
炎症相关细胞因子及趋化因子在小鼠自身免疫性肝损伤不同时期的表达及作用
2020-03-31 03:14雒真龙张志成
医学研究杂志 2020年2期
艾 国 雒真龙 张志成 王 鸣
细胞因子的合成和代谢,相互之间的拮抗或协同效益等在免疫和炎性反应中扮演重要角色,并与肝脏病理损伤密切相关[1,2]。促炎性细胞因子和抗炎性细胞因子在肝脏中的过度表达可造成炎症过程,是肝脏炎症疾病的驱动力量之一,而具有募集免疫细胞及细胞因子作用的趋化因子亦可能参与其中。近年来的研究已经阐明一些重要的细胞因子在慢性肝脏疾病(如慢乙肝、脂肪肝、酒精肝、原发性胆汁性肝硬化等)中的作用,也有研究者关注了部分细胞因子在自身免疫性肝炎(autoimmune hepatitis,AIH)发病中的作用,但是目前对自身免疫性肝炎的不同状态下和肝脏内环境中促炎性、抗炎性以及趋化因子这3类细胞因子的水平变化和相互关系的研究尚不多见[3~6]。为此,笔者选取并检测了促炎性细胞因子IL-6 和TNF-α,抗炎性细胞因子IL-10,以及趋化因子CXCL10在实验性自身免疫性肝炎(experimental autoimmune hepatitis,EAH)小鼠发病期和缓解期中的表达水平差异和相互关系,探讨自身免疫性肝损伤的机制,以期为自身免疫性肝炎的早期诊断和及时治疗提供理论依据。
材料与方法
1.动物:雌性SPF级C57BL/6小鼠40只,4~6周龄,体质量20~22g,购买并饲养于华中科技大学同济医学院实验动物中心,供给无菌饮水及清洁饲料,适应环境温度及湿度。
2.主要试剂:PCR扩增引物(武汉大风生物技术有限公司设计合成);RT-PCR试剂盒(日本东洋纺公司);IL-6、TNF-α、 IL-10和CXCL10 ELISA试剂盒(达科为生物技术有限公司);弗氏完全佐剂(美国Sigma公司);组织蛋白抽提剂(武汉赛维尔生物有限公司)。
3.动物模型建立及分组:将小鼠随机分为模型组和正常对照组,每组各20只;按照造模型方法建立EAH模型[7]。造模后第2周及6周,每组取6只小鼠,乙醚吸入麻醉后,眼眶动脉采取外周血,静置离心后取血清,检测转氨酶ALT、AST水平;小鼠处死后,取肝组织一部分予以4%甲醛固定,行石蜡包埋切片和HE染色,然后光镜下阅片观察肝组织的病理改变情况;一部分快速冻存于液氮中以提取总RNA和组织蛋白。
4.real-time PCR检测肝组织炎性相关细胞因子和趋化因子的mRNA表达水平:采用TRIzol试剂等提取肝组织总RNA,定量反转录合成cDNA,然后用SYBR GreenⅠ荧光染料技术在25μl PCR反应体系进行PCR扩增。引物设计:内参:GADPH:5′-CAGTGCCAGCCTCTGCTCAT-3′(上游引物),5′-ATACTCAGCACCAGCACAT-3′(下游引物);IL-6:5′-GACAAAGCCAGAGTCCTTCAGAGAGATACAG-3′(上游引物),5′-TTGGATGGTCTTGGTCCTTAGCCAC-3′(下游引物);TNF-α:5′-CCACCACGCTCTTCTGTCTACTG-3′(上游引物),5′-GCCATAGAACTGATGAGAGG-3′(下游引物);IL-10:5′-AACATACTGCTAACCGACTC-3′(上游引物), 5′-ATGCTCCTTGATTTCTGG-3′(下游引物);CXCL10:5′-CCAAGTGCTGCCGTCATTTTC-3′(上游引物),5′-GGCTCGCAGGGATGATTTCAA-3′(下游引物)。real-time PCR反应条件:95℃ 1min,95℃ 15s,58℃ 20s,72℃ 40s,主循环40次,采用2-△△CT法,通过所获循环阈值(Ct)计算目的基因相对表达水平。
5.ELISA法检测肝组织炎性相关细胞因子和趋化因子蛋白表达水平:称取50mg肝组织加入400μl含有PMSF的组织蛋白抽提液中,电动匀浆后,2000r/min离心10min,提取上层清液即组织蛋白液,用ELISA试剂盒依照说明书步骤检测肝组织匀浆中IL-6、TNF-α、IL-10和CXCL10的含量,酶标仪450nm检测待测样本,绘制出标准曲线,得到线性回归方程,然后将样本的吸光度值代入回归方程,得到浓度值。
