
生态调控后贵州花溪水库浮游植物群落结构的变化特征及影响因素
2020-03-17 07:13马欣洋李秋华袁振辉李红梅张华俊
环境科学研究 2020年3期
马欣洋, 李秋华*, 陈 倩, 何 应, 袁振辉, 李红梅, 张华俊
1.贵州师范大学, 贵州省山地环境信息系统和生态环境保护重点实验室, 贵州 贵阳 550001 2.贵州中环科技有限公司, 贵州 贵阳 550003
在水生态系统中,浮游植物作为主要的初级生产者,其群落组成和种群变化能直接并快速反映水环境的动态变化[1]. 浮游植物群落的组成种类、数量分布及丰度是评价水库生态系统的关键指标,通常反映水体的富营养化程度[2-5]. 浮游植物生长过程主要受温度、光照、营养盐、水动力等环境因子的影响,而且会随水环境的改变做出响应[6-10]. 浮游植物传统分类方法虽能较好地反映群落结构组成,但在环境生态特征上存在一定的局限性. Reynolds等[11]根据浮游植物生理、形态提出了浮游植物功能群,Padisk等[12]在其基础上进行了补充和完善. 目前,该理论已广泛应用于国内外水生态的研究[13-16]. Devercelli等[17]研究了Paraná河异常水文期浮游植物结构变化过程,发现功能群是重建浮游植物群落分类的一种有效方法. Dokulil等[18]运用功能群研究了欧洲阿尔卑斯山区深水湖泊,得出功能群能够很好地反映出受到人类干扰湖泊水体富营养化的演变过程;董静等[19]探究了3个湖泊浮游植物功能群自1960年以来的演替过程,提出营养盐的增加可能是导致这3个湖泊功能群演替的主要原因. 同时,将浮游植物结构变化动态与生境结合起来,为更合理地揭示生境变化对浮游植物的选择机制、预测浮游植物群落演替提供了有力工具[20].
近年来,湖泊和水库富营养化范围逐渐扩大[21-22],水华出现频率日益增加. 越来越多的学者关注如何控制这一现象的发生. 濮培民等[23]对红枫湖右二湾进行了生态调控,有效改善了其水质状况和富营养化程度. 李秋华等[24]利用物理-生态集成技术有效地改善了百花湖麦西河河口水质状况及其浮游植物的群落结构. 花溪水库是贵阳市重点供水水库之一,属于贵州高原典型喀斯特中型水库,其水质状况对贵阳市尤为重要. 2017年3月发生了水华束丝藻的水华,通过采用无毒高效的生态除藻剂(HP-Ⅰ号)进行控藻和杀藻,同时为防止水华扩散,采取物理隔离工程——垂直帘隔离、藻类调控工程等措施,使水华得到了有效的控制. 该次生态调控为期60 d,但生态调控后该水库浮游植物群落结构变化尚属未知. 该研究对生态调控后花溪水库浮游植物及水质进行了1年的调查研究,分析了其在生态调控后浮游植物的结构变化特征与环境因子的关系,以期为贵州高原水资源保护与治理提供理论依据和技术支撑.
1 材料与方法
1.1 水库概况及采样点设置
花溪水库位于贵阳市花溪区西部,属于亚热带季风气候区,年均气温为14.9 ℃,流域面积为315 km2,设计正常水位高程为 1 100 m,总库容为3.14×105m3. 该研究于2017年3月—2018年3月逐月进行水质和浮游植物采样. 根据《湖泊富营养调查规范》与花溪水库水域特点及生态调控区域设置S1(26°26′25″N、106°35′43″E)、S2(26°27′12″N、106°36′38″E)、S3(26°26′39″N、106°37′26″E)、S4(26°26′32″N、106°38′21″E)、S5(26°26′13″N、106°38′34″E)5个采样点(见图1),每个采样点均取表层(0.5 m)、中层(5 m)、下层(10 m)的水样.

注: 箭头表示水流方向. 图1 花溪水库采样点分布Fig.1 Distribution of Sampling points in Huaxi Reservoir
1.2 样品采集与分析
浮游植物定量样品:每个采样点均取表层、中层和下层水样,混合后每个水样保留1.5 L,用浓度为3%~5%甲醛固定. 放置实验室静置24~48 h后用虹吸法浓缩至30 mL,在显微镜下参考文献[25]鉴定. 浮游植物定性样品:采用25#浮游生物网在水面以下水平及垂直方向呈“∞”字拖网,加入3%~5%甲醛溶液固定. 浮游植物功能群分类参考Reynolds等[11-12]等提出的浮游植物功能群分类方法和文献[25]. 浮游植物的细胞体积根据与其相近几何形状的体积计算,并假定浮游植物的密度为1 gm3,计算浮游植物的生物量[26-27].
用雷磁水质监测仪(DZB-718,上海仪电科学仪器股份有限公司)现场测定WT (水温)、pH和电导率(EC)等,SD (透明度)用塞氏盘(Secchi Disk)进行测定. 按照GB 3838—2002《地表水环境质量标准》测定ρ(TP)、ρ(TN)、ρ(DIN)〔包括ρ(NO3-N)、ρ(NO2-N)、ρ(NH4-N)〕与ρ(PO4-P)等指标.ρ(Chla)采用0.45 μm醋酸纤维滤膜抽滤水样500 mL,反复冻融,用丙酮萃取法测定[28].
