous training,MICT)相比,减体重效果相同,但能明显节省时间(Zhang et al.,2017)。HIIT的另一优势是可减少更多内脏脂肪(Maillard et al.,2018)。HIIT减少内脏脂肪的机制与内脏脂肪对促脂解类激素的敏感性更高有关(Maillard et al.,2016)。前期研究发现,HIIT可通过激素敏感性脂肪酶(hormone-sensitive lipase,HSL)的激活,促进更多内脏脂肪甘油三酯水解(刘阳等,2018)。但甘油三酯被HSL水解后产生的游离脂肪酸(free fat acid,FFA)如何进一步分解氧化,即HIIT较MICT是否能消耗更多内脏脂肪水解释放的FFA,尚待深入研究。
脂肪组织包括白色脂肪组织(white adipose tissue,WAT)与棕色脂肪组织(brown adipose tissue,BAT)两类。WAT的主要功能是储存甘油三酯,BAT则是机体重要的非颤栗产热器官,其线粒体内膜上的解偶联蛋白1(Uncoupling Protein-1,UCP-1)可抑制ATP的合成,使FFA以热能的形式消耗来调节体温。普通情况下,WAT的线粒体浓度和 UCP-1表达远低于 BAT(Orava et al.,2011),但近年来发现WAT中会分化出一些UCP-1高表达的脂肪细胞,被认为是WAT“棕色化”(Browning)后向BAT过渡的中间型(Wu et al.,2012),将命名为米色脂肪细胞(Beige Adipocyte)。WAT在寒冷、饥饿或运动刺激后,会出现UCP-1表达增加的棕色化现象,可脂肪细胞内部直接消耗FFA,所以WAT棕色化被认为是一条较有潜力的减肥途径(Jeremic et al.,2016)。
WAT棕色化的标志是UCP-1表达升高,细胞内外的多个复杂信号共同调控了此过程(Jeremic et al.,2016)。有研究认为,支配脂肪组织的交感神经纤维,通过突触向脂肪细胞分泌神经递质去甲肾上腺素,是刺激WAT棕色化的主要途径(Nedergaard et al.,2014)。但 Boström 等2012年发表在Nature的研究显示,运动激活骨骼肌过氧化物酶体增殖物激活受体γ辅激活因子1α(peroxisome proliferator-activated receptor gamma coactivator 1-alpha,PGC-1α)后,可诱导肌细胞合成纤维连接蛋白Ⅲ型结构域蛋 白 5(fibronectin type Ⅲ domain-containing protein 5,FNDC5),FNDC5经剪切生成鸢尾素(irisin)并分泌入血,运输至WAT后可显著提高UCP-1表达(Boström et al.,2013)。由于骨骼肌与运动的关系极为密切,肌肉因子Irisin因促进WAT棕色化成为近年来运动减肥的研究热点。
研究显示,急性HIIT训练比MICT更显著激活骨骼肌FNDC5/Irisin(Archundia et al.,2017),说明 HIIT 有通过肌肉因子分泌途径促进更多WAT棕色化的潜力。然而,在HIIT促进WAT棕色化的过程中,交感神经活动与肌肉Irisin分泌这2条途径的作用重要性,尚未有研究进行比较。本研究的第一部分,对高糖高脂膳食诱导的肥胖C57BL/6小鼠进行12周的HIIT与MICT训练,比较两者对体重、内脏与皮下脂肪垫重量、脂肪细胞形态及WAT棕色化的影响,从脂肪细胞自身产热消耗FFA的角度,研究HIIT减少更多内脏脂肪的机制。