
荒漠草原建群种及其枯落物的C、N、P生态化学计量特征1)
2020-03-26 11:34万芳蒙仲举党晓宏
东北林业大学学报 2020年2期
万芳 蒙仲举 党晓宏
(内蒙古农业大学,呼和浩特,010018)
生态化学计量学是结合生物学和化学的基本原理,研究C、N和P等元素平衡和系统能量循环的理论[1-3],统一了分子、细胞、有机体、种群、生态系统和全球尺度等生物学科层次[4],为植被养分利用提供新的思路和研究手段[5]。C、N和P是植物生长所必需的元素[6-7],共同参与植物的生理代谢过程[8],其中C是植物生命的骨架元素[8],是通过光合作用合成的,N和P是植物生长的限制元素,成为制约生产力的主要因素[9]。其N/P的大小可以反应植物生长过程中受到哪种元素的限制,为研究植物中元素在其生长过程中发挥的作用提供有效的方法。
枯落物是生态系统养分的储存库,是植物与土壤养分进行转移的一个重要枢纽[10],是土壤养分的的来源之一[11]。土壤养分的积累主要是枯落物分解方式的归还[12]。植物在枯落前会将养分进行转移,实现养分的再利用(养分再吸收),增加了养分在植物体内的存留时间[13],同时也为植物的生长提供了养分[14]。养分再吸收作用是植物适应贫瘠环境、提高物种间竞争的重要途径[15],也是植物增强竞争力、提高养分吸收能力和生产力的重要策略之一[16]。因此,以植物和枯落物元素计量学特征为指标,探讨植物生长中主要的限制元素和养分再吸收效率具有一定的意义。
目前,国内外学者对化学计量比展开了大量的研究,大部分的研究集中在森林[17]、湿地[18]、草地[19]等生态系的叶片和枯落物C、N、P生态化学计量特征,都是对单一组分(叶或者枯落物)或者两个组分组合(植物叶片和枯落物)的C、N、P进行生态化学计量研究[20],对典型荒漠草原植物与其枯落物研究较少。希拉穆仁草原是典型的荒漠草原,是内蒙古草原的重要组成部分,属于草原向荒漠过渡的草原类型,荒漠草原除气候干旱,水资源匮乏,土壤贫瘠养分含量低也是荒漠草原的一大特点,在这种极端的环境下养分限制并不明确。基于此,本研究以希拉穆仁草原建群种(羊草、短花针茅和芨芨草)及其枯落物为对象,运用生态化学计量学方法,分析3种植物养分限制格局及养分再吸收作用,为希拉穆仁草原植被的恢复和草地管理提供科学参考。
1 研究区概况
研究区位于内蒙古达茂旗希拉穆仁镇希拉穆仁草原(42°20′~42°40′N,109°16′~111°25′E),该区属于温带大陆性季风气候,年平均降雨量约为280 mm,主要集中在7、8月份,年平均气温3.4 ℃,蒸发量大,年均蒸发量2 227 mm;地带性土壤为粟钙土,质地粗糙,土壤养分含量较低。植被以旱生和中旱生植物为主,植物耐旱、耐贫瘠、适应性强、根系较发达;4月返青,返青后植物生长较快;建群种有羊草(Leymuschinensis),草地群落以羊草(Leymuschinensis)+短花针茅(StipabrevifloraGriseb)为主,芨芨草(AchnatherumsplendensNevski)、银灰旋花(ConvolvulusammanniiDesr)、冷蒿(Artemisiafrigida)、赖草(Leymussecalinus)等分布也较为广泛,植被类型为典型荒漠草原特征。由于旅游业的发展和过度放牧等人为干扰,使得草原退化严重。试验地位于水利部牧区水利科学研究所实验基地的围封区(围封年限16 a,围封面积133 hm2)。
2 材料与方法
2.1 采样地设置与样品采集