1.3 数据处理与分析
采用ArcGIS软件绘图,NMDS(非度量多维尺度分析)运用R语言中vegan绘制,浮游植物生物量与环境因子的相关性以及Pearson分析采用SPSS 19.0软件,采用Canoco 4.5软件分析浮游植物功能群特征与环境因子的关系,分析过程前对物种数据以及环境变量(pH除外)进行log2(x+1)转换. 其他图采用Origin 9.0软件绘制.
综合营养状态指数[17]计算公式:
式中,TLI(∑)为综合营养状态指数,TLI(j)为第j种参数的营养状态指数,Wj为第j种参数的营养状态指数的相关权重,m为评价参数的个数.
2 结果与分析
2.1 生态调控前后环境因子的变化特征
生态调控前后花溪水库环境因子变化如图2所示. 由图2可见:与生态调控前花溪水库营养盐浓度相比,生态调控后有所降低,但也呈较强的季节性变化;ρ(TP)季节性波动较大,范围为0.01~0.19 mg/L,方差分析表明,各月之间存在显著差异(P<0.05);生态调控前,ρ(TN)平均值约为5.94 mg/L,生态调控后,ρ(TN)平均值约为2.29 mg/L,且在S2采样点平均值高达3.22 mg/L,方差分析表明,2017年3月、7月与其他月份存在显著差异(P<0.05);WT(水温)年均值为18 ℃,最高值为26 ℃,变化范围较大,各月之间存在显著差异(P<0.05);ρ(CODMn)变化不大,年均值约为2.00 mg/L,最高不超过6.00 mg/L,生态调控前后各月存在显著差异(P<0.05);生态调控前,ρ(Chla)平均值为54.14 mg/m3,最大值出现在S5采样点(81.60 mg/m3),生态调控后,最小值出现在11月的S2采样点(1.53 mg/m3).
2.2 浮游植物群落的变化特征
2.2.1浮游植物种类组成
花溪水库生态调控前的一次采样中5个采样点共鉴定出浮游植物4门18种,其中硅藻门4种、绿藻门9种、蓝藻门2种、甲藻门3种. 绿藻门种类最多,约占50.0%;硅藻门和甲藻门次之,分别约占22.2%和16.7%;因发生水华束丝藻水华,蓝藻门物种较为单一,约占11.1%. 生态调控后13次采样中5个采样点共鉴定出浮游植物6门66种,其中硅藻门23种、绿藻门25种、蓝藻门9种、甲藻门5种、裸藻门3种、隐藻门1种. 绿藻门占比最大,约占37.9%;硅藻门和蓝藻门次之,分别约占34.9%和13.6%;甲藻门相对较少,约占7.6%;裸藻门和隐藻门均较少,二者之和约占6%. 生态调控前,花溪水库浮游植物生物量中蓝藻占绝对优势(95.0%),生态调控后硅藻占主要优势(50.2%),甲藻和绿藻次之(分别为33.1%和12.5%),可见生态调控后花溪水库浮游植物群落结构发生了较大的变化(见表1).
根据Reynolds等[11-12]提出的浮游植物功能群分类方法,在研究期间,花溪水库生态调控前共归类出10个功能类群,分别为D、P、B、J、X1、F、X2、H1、S1、Lo. 生态调控后共归类出20个功能类群,分别为D、P、B、J、X1、F、X2、H1、S1、Lo、MP、C、G、TC、X3、N、W1、T、W2、M(见表2).
2.2.2生态调控前后浮游植物功能群及生物量时空分布
将相对生物量大于5%的功能群定义为该点的优势功能群,大于50%为该点绝对优势功能群[29]. 由图3可见,生态调控前浮游植物生物量峰值为 7 796.8 μg/L,代表藻种为水华束丝藻,其主要由功能群H1贡献. 生态调控后浮游植物生物量整体有所降低,变化范围为263.7~5 233.1 μg/L,由于生物量较大的小环藻和多甲藻大量的生长,最高值出现在2017年9月;其次是2017年10月、11月和2018年1月,生物量分别为 4 188.9、3 982.5 和 3 603.6 μg/L;最低值出现在2018年2月. 生态调控前功能群H1占绝对优势(达95%),生态调控后优势功能群由H1转变为Lo/B(2017年4月),2017年5月功能群B开始大量出现,占比高达90%,且之后一直为优势功能群,占比为25.4%~63.1%;2017年4月、7月、9月和2018年3月功能群Lo所占比例较大,达33.3%~68%;2017年6月、8月和10月功能群N也占一定优势,分别为12.6%、13.3%和33%;除优势功能群B外,2018年1月和2月功能群P/W2和D/P/W1也占一定比例.