同时观察了促进UCP-1表达的2条上游信号:代表交感神经活动的下丘脑阿片促黑素皮质素原(proopiomelanocortin,POMC)与脂肪交感神经酪氨酸羟化酶(tyrosine hydroxylase,TH),以及代表Irisin分泌的骨骼肌PGC-1α与FNDC5,对比HIIT是否通过交感神经活动亦或肌肉因子分泌途径促进了WAT棕色化。分析研究一的数据后发现,TH与内脏WAT的UCP-1表达呈显著相关,故研究通过特异性的拮抗剂SR59230a,抑制内脏WAT接受交感神经调控的β3肾上腺素能,验证并确定HIIT可通过支配内脏脂肪的交感神经促进WAT棕色化。
1 研究对象与方法
1.1 研究一:MICT与HIIT促进不同部位脂肪棕色化的效果及上游调控通路的对比
1.1.1 实验动物分组与肥胖模型建立
由于本团队以往关于HIIT减肥效果的研究均选取肥胖女性青年群体作为研究对象,同时根据Maillard等(2018)的研究,性别差异不会对HIIT减肥效果造成影响,为与人体研究保持一致,本实验采用雌性c57bl/6小鼠作为研究对象。首先对小鼠进行高糖高脂膳食饲养,建立肥胖动物模型:3周龄小鼠(体重9.67±1.12g,北京维通利华)随机分为标准膳食对照组C(n=6只)和高糖高脂膳食诱导的肥胖建模组(高糖高脂饲料包含60%标准饲料+16%白砂糖+5%猪油+18%蛋黄粉+1%胆酸钠)。饲养15周后,选取肥胖建模组(体重超过C组10%以上)小鼠18只,进一步随机分为高糖高脂膳食肥胖对照组(HFD)、中等强度持续训练组(MICT)以及高强度间歇训练组(HIIT),每组各6只。在饲养与训练过程中,室温始终保持20℃~22℃。
1.1.2 训练方案
肥胖动物模型建立后,给予MICT与HIIT组小鼠12周25°上坡跑训练,HFD作为对照组不进行训练,3组在训练期间均保持高糖高脂饲养。MICT组单次运动的跑速恒定,第1周跑速13 m/min,并在5周后逐步增加至17 m/min。HIIT组进行“高强度1 min,低强度2 min”为一循环的间歇训练,间歇期的中强度跑速与MICT相同,高强度跑速由第1周的23 m/min逐步递增至12周的30 m/min。以上运动强度参考了Høydal等(2007)的研究确定:MICT组强度始终保持在50%~70%最大摄氧量,HIIT组高强度约90%~110%最大摄氧量。MICT组一次训练时间45 min,HIIT组小鼠在奔跑距离达到MICT组水平后,停止该次训练。由于跑速更高,HIIT组完成一次训练的时间在25 min左右,远低于MICT组(刘阳 等,2018)。
1.1.3 动物取材
12周训练结束48 h后取材,取材前禁食12 h。腹腔注射4%戊巴比妥钠麻醉,生理盐水动脉灌流后,剥离完整的子宫周围脂肪垫(代表内脏脂肪)和腹股沟皮下脂肪垫(代表皮下脂肪)称重,剥离腓肠肌并剔除结缔组织后取肌腹部分。Western blot测试所需组织在-80℃冻存,H.E.染色所需组织以4%多聚甲醛固定。
1.1.4 脂肪组织H.E.染色
组织从固定液取出,修平,梯度酒精脱水,二甲苯透明处理,浸石蜡包埋。5 μm切片,修整后38℃铺片机展片,60℃烘箱烤片。切片梯度酒精复水后,苏木精染色5 min,自来水冲洗,1%盐酸酒精分化,自来水冲洗,0.6%氨水返蓝,伊红染色2 min,梯度酒精脱水,中性树胶封片。
1.1.5 骨骼肌PGC-1α、FNDC5、脂肪TH、UCP1蛋白表达