2018年8月中旬在希拉穆仁草原的围封区选取地势平坦、土壤类型基本相似、群落分布一致的区域,采用样线与样方结合的方法,在选好的区域内以一个点为中心,东西方向和南北方向各布设一条100 m样线,两条样线交叉呈十字。将选择的区域分成4个小区域,在每个区域内布设4个1 m×1 m的样方,且4个样方呈正方形布设(样方距离样线10 m、样方间距为10 m),总共布设16个样方,采集羊草、短花针茅、芨芨草植物叶片与其枯落物。此时的植物处于生长旺盛期,植物叶片与枯落物并存,在所有的样方中采集每种植物叶片及其枯落物大约100 g(采集干燥且枯黄的枯落叶片),混合均匀,对所有采集回来的植物叶片与枯落物要进行预处理去除植物表面的杂质,将处理好的植物叶片和枯落物分别装进信封袋中放入烘箱中,先在105 ℃下杀青30 min,然后在65 ℃烘至恒质量。
2.2 样品测定
将从野外采集回来的植物叶片和枯落物用粉碎机粉碎,并且过0.15 mm的筛,磨成粉末后测定其C、N、P的质量分数。植物叶片与枯落物的C采用重铬酸钾-外加热法测定;N、P采用H2SO4-H2O2消煮,将消煮液分别用凯氏法测定N,钒钼黄比色法测定P[21]。养分再吸收效率指的是植物正常生长的养分与枯落植物的差值,即植物枯落前养分转移的质量分数[20]。养分再吸收效率的计算公式为:
式中:Re为养分再吸收效率;TH为植物正常生长的养分质量分数(g/kg);TK为枯落物中养分的质量分数(g/kg)。
2.3 数据处理
应用Excel 2013和SPSS 23.0统计分析软件对测定数据进行整理和分析。所有统计数据以平均值及标准误差表示,对不同植物叶片和枯落物叶片的养分质量分数及化学计量比进行单因素方差分析(One-Way ANOVA)以及多重比较(LSD),用CANOCO 5.0软件对不同植物叶片和枯落物C、N、P质量分数与化学计量比进行关系分析。
3 结果与分析
3.1 不同植物叶片和枯落物C、N、P质量分数及其N、P养分再吸收效率
不同植物叶片C、N、P质量分数变化范围分别为383.59~406.49、23.14~25.80、1.18~1.98 g/kg,均值分别为398.74、24.48、1.55 g/kg,变异系数分别为0.76%、2.88%、13.65%(图1)。从变异系数可以得出不同植物叶片的C、N质量分数变化较小,P质量分数变化较大。不同枯落物的养分质量分数均值分别为388.97、1.79、1.17 g/kg;变异系数分别为0.90%、8.00%、8.93%,可以看出不同枯落物N、P质量分数变化比较明显、C质量分数变化较小。植物枯落后C、N和P的质量分数小于其植物叶片C、N和P质量分数,说明植物在枯落前养分进行了转移,实现了养分的再吸收利用。3种建群种与其枯落物的C、N质量分数均存在差异显著(P<0.05),植物叶片的P质量分数差异不显著(P>0.05),而枯落物的P质量分数在不同植物间差异显著(P<0.05)。

表1 3种植物的叶片与枯落物碳氮磷质量分数
注:表中数值为平均值±标准差;同行不同大写字母表示同种植物和枯落物质量分数差异显(P<0.05),同列不同小写字母表示不同植物间或枯落物质量分数差异显(P<0.05)。
养分再吸收效率的大小表示养分再吸收的能力。N的再吸收效率在13.51%~77.90%,P的再吸收效率在2.65%~49.51%。其中芨芨草的N、P的回流率均大于羊草与短花针茅,说明芨芨草在枯萎时对N、P的利用效率高(表2)。

表2 3种植物叶片与枯落物的N、P的养分再吸收效率
3.2 不同植物叶片和枯落物的生态化学计量特征