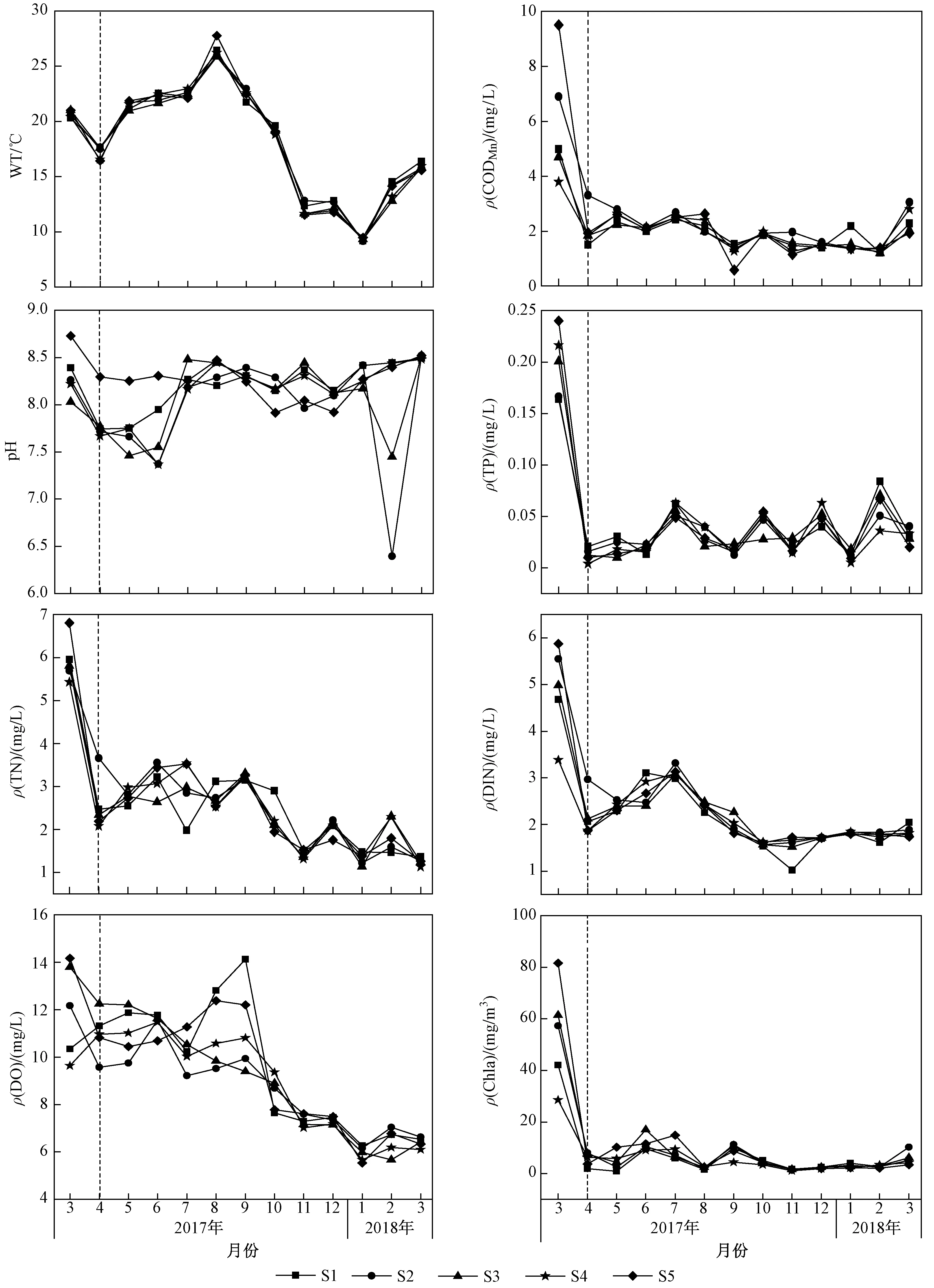
注: 虚线左边为生态调控前;虚线右边为生态调控后. 图2 生态调控前后花溪水库环境因子变化Fig.2 Changes of environmental factors before and after ecological regulation of Huaxi Reservoir
2.2.3生态调控前后浮游植物群落结构变化特征
基于Bray-Curtis矩阵相似性系数,采用NMDS对花溪水库浮游植物群落结构进行排序. 在二维空间上应力值为0.141,小于0.2,表明排序效果比较理想. 由图4可见,生态调控前后花溪水库浮游植物群落结构存在明显差异.生态调控前主要以蓝藻门为主,对应功能群H1. 生态调控后主要以硅藻门和甲藻门为主,对应功能群B和Lo;2017年7月和8月绿藻门也占一定优势,其对应功能群N和X2;2018年1月和2月裸藻门开始出现,其对应功能群W1和W2;2017年3月H1为优势功能群,而2018年3月优势功能群主要由B/Lo组成,浮游植物群落结构发

表1 生态调控前后花溪水库浮游植物组成对比
生了明显变化. W1和W2所占生物量相对较小,在生物量排序图中不能够明显显示,但在NMDS结果中可以很清晰地反映花溪水库浮游植物功能群的分布情况.
2.3 生态调控前后浮游植物功能群与环境因子关系
2.3.1RDA分析
通过对生态调控前后花溪水库浮游植物优势功能群生物量进行DCA分析,发现生态调控前后的排序轴梯度长度最大值均小于3,因此选择线性模型. 根据生态调控前后浮游植物功能群出现频率及相对生物量,选取了D、P、B、J、X1、F、H1、Lo等15个功能群和ρ(TN)、ρ(DIN)、ρ(TP)、WT、EC、ρ(DO)等12个环境因子进行RDA分析. 采用蒙特卡拟合方法分别对生态调控前后的浮游植物功能群生物量和环境因子关系进行显著性检验,结果(见图5)显示,生态调控前显著解释性的环境因子为ρ(DIN)、SD、ρ(CODMn)(P<0.05);生态调控后显著解释性的环境因子为ρ(DIN)、WT、ρ(DO)(P<0.05). 生态调控前轴1、轴2的特征值分别为0.735和0.164,生态调控后轴1、轴2的特征值分别为0.406和0.216,表明轴1、轴2能够解释绝大部分浮游植物的分布情况.