使用Western blot法对蛋白表达进行测试。样品使用RIPA裂解30 min(骨骼肌与裂解液1:10,脂肪与裂解液1:5),离心取上清。BCA法测定蛋白浓度后煮沸5 min。加入Sample buffer后以每孔20 μl上样。采用4%浓缩胶,10%分离胶电泳。湿转法转至PVDF膜,脱脂奶粉封闭2 h,一抗4℃孵育过夜(一抗浓度PGC-1α与FNDC5为1:2 000,UCP-1为1:1 000,TH为1:10 000,一抗均购于Abcam)。TBST洗5次后二抗孵育2 h,TBST洗3次后发光。骨骼肌内参选用Tubulin,脂肪选用β-actin。Image J 1.51计算灰度值,用“目的蛋白/内参”计算蛋白表达量。
1.2 研究二:HIIT通过交感神经途径促进内脏脂肪棕色化的机制验证
1.2.1 实验动物分组与干预
经研究一的数据发现,内脏脂肪FNDC5与交感神经活动标志物TH存在显著相关性,故通过对脂肪交感神经受体的特异性抑制,来验证交感神经活动促进内脏脂肪棕色化的效果。12只雌性c57bl/6小鼠随机分为安慰剂组(Saline组,n=6)与脂肪交感神经受体抑制组(SR组,n=6)。两组均进行10天HIIT训练,训练方法同研究一。在每次训练前2 h,SR组腹腔注射β3肾上腺素能受体拮抗剂 SR59230a( 购于Sigma,剂量为1 mg/kg,Sipe et al.,2017)。由于交感神经对脂肪组织的调控主要是通过轴突末梢分泌的去甲肾上腺素作用于脂肪细胞的β3肾上腺素能受体实现的(Bartness et al.,2014),故SR组在进行HIIT训练时,内脏脂肪的交感神经调控已经被阻断。10天HIIT训练结束后,取Saline组与SR组小鼠内脏脂肪,测试UCP-1表达水平。需指出的是,研究一的数据显示,仅有内脏脂肪UCP-1表达与TH呈显著相关,同时无法确定本研究二中所采用的化学抑制方法是否能对腹股沟皮下WAT的交感神经作用产生抑制[腹股沟皮下脂肪交感神经的抑制,应采用腹股沟皮下注射抑制来实现(Dodd et al.,2015)],故研究二仅对内脏脂肪UCP-1表达进行测试。由于c57bl/6小鼠的动情周期仅4~5天(Byers et al.,2012),10天内雌性小鼠经历2个完整周期,同时性激素的变化不会对白色脂肪棕色化产生明显影响(Dixen et al.,2013),所以小鼠动情周期的个体差异不会对研究二的结果产生明显影响。
1.2.2 内脏脂肪UCP-1表达
采用Western blot法测试小鼠子宫周围内脏UCP-1表达,具体方法同研究一。
1.3 统计学方法
研究一在建模阶段使用单因素方差分析计算对照组C与肥胖建模组的体重差异。在训练干预阶段,将摄食量作为协变量使用协方差分析比较小鼠体重及脂肪重量差异,以消除摄食量对以上指标的影响。使用单因素方差分析比较蛋白表达差异。Pearson法分析UCP-1与TH、FNDC5的相关性,双尾检测显著性,R≥0.8为高度相关,0.8>R>0.3为中度相关,R≤0.3为低度相关。研究二使用单因素方法分析比较体重、脂肪重量与蛋白表达的差异。以上显著性均取P<0.05为显著,P<0.01为非常显著。数据使用SPSS 24.0进行统计分析。
2 实验结果
2.1 研究一:MICT与HIIT促进不同部位脂肪棕色化的效果及上游调控通路的对比
2.1.1 HIIT与MICT对体重、子宫周围与腹股沟皮下脂肪重量的影响