不同植物叶片的N/P、C/N、C/P的值分别在11.81~21.70、15.76~16.60、195.47~273.37之间波动,变异系数分别为12.82%、3.23%、13.95%。不同枯落物的N/P、C/N、C/P的值分别为7.72~17.05、17.71~64.46、215.43~487.46,变异系数为15.50%、2.10%、9.14%(表3)。从化学计量比可以得知植物叶片C/N和C/P都小于枯落物,变异系数C/N较小,说明植物体内C质量分数变化较小,主要取决于N和P质量分数。3种植物叶片的N/P、C/P差异显著(P<0.05),但3种植物叶片的C/N无显著差异(P>0.05);3种枯落物的N/P、C/P差异显著(P<0.05),芨芨草叶片与枯落物的N/P、C/N、C/P差异显著(P<0.05)。
注:表中数值为平均值±标准差;同行不同大写字母表示同种植物和枯落物质量分数差异显(P<0.05),同列不同小写字母表示不同植物间或枯落物质量分数差异显(P<0.05)。
3.3 不同植物叶片和枯落物C、N、P质量分数与生态化学计量学特征关系
植物叶片C和P质量分数与C/N呈正相关,而与N/P和C/P呈负相关;N质量分数与N/P和C/P呈正相关。羊草受C/N的影响较大,短花针茅受C/P影响较大。枯落物C与N/P、C/P和C/N呈正相关;N与N/P呈正相关与C/N和C/P呈负相关,P与N/P呈正相关与N/P和C/P呈负相关。芨芨草枯落受C/N的影响大,短花针茅受N/P的影响较大(图1)。
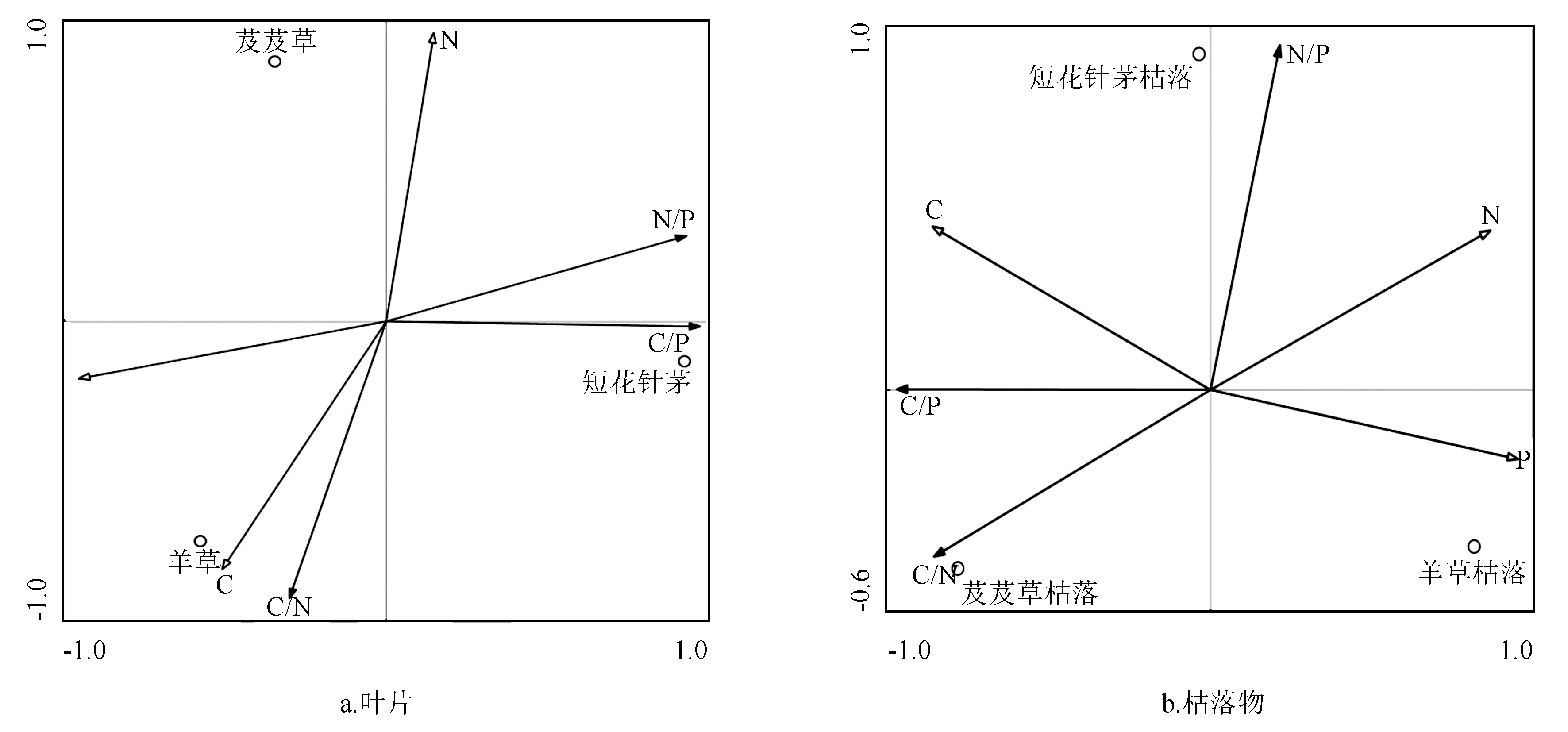
图1 3种植物叶片和枯落物C、N、P质量分数与生态化学计量学特征关系
4 讨论
4.1 不同植物叶片及其枯落物C、N、P质量分数特征与养分再吸收能力
3种植物养分质量分数及对养分的利用效率都存在差异性。一些研究表明植物叶片的C、N和P质量分数均高于枯落物的[22],本研究结果表明3种植物C、N和P质量分数高于其枯落物的,植物在凋落前养分会进行转移。其中植物叶片的平均C质量分数(398.74 g/kg)低于全球492种陆生植物碳平均C质量分数(464 g/kg)[1]546,因为不同植物对环境的适应是不同的,说明该区C质量分数较低,固C能力可能较弱。N、P是植物体内蛋白质、叶绿素、核酸、酶等的主要成分,共同参与植物的生理活动[23]。N质量分数的平均值(24.48 g/kg)高于全球尺度的N平均质量分数(20.6 g/kg)[24],N质量分数高说明植物的光合作用强,植物体内的蛋白含量高。P的平均质量分数(1.55 g/kg)大于全球尺度P的平均质量分数(1.40 g/kg)[25]579。因为荒漠区的植物根系都比较发达,吸收养分的能力强,而根系作为植物运输的通道,养分不会在根系中大量的储存,会把养分运送到植物的各个器官中,因此植物的N和P的质量分数高,同时也可说明植物在干旱的条件下贮存养分的能力强。而枯落物C、N、P平均质量分数分别为407.15、14.79、1.17 g/kg,与全球、国内陆地植物的C(371.1~522.1 g/kg)、N(8.0~16.6 g/kg)、P(0.4~1.3 g/kg)[25-26]相比,N、P的质量分数较高,C质量分数较低,说明植物体内N、P养分的利用效率较高,再次证明植物在极端条件下具有较高的适应能力,养分的重吸收作用象征植物适应贫瘠环境,是植物体内养分保护的一种策略[27]。3种枯落物叶片的N和P质量分数与植物本身的再吸收能力有关[22]。植物在枯落前会发生养分的转移,使养分在植物体内存留,防止养分的流失[22]。本研究结果表明,3种植物的N和P质量分数显著高于枯落物的,说明植物在枯落前养分进行了转移,实现了养分再吸收利用。本研究发现芨芨草N的养分再吸收率角高于高于短花针茅与羊草,说明芨芨草在干旱贫瘠的环境下适应能力强。