表2 浮游植物功能群的代表性藻种及适宜生境
2.3.2Pearson相关性分析
由表3可见:生态调控前功能群H1与ρ(TN)、ρ(DIN)呈显著正相关,与ρ(DO)、SD呈显著负相关;功能群D与pH呈显著负相关;功能群B与ρ(CODMn)呈显著正相关,与其他环境因子无显著相关性. 生态调控后功能群B与ρ(CODMn)呈显著负相关;功能群P与ρ(PO4-P)呈极显著相关,与ρ(TN)、WT、ρ(DO)呈显著负相关;功能群S1与ρ(PO4-P)、SD呈极显著正相关,与WT呈显著负相关;功能群D与pH呈正相关,与其他环境因子无显著相关性. 结果表明,生态调控前后优势功能群与环境因子的相关性发生了改变,也从侧面反映出生态调控前后花溪水库浮游植物功能群结构的变化.
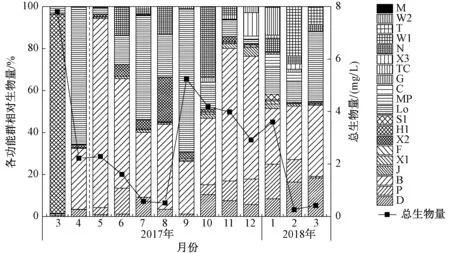
图3 生态调控前后花溪水库浮游植物各功能群相对生物量和总生物量的变化Fig.3 Relative biomass and biomass of algae functional groups in Huaxi Reservoir of the before and after ecological regulation
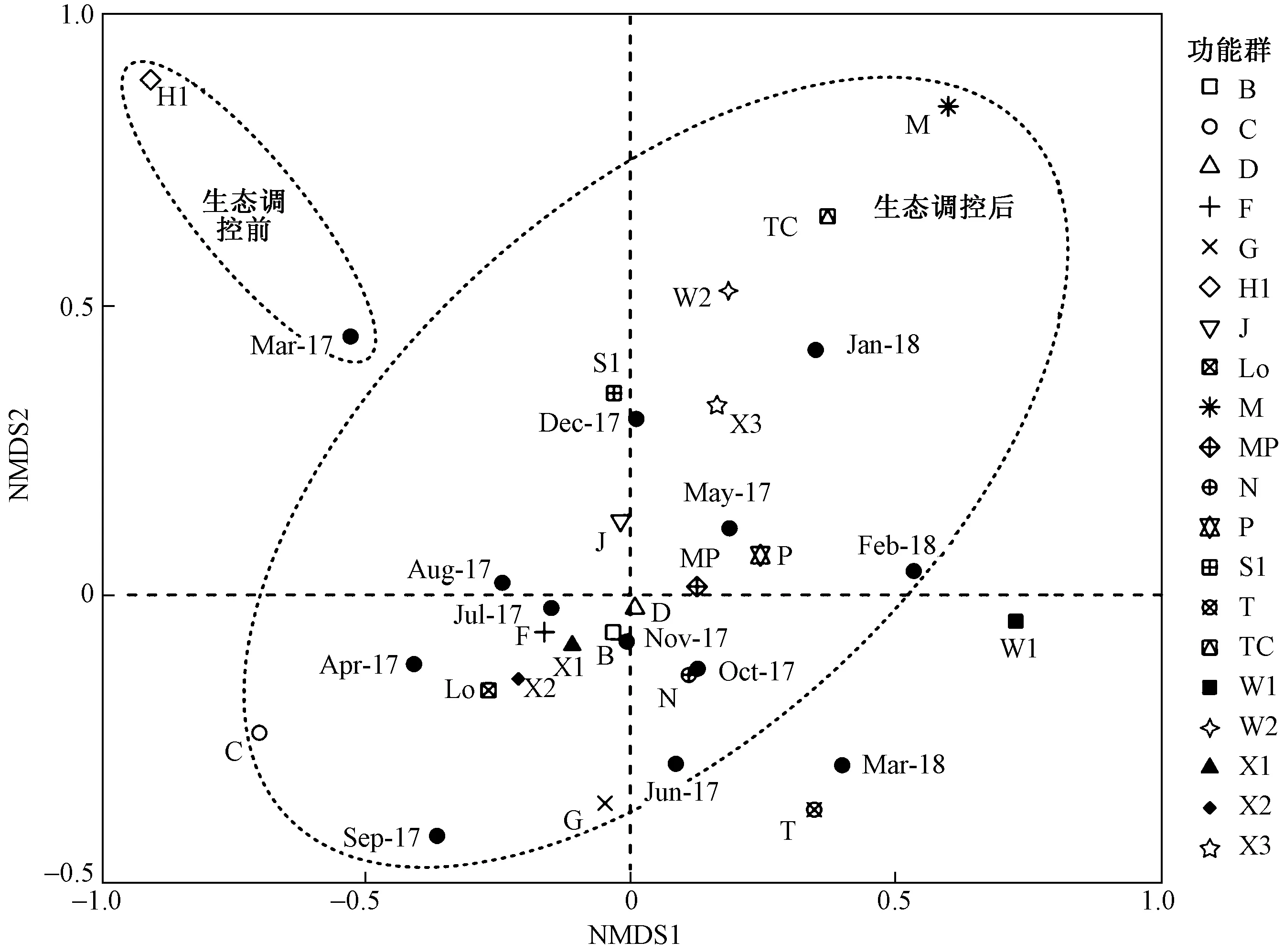
注: Mar-17表示2017年3月,Mar-18表示2018年3月,依次类推. 图4 花溪水库浮游植物功能群NMDS结果Fig.4 NMDS analysis of phytoplankton functional groups in Huaxi Reservoir