在建模阶段的第4~15周,肥胖建模组小鼠的体重显著高于C组(P<0.05或P<0.01)。运动干预期间,HIIT组与MICT组小鼠体重均显著低于HFD组(P<0.05或P<0.01),HIIT组与MICT组无显著差异。在12周干预结束后,代表内脏脂肪的子宫周围脂肪垫重量,HIIT组非常显著低于MICT组和HFD组(P<0.01),代表皮下脂肪的腹股沟脂肪垫重量,HIIT组非常显著低于HFD组(P<0.01),但HIIT组与MICT组间无显著差异(图1)。

图1 研究一中各组小鼠体重变化及运动干预结束后不同部位脂肪重量Figure 1. Body Mass of Mice during Exercise Intervention and Fat Weight at the End of Exercise Intervention
2.1.2 HIIT组与MICT组对脂肪细胞形态的影响
MICT组、HIIT组经12周训练,脂肪细胞的体积较HFD组均明显缩小(图2),切片单位面积内细胞数量(由细胞核数量代表)较HFD组明显增加。同时可见,HIIT组内脏脂肪细胞体积小于MICT组,单位面积内细胞数量高于MICT组(图2B、2C)。
2.1.3 HIIT与MICT对脂肪组织棕色化标志物UCP1表达的影响
HIIT组子宫周围内脏脂肪UCP-1显著高于MICT组及HFD组(P<0.05);MICT组与HIIT组的腹股沟皮下脂肪UCP-1表达较HFD组有升高的趋势但无显著性(图3)。
2.1.4 HIIT与MICT对腓肠肌PGC-1α、FNDC5蛋白表达的影响
HIIT组PGC-1α显著高于HFD组(P<0.05),与MICT组无差异;MICT组FNDC5表达有高于HIIT组与HFD组的趋势,但由于标准差较大,无显著差异(图4)。
2.1.5 HIIT与MICT对脂肪组织交感神经标志物TH表达的影响

图2 运动干预后子宫周围与腹股沟脂肪细胞H.E.染色Figure 2. H.E.Staining of Peri-uterine and InguinalAdipocytes after Exercise Intervention
HFD组与HIIT组子宫周围内脏脂肪显著高于MICT组(P<0.05与P<0.01),HIIT组腹股沟皮下脂肪显著高于HFD组与MICT组(P<0.05与P<0.01)(图5)。
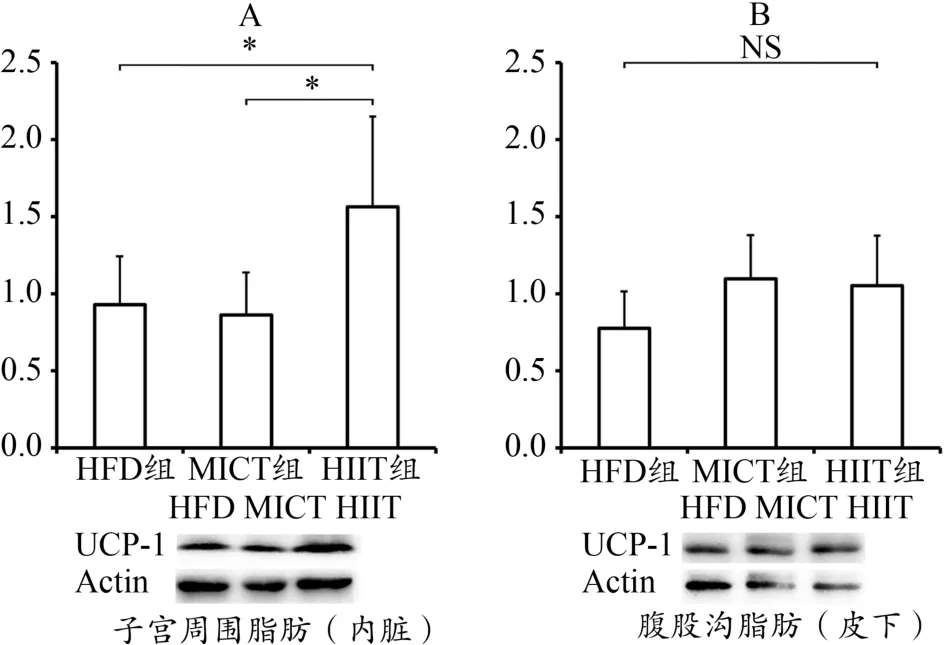
图3 子宫周围与腹股沟脂肪UCP-1表达Figure 3. UCP-1 Expression of Peri-uterine and Inguinal Fat
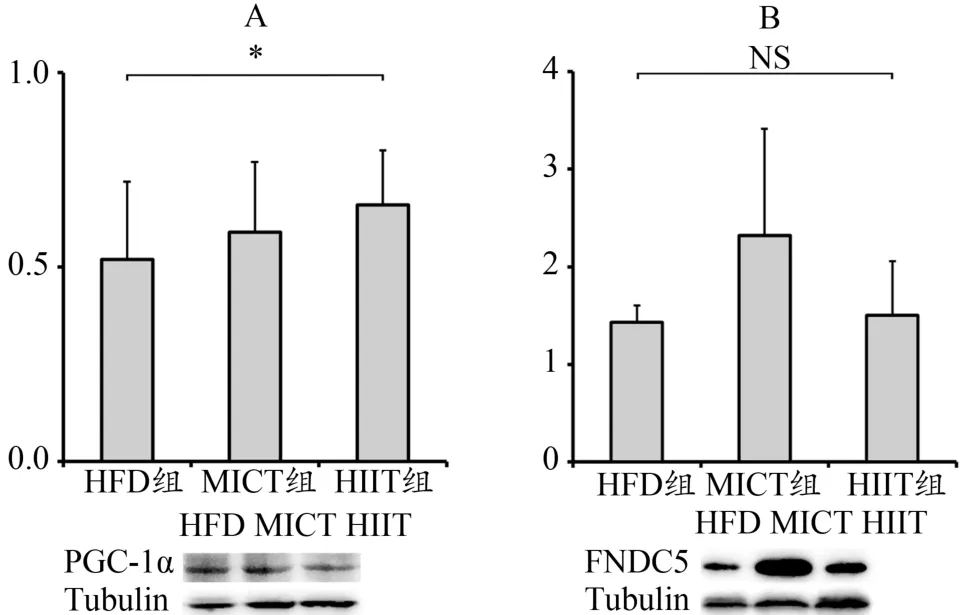
图4 腓肠肌PGC-1α与FNDC5表达Figure 4. PGC-1α and FNDC5 Expression of Gastrocnemius
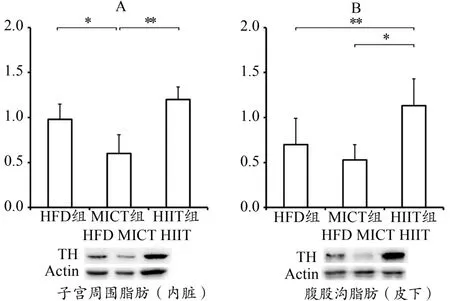
图5 子宫周围与腹股沟脂肪TH表达Figure 5. TH Expression of Peri-uterine and Inguinal Fat
2.1.6 棕色化标志物UCP-1表达与交感神经标志物TH、肌肉因子标志物FNDC5表达间的相关性分析