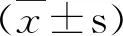
结 果
1.小鼠肝脏病理学改变:EAH组小鼠在建模后2周即发病期,肝脏HE染色显示较多淋巴细胞在血管周围和汇管区等部位浸润,肝细胞部分出现局灶性坏死,炎性反应较明显(图1B);而在缓解恢复期(6周),炎性细胞的浸润以及肝细胞的坏死程度较2周减轻(图1C)。正常小鼠肝脏HE染色则如图1A所示。
2.小鼠血清转氨酶水平改变:EAH小鼠的ALT和AST水平在2周时较正常对照组(0周)均明显升高,差异有统计学意义(P<0.01),两者的升高与肝脏病理改变程度相一致。在6周时的ALT 水平较正常对照组略高,差异无统计学意义;AST 水平则仍较正常对照组偏高,差异有统计学意义(P<0.05,图2)。
图1 小鼠肝脏病理改变(HE染色,×400)A.正常对照组;B.EAH组2周;C.EAH组6周

图2 各组小鼠血清ALT、AST的生化检测结果*P<0.05,**P<0.01
3.小鼠肝脏炎症相关细胞因子及趋化因子mRNA水平变化:通过real-time PCR检测观察到, EAH小鼠肝脏组织IL-6(P=0.035)、TNF-α(P=0.047)、IL-10(P=0.032)和CXCL10(P=0.028)在2周时mRNA表达水平较正常对照组明显增高,在6周时的IL-6(P=0.047)和IL-10(P=0.016)的mRNA表达水平仍稍高于正常对照组,差异有统计学意义;TNF-α(P=0.082)和CXCL10(P=0.061)的mRNA表达水平略高于正常对照组,差异无统计学意义。2周与6周之间比较,EAH小鼠肝组织IL-6 (P=0.080)、TNF-α (P=0.082)、IL-10(P=0.058)的mRNA表达水平下降但差异无统计学意义。CXCL10 mRNA水平下降(P=0.047),差异有统计学意义(图3)。

图3 小鼠肝脏炎症相关细胞因子及趋化因子mRNA水平变化*P<0.05
4.小鼠肝脏炎症相关细胞因子及趋化因子蛋白水平变化:通过ELISA法检测观察到,EAH小鼠肝脏组织IL-6(P=0.004)、TNF-α(P=0.038)、IL-10(P=0.043)和CXCL10(P=0.035) 在2周时蛋白表达水平较正常对照组明显增高,在6周时的IL-6(P=0.006)、TNF-α(P=0.015)和IL-10(P=0.045)的蛋白表达水平仍高于正常对照组,差异有统计学意义;CXCL10(P=0.560)的蛋白表达水平略高于正常对照组,差异无统计学意义。2周与6周之间比较,EAH小鼠肝组织IL-6 (P=0.015)和CXCL10 (P=0.040)的蛋白表达水平下降,差异有统计学意义;TNF-α(P=0.137)、IL-10(P=0.180) 的蛋白表达水平下降,但差异无统计学意义(图4)。

图4 小鼠肝脏炎症相关细胞因子及趋化因子蛋白水平变化*P<0.05,**P<0.01
5.相关性分析:Pearson相关分析显示,EAH小鼠肝组织匀浆中CXCL10与IL-6、TNF-α、IL-10均呈正相关(r分别为0.8214、0.8910、0.8504,P均<0.05);IL-10与 IL-6、TNF-α均呈正相关(r分别为 0.9644、0.8897,P均<0.05)。
讨 论
自身免疫性肝炎是异常的自身免疫反应介导的肝细胞损伤和肝实质炎性病变,目前在肝病方面关于相关细胞因子的研究结果尚存在一定差异,尤其在自身免疫性肝炎的细致变化仍具有一定研究空间。结合既往文献,本研究选取部分细胞因子并就其在EAH中的表达变化进行了探索分析。
研究结果显示,在EAH发病期,肝脏内的IL-6在mRNA和蛋白表达水平均明显升高,同时伴随血清转氨酶升高和肝脏炎性细胞浸润增加。IL-6作为一种重要的炎性细胞因子,在宿主防御、调控B细胞、活化自然杀伤细胞(natural killer cell,NK)、参与炎性反应进程中均发挥重要作用[8]。尽管其表达调控遵循转录机制,一旦转录后机制失调,持续产生的IL-6却对慢性炎症及自身免疫性疾病具有病理作用[9]。IL-6可通过gp130 激活JAK/STAT 信号通路介导炎性反应[10]。IL-6/STAT3信号通路可招募并活化中性粒细胞(PMN),经过趋化、黏附,渗出血管壁浸润至肝实质内的PMN活化,可引起肝细胞的损伤[11]。已有临床研究表明,慢性肝脏疾病、肝硬化患者血清IL-6水平的持续升高,对肝脏组织造成不良后果[6]。本研究显示,IL-6的表达在缓解期虽有下降仍高于正常水平,这也说明持续的IL-6表达可作用于整个自身免疫性炎症发生、发展的过程。