图5 生态调控前后花溪水库浮游植物功能群与环境因子RDA结果Fig.5 Redundancy analyancy of phytoplankton functional groups related to environmental factors of the before and after ecological regulation in Huaxi Reservoir
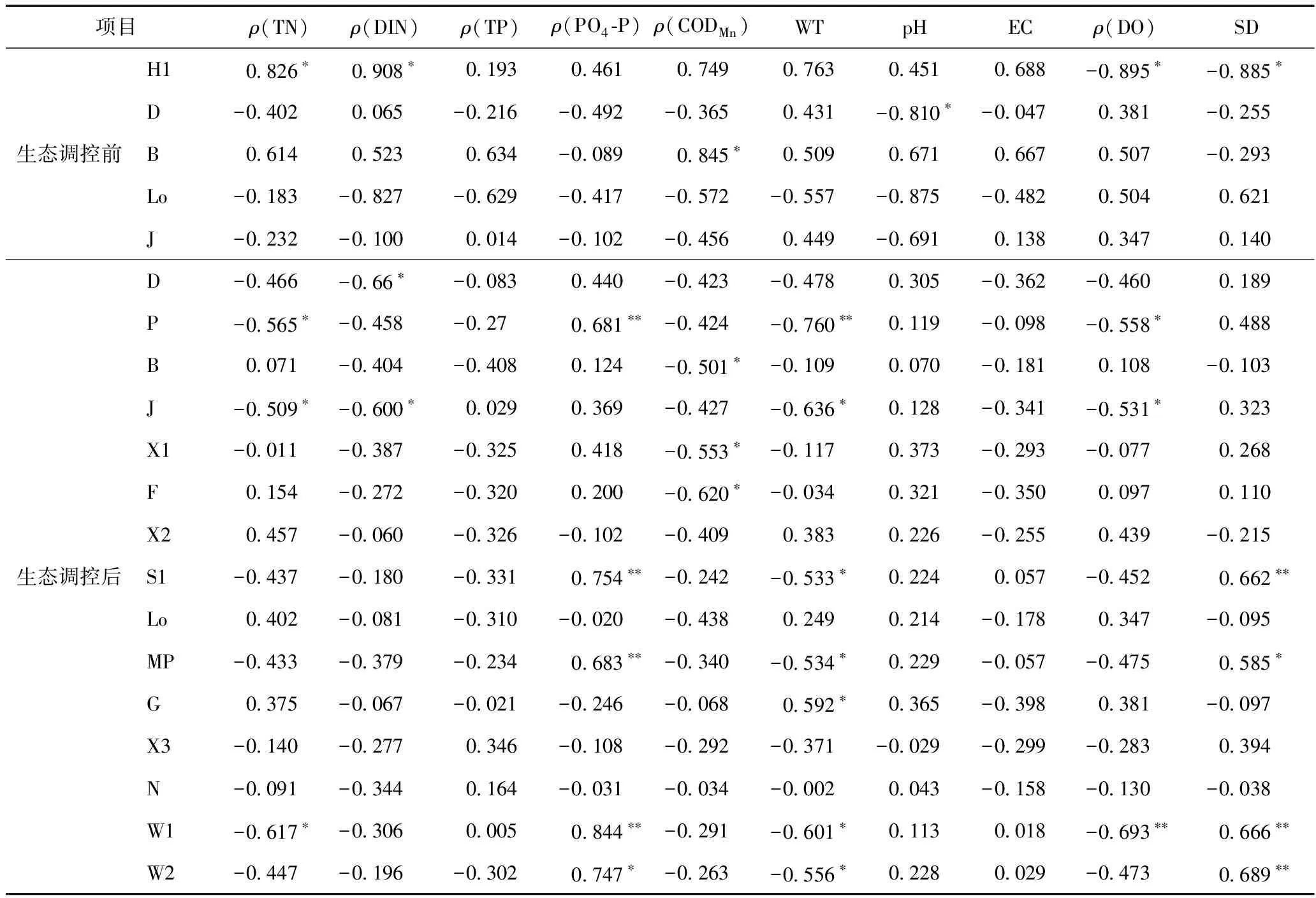
表3 生态调控前后花溪水库浮游植物功能群与环境因子Pearson相关性分析
注:*表示在P<0.05上显著相关; ** 表示在P<0.01上极显著相关.
3 讨论
3.1 生态调控前后花溪水库浮游植物功能群变化特征
浮游植物功能群变化主要受环境因子的影响,当水体生态平衡被打破,浮游植物会通过改变群落结构达到新的平衡状态[11]. 通过物理-生物调控后,花溪水库环境因子(如营养盐浓度)发生了明显改变,同时浮游植物群落结构也发生了变化. 生态调控前功能群H1占主导地位,主要由水华束丝藻组成,在水库营养盐浓度较高的背景下大量繁殖并发生水华. 生态调控后浮游植物功能群发生了变化.
秋季(9—11月)仍以具有广适性的功能群Lo和适宜生存在中营养的功能群B为主要优势类群,另外进入秋季温度开始降低,光照强度逐渐减小,这使得能够耐受低温低光的功能群DP占一定优势.
冬季(12月—翌年3月)与秋季功能群结构变化不大,仍以功能群BLo占优势,但功能群W1W2MP在这一时期开始出现并占一定优势. 功能群W1适宜在有机质丰富的小型浅水水体,可能由于花溪水库周边农业灌溉,使大量有机质流入库区,为功能群W1提供了竞争条件[25]. 冬末初春营养盐水平开始逐渐升高,这为具有强营养吸收能力的功能群MP和适宜在富营养水体生存的功能群W2提供了合适的条件[11,31].