对内脏与皮下脂肪UCP-1分别与TH、FNDC5进行相关分析显示,内脏脂肪UCP-1表达与内脏脂肪TH间呈中度相关(R=0.578,P<0.05),皮下脂肪 UCP-1与骨骼肌FNDC5呈中度相关,但不具显著性(R=0.340,P>0.05)。其余指标间呈低度相关且不具显著性(P>0.05,图6)。
2.2 研究二:HIIT通过交感神经途径促进内脏脂肪棕色化的机制验证
SR组在注射β3型肾上腺素能受体拮抗剂SR59230a,阻断了交感神经对脂肪组织的调节途径后,进行10天HIIT训练,SR组小鼠体重有大于Saline组的趋势,但无显著性(P>0.05);内脏脂肪重量SR组有大于Saline组的趋势,但无显著性(P>0.05);SR组内脏脂肪UCP-1表达显著低于Saline组(P<0.05,图7)。
3 讨论
3.1 MICT与HIIT的降体重效果相同,但HIIT能减少更多内脏脂肪积累
在经典的能量代谢理论中,单位时间脂肪消耗绝对值最高的方式是中等强度有氧运动,所以传统观点一直认为MICT是最佳的减肥运动方式。然而MICT必须达到较长的运动时间才能获得明显的减重效果(Nielsen et al.,2011),而缺乏时间恰恰是妨碍现代人运动参与的一个重要因素(Trost et al.,2002),所以HIIT最初被用于运动减肥的目的是节省运动时间。对18~45岁超重或肥胖人群运动减肥的Meta分析显示,HIIT与MICT减轻体重的效果相同,但HIIT能节省40%以上的时间(Wewege et al.,2017)。随着HIIT研究的深入,多个系统综述与Meta分析也显示,HIIT相比MICT能使机体在心脏、血管、血压、血糖等方面获得更多健康益处(Batacan et al.,2017;Milanoviä et al.,2015;Ramos et al.,2015)。在相同运动量下,HIIT比MICT可以减少更多内脏脂肪(Maillard et al.,2018;Zhang et al.,2015),由于内脏脂肪堆积是代谢疾病的独立风险因素,此发现可以证明HIIT对改善代谢类疾病的作用。
本研究中,在奔跑距离相等的条件下,12周MICT与HIIT训练都可以显著抑制高糖高脂膳食引起的体重增长。由于跑速不同,HIIT的运动时间明显少于MICT,但HIIT组体重与MICT组无显著差异,说明HIIT在达到相同的减重效果时,确实能够节省时间。12周运动结束后,HIIT组小鼠子宫周围脂肪垫重量显著低于MICT组,腹股沟皮下脂肪较MICT有轻微降低的趋势,但不具显著性。此结果与以往的研究相符,说明运动量相同的情况下HIIT能够减少更多内脏脂肪。
3.2 HIIT相比MICT能够促进更多内脏WAT棕色化
脂肪组织的棕色化可以使以储存能量为主要功能的WAT具备一定的非颤栗性产热能力,从而能够消耗更多FFA。自被发现以来,WAT棕色化一直被认为是治疗肥胖、糖尿病等代谢疾病的关键途径(Festuccia et al.,2017)。Bartesaghi等(2014)借助基因敲除发现,人体米色脂肪细胞的产热能力完全取决于UCP-1表达,所以可使用UCP-1表达量来反映WAT棕色化的程度。寒冷刺激可促使脂肪组织UCP-1表达大幅提高(Jeremic et al.,2016),值得注意的是,运动训练对脂肪组织UCP-1表达的改变不像寒冷刺激那样剧烈。Boström等(2013)的研究在RNA转录层面上证明了中等强度运动可以剧烈增加内脏与皮下脂肪组织UCP-1 mRNA,但在蛋白表达层面上,中等强度运动引起的UCP-1升高并不显著。徐瑞等(2017)的研究显示,给予大鼠一次运动刺激,中等强度训练不能改变运动后6 h内内脏脂肪组织UCP-1表达,HIIT能提高运动后即刻UCP-1表达,但在运动后6 h即回落至安静水平,长期HIIT的作用尚未可知。Norheim等(2014)的人体研究显示,长期进行MICT虽有提升健康人和糖尿病风险人群皮下脂肪UCP-1 mRNA的趋势,但只有将健康组与风险组数据合并,UCP-1 mRNA的升高才具有显著性,且升幅不大,说明MICT促皮下脂肪棕色化的作用有限。