与IL-6的表达变化类似,笔者发现促炎细胞因子TNF-α亦表现为发病期表达水平均升高,在缓解期随着血清转氨酶水平下降和肝内炎性细胞浸润减少,TNF-α的表达水平虽较发病期有所下降,但基本仍然高于正常水平。TNF-α主要是由巨噬细胞和单核细胞分泌,可触发TNF-α和IL-6本身的级联反应。TNF-α在机体的炎性反应和免疫调控中具有较为重要的功能,即活化巨噬细胞、增加细胞毒性、抑制B细胞的激活及抗体的产生、调节HLAⅡ类分子和黏附分子的表达,以及促进迟发型超敏反应等。TNF-α的这些生物学效应能够加重机体自身免疫反应,从而令自身免疫性疾病进展加速[12]。在关于Th17细胞及其细胞因子与AIH相关性的研究中发现,Th17细胞的数量及分泌的细胞因子如IL-6和TNF-α等增加与肝脏炎症和纤维化严重程度有关[13]。这也与笔者在EAH模型中观察到的结果基本相符。结合本研究观察结果和相关文献报道,笔者推测IL-6及TNF-α作用于整个自身免疫性炎症发生、发展的过程,可作为炎性反应的敏感指标,亦是疾病进展的重要影响因素。
IL-10是具有抑制细胞免疫反应、促进体液免疫反应和抗炎功能的细胞因子,在细胞因子网络调节中发挥重要作用[14]。IL-10可抑制单核细胞生成TNF-α、IL-1、IL-6、IFN-γ等细胞因子,刺激单核细胞产生IL-1受体拮抗剂,发挥抗炎功能[15]。本研究显示,EAH小鼠肝脏IL-10的表达水平从发病期至缓解期均高于正常,且伴随IL-6及TNF-α变化而变化,相关性分析显示IL-10与 IL-6、TNF-α均呈正相关,而并非负相关,推测IL-10表达水平的这种变化可能为机体维持免疫平衡的代偿性改变。以上几种促炎性细胞因子与抗炎性细胞因子的表达变化和关系如上所述,那么跟两者密切相关的趋化因子的表达状况又会是如何?
趋化因子家族在肝脏微环境中关于细胞之间的信号调节方面的研究已逐渐受到重视[16]。其中CXCL家族可在某些自身免疫性疾病中发挥明显的促炎效应,如原发性胆汁性肝硬化患者外周血和肝组织中CXCL9和CXCL10表达水平显著增加,且随疾病进展而升高[17]。而在本研究中EAH小鼠肝脏内CXCL10表达水平表现为在发病期增高,缓解期则下降。趋化因子本身能够与血管内皮细胞相结合,具有激活并趋化白细胞移动的作用[18]。趋化因子信号系统对于效应细胞和调节细胞的募集有关键作用,参与肝脏免疫性疾病的发生[19]。本研究中EAH小鼠肝脏内CXCL10表达水平与IL-6、TNF-α、IL-10均呈正相关,表明CXCL10在自身免疫性肝炎中可能既募集表达促炎性细胞因子又能募集表达抗炎性细胞因子的免疫细胞,参与肝脏炎症活动,某种程度上可能具有双向调节的作用。
综上所述,促炎性细胞因子、抗炎性细胞因子及趋化因子等细胞因子形成错综复杂的细胞因子调节网络,作用于肝脏并影响肝脏疾病发生、发展;理解这些细胞因子在疾病发生、发展过程中的变化和作用,对于阐释AIH的肝损伤机制具有重要意义;针对细胞因子的分子靶向治疗也为肝脏疾病提供了新的诊疗思路和干预手段。
猜你喜欢
生物化工(2022年4期)2022-09-20
现代临床医学(2022年1期)2022-02-12
现代临床医学(2021年4期)2021-07-31
昆明医科大学学报(2021年4期)2021-07-23
天津医科大学学报(2021年2期)2021-03-29
世界最新医学信息文摘(2021年55期)2021-01-07
中西医结合心脑血管病杂志(2016年20期)2016-03-01
中国医药生物技术(2015年4期)2015-12-26
中华胰腺病杂志(2015年5期)2015-12-08
中国全科医学(2015年33期)2015-12-02