总体而言,生态调控前花溪水库浮游植物功能群H1占优势,生态调控后优势功能群结构发生了变化,其特征为BLo(春季)→DBLoX2NP(夏季)→DBLoN(秋季)→DMPBLoW1W2(冬季).研究[23]表明,通过生态修复等手段后,水体富营养化水平降低,优势藻种发生改变. 生态调控后花溪水库浮游植物主要以硅藻为主,与黄成等[32]对亚热带地区南水水库的研究结果相似.
3.2 影响浮游植物功能群变化的因素
WT、光照强度、营养盐、水动力、浮游动物的牧食等是影响浮游植物生长和消亡的主要因素[11,33]. 该研究显示,生态调控前SD、ρ(DIN)是影响浮游植物功能群分布的主要环境因子,功能群H1大量繁殖并发生水华. Padisk等[12]研究表明,功能群H1适宜生长在富营养化较高的水体中. 生态调控前ρ(TN)和ρ(TP)最高分别达5.87和0.24 mg/L,这为藻类尤其是蓝藻的生长提供了大量的营养元素,使其能够大量的繁殖并形成优势种. 因而在生态调控中控制水体氮磷的含量,对处理蓝藻水华有重要的作用. Pearson分析显示,生态调控前优势功能群H1与ρ(TN)、ρ(DIN)均呈显著正相关,与SD呈显著负相关. 蓝藻本身有固氮的作用,能够在氮含量较低的水体中大量繁殖[11],为其占据优势提供了条件. 功能群H1大量的繁殖需要消耗水体中的ρ(DO),而ρ(DO)限制了功能群H1的继续繁殖. 大面积水华发生导致湖面SD降低,抑制了光照,而功能群H1对低光照有很好的耐受性[34],促进了功能群H1的繁殖.
生态调控后影响功能群分布的主要环境因子是WT、ρ(DIN)、ρ(DO). 营养盐浓度对浮游植物的生长具有绝对限制性,淡水中浮游植物氮磷的绝对限制阈值范围分别为11~14 μg/L〔以ρ(DIN)计〕、3~4 μg/L〔以ρ(SRP)计〕[1]. 生态调控后花溪水库ρ(DIN)和ρ(SRP)范围分别为1.51~3.12 mg/L、4.15~48.16 μg/L,相比生态调控前营养盐浓度虽有所降低,但仍超过限制阈值,产生这种现象的原因可能与水库周边村寨人类活动以及土壤淋溶有关. 因此,WT、ρ(DO)是影响花溪水库浮游植物生长的主要环境因子,营养盐浓度已不再是影响浮游植物生长的限制性因素. 温度能够通过光合作用影响浮游植物酶促反应或呼吸作用从而直接影响浮游植物的生长[35],花溪水库虽处于亚热带季风气候,但不同季节和昼夜的WT依然存在明显的差异. Reynolds等[11]认为,蓝藻、绿藻适宜在较高WT下生长,硅藻适宜在较低WT下生长. 花溪水库年均WT相对较低,适合功能群B/Lo生长. 夏季WT相对较高,RDA分析显示,WT与功能群X2呈正相关,功能群X2在该时期占据优势. 贵州高原水库其他研究[36-37]中也发现,WT是影响浮游植物功能群组成的主要因素. 研究[38]表明,水库中ρ(DO)主要受浮游植物生物量的影响. 秋冬季浮游植物生物量处于较低水平,水库中营养充分但光照不足,限制了水库中浮游植物生长繁殖,从而浮游植物呼吸作用强于光合作用,消耗大量O2,释放出CO2;春夏季随着温度的升高,浮游植物大量繁殖,从而浮游植物光合作用大于呼吸作用,释放大量O2,从而功能群B、Lo与ρ(DO)呈正相关.
4 结论
a) 生态调控前花溪水库浮游植物共鉴定出4门18种,生态调控后共鉴定出6门66种,根据功能群划分原则,生态调控前共归类出10个功能类群,H1为优势功能群;生态调控后共归类出20个功能类群,B、Lo为主要功能群.
b) 受生态调控和环境因子的影响,花溪水库优势功能群结构发生了变化,其变化特征为BLo(春季)→DBLoX2NP(夏季)→DBLoN(秋季)→DMPBLoW1W2(冬季). 优势功能群B生物量在春季达到峰值(3 056.3 μgL),与其对低营养环境有良好的耐受性有关;优势功能群Lo生物量在秋季达到峰值(2 900.9 μgL),与组成功能群Lo的甲藻特性有关.
c) RDA分析表明,生态调控前影响浮游植物生长的影响因子为SD、ρ(DIN);生态调控后影响浮游植物生长的影响因子为WT、ρ(DO)、ρ(DIN).
参考文献(References):
[1] REYNOLDS C S.The ecology of freshwater phytoplankton[M].Cambridge:Cambridge University Press,2006:24-28.
[2] 赵孟绪,雷腊梅,韩博平.亚热带水库浮游植物群落季节变化及其影响因素分析:以汤溪水库为例[J].热带亚热带植物学报,2005,13(5):386-392.
ZHAO Mengxu,LEI Lamei,HAN Boping.Seasonal change in phytoplankton communities in Tangxi Reservoir and effecting factors[J].Journal of Tropical and Subtropical Botany,2005,13(5):386-392.
[3] 许金花,潘伟斌,张海燕.城市小型浅水人工湖泊浮游藻类与水质特征研究[J].生态科学,2007,26(1):36-40.