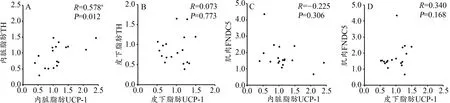
图6 UCP-1与FNDC5及TH的相关性分析Figure 6. CorrelationAnalysis among UCP-1 and FNDC5 and TH

图7 Saline组与SR组在10天HIIT训练后体重、脂肪重量及内脏脂肪UCP-1表达Figure 7. Body Mass,Fat Weight and Visceral Fat UCP-1 Expression in the Saline and SR Groups after 10-day HIIT
本研究发现,HIIT组内脏脂肪组织UCP-1显著增加,MICT组无增加。MICT组和HIIT组皮下脂肪UCP-1虽有增加趋势,但不具显著性。此结果与Norheim的研究相近,说明MICT与HIIT促皮下脂肪组织棕色化的作用不明显,但HIIT有显著的促内脏脂肪棕色化的作用。再者,研究验证了HIIT相比MICT确实能够减少更多内脏脂肪,提示HIIT减少内脏脂肪的机制与内脏脂肪组织棕色化关系密切。
3.3 长期HIIT引起的脂肪组织棕色化现象与FNDC5/Irisin信号无显著相关
2013年首次报导的Irisin被认为是一个非常有希望改善肥胖及相关代谢疾病的肌肉因子。然而此后几年内,大量关于运动是否能够提高血液Irisin浓度的研究结果却相当模糊且相互矛盾(Dinas et al.,2017)。由于血液Irisin浓度测试主要基于商业Elisa试剂盒,Albrecht等对市售的4种主流试剂盒进行了测试,发现它们均会与非特异性蛋白产生交叉反应。此外,人血浆中较低浓度的Irisin能被串联质谱法检出时,此4种试剂盒却无法测出(Albrecht et al.,2015)。Archundia等(2017)分别使用 Western blot与Elisa对肌肉和血液内的Irisin进行了检测,发现运动虽然能显著增加肌肉Irisin(WB测试结果),但血液Irisin并无变化(Elisa测试结果)。由于上述研究均质疑了Elisa方法对血液Irisin检测的灵敏性,目前国际上仅对使用串联质谱检法测血液Irisin浓度的结果较为认可。但鉴于难度与成本,研究者大多改用Western blot测试的肌肉FNDC5含量来间接反映Irisin的分泌量。
本研究同样采用肌肉FNDC5及其上游信号PGC-1α作为运动促进Irisin分泌的代表指标。与Granata等(2015)的研究相近,HIIT相比MICT能够更显著的激活PGC-1α,这说明PGC-1α可能与运动强度存在关联。当然也有某些研究发现HIIT与MICT对PGC-1α的激活无差异(Shirvani et al.,2018),这可能与不同的物种、训练方法或其他条件有关。值得注意的是,本实验中FNDC5的变化并不与PGC-1α同步:HIIT组PGC-1α虽然显著升高,但MICT组的FNDC5却具有升高的趋势,而HIIT组无变化(图4A,4B)。