4.2 不同植物叶片及其枯落物碳积累与N、P养分格局限制
生态化学计量比是生态系统和功能重要的特征,植物体内的的化学计量比表征了C积累动态以及N和P养分的限制格局[8]。植物的C/N和C/P能够反映同化C的能力,在一定程度上反应植物养分利用率,也可以表示C与N、P之间的互相协调能力,其比值在合理范围内表示C与N、P元素相互平衡和制约,有利于植物生长[28]。3种不同类型的植物叶片的C/N比值(16.32)低于全球水平(22.5),说明C的利用效率低,对C的吸收能力较弱。C/P比值(276.37)高于全球水平(232)[1],说明植物的养分利用效率相对较高,可能与研究区土壤温湿度、气候条件和地形条件有关。而枯落物的C/N(34.01)与C/P(347.09)低于全球枯落物C/N(66.2)、C/P(3144)[29]。枯落物C/N和C/P高于植物叶片C/N和C/P,这是因为植物在凋落之前通过养分再吸收对N、P进行了重吸收作用[30],因此枯落物中C/N和C/P比均高于植物叶片。
生态化学计量学应用的一个重要方面是根据植物的N/P比来判断环境对植物生长的养分供应状况[4],可以明确表示植物受到N、P哪种元素的限制。由于研究区域的不同,植物种的不同以及植物不同生长期N、P质量分数的差异都会影响到N/P临界值的变化[31]。通常将N/P比值为16作为评价植物生长受N或P限制的临界值[22]。N/P比大于16受P的限制,小于14受N的限制,介于14~16则受N和P的共同限制。本实验的研究结果表明N/P平均比值(16.95)高于全球尺度的N/P(13.8),说明植物受P的限制,这与Han等[20]对中国753种陆地植物的研究结果一致,即我国陆地植被大部分受P的限制。枯落物N/P是影响枯落物分解和养分归还速率的重要因素之一[8],3种枯落物的N/P的平均值为(11.66)低于全球45.5[29],枯落物N/P>25时分解主要受P限制[32]。本研究中,枯落物N/P(12.32)比均低于25,说明枯落物的分解主要受N限制。
5 结论
本研究区3种植物叶片的C、N、P质量分数高于其枯落物的的质量分数,且差异显著(P<0.05),表明植物在枯落时会发生养分转移,短花针茅与芨芨草的养分再吸收效率均表现为N大于P,说明对N元素的养分再吸收能力较强,有利于在贫瘠的环境中生存。
3种植物叶片与枯落物的C质量分数变化幅度较小,因此,C/N和C/P的大小主要受N、P质量分数的影响。3种植物N/P均值大于16,表明植物在生长过程中主要受P元素的限制,在后期抚育管理时可以适当地施加P肥,弥补土壤中缺P的现象,有利于植被的恢复。
猜你喜欢
中国土壤与肥料(2022年4期)2022-06-14
第二课堂(课外活动版)(2022年2期)2022-04-23
农业工程学报(2022年1期)2022-03-25
英语文摘(2021年3期)2021-07-22
落叶果树(2021年6期)2021-02-12
植物保护(2020年3期)2020-06-08
文苑(2020年12期)2020-04-13
小星星·阅读100分(低年级)(2019年10期)2019-11-26
安徽文学(2019年6期)2019-06-28
海峡姐妹(2019年1期)2019-03-23