XU Jinhua,PAN Weibin,ZHANG Haiyan.Studying on planktonic algae and characteristics of water quality of some and adlittoral artificial lake in the city zone[J].Ecological Science,2007,26(1):36-40.
[4] HOYER A B,MORENOOSTOS E,VIDAL J,etal.The influence of external perturbations on the functional composition of phytoplankton in a Mediterranean Reservoir[J].Hydrobiologia,2009,636(1):49-64.
[5] HAMILTON P B,LAVOIE I,LEY L M,etal.Factors contributing to the spatial and temporal variability of phytoplankton communities in the Rideau River (Ontario,Canada)[J].River Systems,2011,19(3):189-205.
[6] XIAO Lijuan,WANG Tian,HU Ren,etal.Succession of phytoplankton groups regulated by monsoonal hydrology in a large canyon-shaped reservoir[J].Water Research,2011,45(16):5009-5019.
[7] LV Hong,YANG Jun,LIU Lemian.Temporal pattern prevails over spatial variability in phytoplankton communities from a subtropical water supply reservoir[J].Oceanological and Hydrobiological Studies,2013,42(4):420-430.
[8] SOMMER U,ADRIAN R,DOMIS L D S,etal.Beyond the plankton ecology group (PEG) model:mechanisms driving plankton succession[J].Annual Review of Ecology Evolution & Systematics,2012,43(1):429-448.
[9] GAEDKE U,RUHENSTROTHBAUERR M,WIEGAND I,etal.Biotic interactions may overrule direct climate effects on spring phytoplankton dynamics[J].Global Change Biology,2010,16(3):1122-1136.
[10] ABRANTES N,ANTUNES S C,PEREIRA M J,etal.Seasonal succession of cladocerans and phytoplankton and their interactions in a shallow eutrophic lake (Lake Vela,Portugal)[J].Acta Oecologica,2006,29(1):54-64.
[11] REYNOLDS C S,HUSZAR V,KRUK C,etal.Towards a functional classification of the freshwater phytoplankton[J].Journal of Plankton Research,2002,24(5):417-428.
[13] MARIA C,LANCONA M,LEITAO M,etal.Phytoplankton functional response to spatial and temporal differences in a cold and oligotrophic lake[J].Hydrobiologia,2016,764(1):199-209.
[14] 黄国佳,李秋华,陈椽,等.贵州高原红枫湖水库浮游植物功能分组及其时空分布特征[J].生态学报,2015,35(17):5573-5584.
HUANG Guojia,LI Qiuhua,CHEN Chuan,etal.Phytoplankton functional groups and their spatial and temporal distribution characteristics in Hongfeng Reservoir,Guizhou Province[J].Acta Ecologica Sinica,2015,35(17):5573-5584.
[15] YU Hongxian,WU Junhua,etal.Seasonal dynamics of phytoplankton functional groups and its relationship with the environment in river:a case study in northeast China[J].Journal of Freshwater Ecology,2012,27(3):429-441.
[16] BECKER V,HUSZAR V L M,CROSSETTI L O.Responses of phytoplankton functional groups to the mixing regime in a deep subtropical reservoir[J].Hydrobiologia,2009,628(1):137-151.
[17] DEVERCELLI M.Phytoplankton of the Middle Paraná River during an anomalous hydrological period:a morphological and functional approach[J].Hydrobiologia,2006,563(1):465-478.
[18] DOKULIL M,TEUBNER K.Steady state phytoplankton assemblages during thermal stratification in deep alpine lakes.do they occur?[J].Hydrobiologia,2003,502(123):65-72.
[19] 董静,李根保,宋立荣.抚仙湖、洱海、滇池浮游藻类功能群1960s以来演变特征[J].湖泊科学,2014,26(5):735-742.
DONG Jing,LI Genbao,SONG Lirong.Historical changes of phytoplankton functional groups in Lake Fuxian,Lake Erhai and Lake Dianchi since 1960s[J].Journal of Lake Sciences,2014,26(5):735-742.
[20] KRUK C,MAZZEO N,LACEROT G,etal.Classification schemes for phytoplankton:a local validation of a functional approach to the analysis of species temporal replacement[J].Journal of Plankton Research,2002,24(9):901-912.
[21] 李秋华.贵州高原水库富营养化特征及评价[J].贵州师范大学学报(自然科学版),2018,36(2):1-8.
LI Qiuhua.Characteristic and evaluation of eutrophication in Guizhou Plateau reservoirs[J].Journal of Guizhou Normal University(Natural Science),2018,36(2):1-8.
[22] 韩博平.中国水库生态学研究的回顾与展望[J].湖泊科学,2010,22(2):151-160.
HAN Boping.Reservoir ecology and limnology in China:a retrospective comment[J].Journal of Lake Sciences,2010,22(2):151-160.
[23] 濮培民,李裕红,张晋芳,等.用生态修复调控浮游植物种群局部控制富营养化:以贵州红枫湖水质生态修复工程为例[J].湖泊科学,2012,24(4):503-512.
PU Peimin,LI Yuhong,ZHANG Jinfang,etal.Eutrophication control in local area through phytoplankton population regulation by eco-remediation:a case study on aqua-eco-remediation engineering in Lake Hongfeng,Guizhou Province[J].Journal of Lake Sciences,2012,24(4):503-512.
[24] 李秋华,夏品华,吴红,等.用物理-生态集成技术局部控制富营养化[J].环境科学,2012,33(7):2303-2308.