同时,MICT组内FNDC5个体差异很大(图4B,MICT组标准差较大),这说明MICT仅对组内某些个体的FNDC5表达有显著的激活作用。Bonafiglia等(2016)的研究与本研究结果相符,他认为“个体差异”是影响运动激活Irisin信号的重要因素。Dinas在2017年的系统综述中,检索到了16项关于运动激活PGC-1α、FNDC5与Irisin的研究,但其中仅有4项研究显示运动能使PGC-1α与FNDC5同步升高;45项对血液Irisin浓度的研究中,仅有22项显示运动后Irisin浓度上升(Dinas et al.,2017)。Pekkala等(2013)发现,Irisin仅在急性高强度运动后显著增加,长期运动后并无变化。Archundia等(2017)认为,FNDC5与Irisin的激活,不仅依赖与运动强度相关的PGC-1α信号,更多的是与个体差异、机体对急性运动的不适应有关。综上,Irisin信号的激活并不完全与运动强度相关,本研究采用的12周长期HIIT训练并不能激活FNDC5表达。12周长期MICT训练虽然能使一些个体出现FNDC5显著升高的现象,但整体不具显著性,关于MICT是否能够显著激活FNDC5/Irisin信号,其激活作用对何种个体更明显,还需后续的深入研究阐明。本研究同时也对FNDC5与UCP-1表达进行了相关分析,发现无论内脏或皮下脂肪均无显著相关(图6C,6D),提示本研究中HIIT促小鼠内脏脂肪棕色化与骨骼肌分泌的FNDC5/Irisin信号无关。
3.4 HIIT通过交感神经途径促进内脏WAT棕色化
交感神经活动在机体调控脂肪分解的过程中占据核心地位(Braun et al.,2018),交感神经节后纤维直接支配皮下、腹腔及胸部脂肪垫,并通过突触分泌的去甲肾上腺素,促进脂肪分解(Vaughan et al.,2014)。Hillebrand等(2014)对868名成年人的调查发现,人体的脂肪重量,特别是内脏脂肪重量,与交感神活动的高低有关。Sipe等(2017)发现,食物热量摄入限制导致的小鼠脂肪重量减轻,伴随着交感神经活动的增强,同时,交感神经会优先动员内脏脂肪垫分解。这些研究提示,HIIT更显著减少内脏脂肪的作用也可能是通过支配脂肪的交感神经实现的。下丘脑调节机体的摄食、饮水、性行为、打斗、体温等生理机能,同时也是交感与副交感神经的最高位中枢。生理学者在多年前就已知道下丘脑通过交感神经活动直接调节BAT产热(Morrison et al.,2014)。在WAT能够棕色化生成米色脂肪的现象被发现后,Dodd等(2015)发表在Cell上的研究直接证明了下丘脑POMC神经元可通过交感神经活动直接促进米色脂肪的增加,在理论上证实了“下丘脑-脂肪交感神经”轴对WAT棕色化具有明显的作用效果。
TH是合成儿茶酚胺类神经递质的关键限速酶,仅在中枢与周围神经系统及肾上腺髓质内表达。由于交感神经末梢的神经递质主要是去甲肾上腺素,所以TH一直被当作交感神经外周组织调控的活性标志物。TH的短期调节通过丝氨酸位点磷酸化实现,而长期调节则在转录及蛋白表达层面上完成(Nagatsu,2007)。由于本研究的训练方案为12周长期干预,故选取TH表达代表交感神经调控脂肪组织分解的活动程度。无论内脏或皮下脂肪,HIIT组TH表达均高于MICT,说明HIIT组的脂肪交感神经活动相比MICT更加剧烈。