LI Qiuhua,XIA Pinhua,WU Hong,etal.Eutrophication control in local area by physic-ecological engineering[J].Environmental Science,2012,33(7):2303-2308.
[25] 胡鸿钧,魏印心.中国淡水藻类:系统、分类及生态[M].北京:科学出版社,2006:23-925.
[26] 孙军,刘东艳,钱树本.浮游植物生物量研究Ⅰ:浮游植物生物量细胞体积转化法[J].海洋学报,1999,21(2):75-85.
SUN Jun,LIU Dongyan,QIAN Shuben.Study on phytoplankton biomass Ⅰ:phytoplankton measurement biomass from cell volume or plasma volume[J].Acta Oceanologica Sinica,1999,21(2):75-85.
[27] 刘鸿亮.湖泊富营养化调查规范[M].北京:中国环境科学出版社,1987:291-294.
[28] 林少君,贺立静,黄沛生,等.浮游植物中叶绿素a提取方法的比较与改进[J].生态科学,2005,24(1):9-11.
LIN ShaoJun,HE Lijing,HUANG Peisheng,etal.Comparison and improvement on the extraction method for chlorophyll a in phytoplankton[J].Ecologic Sciences,2005,24(1):9-11.
[29] 黄享辉,胡韧,雷腊梅,等.南亚热带典型中小型水库浮游植物功能类群季节演替特征[J].生态环境学报,2013(2):311-318.
HUANG Xianghui,HU Ren,LEI Lamei,etal.Seasonal succession of phytoplankton functional groups in typical small and medium-sized reservoirs in southern China[J].Ecology and Environmental Sciences,2013,22(2):311-318.
[30] 张怡,胡韧,肖利娟,等.南亚热带两座不同水文动态的水库浮游植物的功能类群演替比较[J].生态环境学报,2012,21(1):107-117.
ZHANG Yi,HU Ren,XIAO Lijuan,etal.Comparative analysis of the phytoplankton functional groups in two reservoirs with different hydrodynamics in southern China[J].Ecology and Environmental Sciences,2012,21(1):107-117.
[31] WANG L,CAI Q H,TAN L,etal.Phytoplankton development and ecological status during a cyanobacterial bloom in a tributary bay of the Three Gorges Reservoir,China[J].Science of the Total Environment,2011,409(19):3820-3828.
[32] 黄成,岳强,史元康,等.广东省韶关市3座大中型饮用水水库浮游植物功能类群特征[J].生态科学,2014,33(2):282-287.
HUANG Cheng,YUE Qiang,SHI Yuankang,etal.Characteristics of phytoplankton functional groups in main reservoirs for large and medium sized drinking water supply in Shaoguan,China[J].Ecological Science,2014,33(2):282-287.
[33] 黄国佳,李秋华,陈椽,等.贵州高原三板溪水库浮游植物功能群时空分布特征[J].环境科学学报,2015,35(2):418-428.
HUANG Guojia,LI Qiuhua,CHEN Chuan,etal.Phytoplankton functional groups and their spatial and temporal distribution characteristics in Sanbanxi Reservoir,Guizhou Province[J].Acta Scientiae Circumstantiae,2015,35(2):418-428.
[34] 董静,李艳晖,李根保,等.东江水系浮游植物功能群季节动态特征及影响因子[J].水生生物学报,2013,37(5):836-843.
DONG Jing,LI Yanhui,LI Genbao,etal.Seasonal dynamics characteristics and affecting physical factors of phytoplankton functional groups in Dongjiang River[J].Acta Hydrobiolagica Sinica,2013,37(5):837-843.
[35] BLINN D W.Diatom community structure along physicochemical gradients in saline lakes[J].Ecology,1993,74(4):1246-1263.
[36] 陈倩,李秋华,胡月敏,等.贵州百花水库浮游藻类功能群时空分布特征及影响因子分析[J].环境科学研究,2018,31(7):1266-1274.
CHEN Qian,LI Qiuhua,HU Yuemin,etal.Spatial and temporal distribution characteristics of phytoplankton functional groups and their influencing factors in Baihua Reservoir,Guizhou Province[J].Research of Environmental Sciences,2018,31(7):1266-1274.
[37] 胡月敏,李秋华,朱冲冲,等.基于功能群对比分析黔中普定水库和桂家湖水库浮游植物群落结构特征[J].湖泊科学,2018,30(2):403-416.
HU Yuemin,LI Qiuhua,ZHU Chongchong,etal.Contrasting phytoplankton structure based on functional groups of Puding Reservoir and Guijiahu Reservoir in the Central of Guizhou Province[J].Journal of Lake Sciences,2018,30(2):403-416.
[38] 俞焰,刘德富,杨正健,等.千岛湖溶解氧与浮游植物垂向分层特征及其影响因素[J].环境科学,2017,38(4):1393-1402.
YU Yan,LIU Defu,YANG Zhengjian,etal.Vertical stratification characteristics of dissolved oxygen and phytoplankton in Thousand-Island Lake and their influencing factors[J].Environmental Science,2017,38(4):1393-1402.
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
当代水产(2022年6期)2022-06-29
乡村地理(2021年4期)2021-03-28
今日农业(2020年19期)2020-12-14
文苑(2020年5期)2020-06-16
安徽农学通报(2020年7期)2020-05-26
流行色(2019年10期)2019-12-06
装饰装修天地(2019年9期)2019-10-21
中国野生植物资源(2019年2期)2019-06-11
当代贵州(2015年25期)2015-06-13