需要注意的是,HFD组虽无运动干预,但内脏脂肪TH表达相比MICT组也显著升高,推测存在2种可能:1)由于饲养与训练温度控制在20℃~22℃,而小鼠的体温调节功能较差,此温度激活了HFD小鼠的脂肪交感神经活动。Cui等(2016)的研究有类似的发现,同样的高脂膳食饲养小鼠,22℃饲养个体的脂肪TH表达显著高于30℃饲养的个体。2)由于MICT改善了小鼠体质,MICT小鼠的体温调节相对HFD小鼠更强,对脂肪非颤栗性产热的依赖较少,所以脂肪交感神经活动降低。虽然HIIT同样能够改善小鼠体质,但由于其对植物神经的刺激远大于MICT,所以脂肪TH表达依然较高。由于本研究未对参与体温调节的其他组织器官进行观察,故推论二还需更多实验证据验证。
对UCP-1与TH表达的相关分析显示,皮下脂肪内两者不具显著相关,但内脏脂肪UCP-1与TH呈中度相关(图6A,R=0.578,P<0.05),说明HIIT应促进更多内脏WAT棕色化的过程中,脂肪交感神经对脂肪细胞的调控可能具备潜在的重要作用。由于相关分析并不能验证上游信号(交感神经活动)与下游现象(内脏WAT棕色化)之间存在直接的因果关系,故研究二的内容通过对小鼠进行10天HIIT训练,并阻断内脏脂肪的交感神经调节活动,观察WAT的UCP-1表达是否会因为失去交感神经的调控活动而下降。交感神经对组织的调控,主要末梢分泌去甲肾上腺素,并激活下游细胞的β型肾上腺素能受体来实现。与其他组织不同,脂肪组织的肾上腺素能受体亚型为β3型,故腹腔注射β3型受体的特异性拮抗剂SR59230a,能特异性抑制内脏WAT的交感神经支配(Sipe et al.,2017)。Dewal等(2019)综述了运动促进WAT棕色化的效果,指出10~60天的运动周期均可显著提升UCP-1表达,且UCP-1在运动开始前3周的幅度最明显。而本研究显示,经过10天HIIT后,SR组相比Saline组内脏WAT的UCP-1表达显著降低,说明交感神经末梢的β3受体被阻断后,HIIT促进UCP-1表达的效果被抑制。此结果可有力证明HIIT通过交感神经途径促进了内脏脂肪细胞UCP-1表达,提高了内脏脂肪非颤栗性产热,有利于特异性的减少内脏脂肪。
总之,本研究的相关结果,在分子层面上确认了HIIT能够显著提高内脏WAT的棕色化,且此过程是依靠交感神经途径实现的(图8)。应指出的是,骨骼肌FNDC5与皮下脂肪UCP-1表达间具有中度相关的趋势,但由于目前Irisin作用于脂肪细胞的受体尚不明确(Dinas et al.,2017),很难阻断Irisin对皮下脂肪细胞的调控,故骨骼肌FNDC5/Irisin信号促皮下脂肪的作用,有待进一步研究。

图8 HIIT与MICT促进内脏与皮下WAT棕色化的信号途径
4 结论
相比中等强度持续训练(MICT),高强度间歇训练(HIIT)能够更显著地促进内脏白色脂肪组织棕色化。HIIT促进内脏脂肪组织棕色化与交感神经纤维对脂肪细胞的调控活动密切相关。
猜你喜欢
科普童话·学霸日记(2021年6期)2021-09-05
保健与生活(2021年16期)2021-08-16
恋爱婚姻家庭·养生版(2018年6期)2018-08-01
科学之谜(2018年5期)2018-07-26
家庭百事通·健康一点通(2018年5期)2018-06-14
科学中国人(2016年9期)2016-11-04
米娜·女性大世界(2016年8期)2016-08-17
健康博览(2015年3期)2015-07-03
秀·媛尚(2013年9期)2013-11-06
数学大王·低年级(2009年10期)2009-12-21
