
NaCl胁迫对葡萄砧木光合特性与叶绿素荧光参数的影响
2020-03-15 03:26:08尹勇刚袁军伟刘长江韩斌李敏敏孙艳贾楠郭紫娟赵胜建
中国农业科技导报 2020年8期
尹勇刚, 袁军伟, 刘长江, 韩斌, 李敏敏, 孙艳,贾楠, 郭紫娟, 赵胜建
(河北省农林科学院昌黎果树研究所, 河北 秦皇岛 066000)
葡萄是世界上栽培面积最广的果树之一。据联合国粮农组织(FAO)统计,2017年世界葡萄种植面积达693.14万hm2,年产量7 427.66万t;中国葡萄栽培总面积达77.86万hm2,产量1 316.08万t[1]。我国西北地区与渤海湾区因其极为适宜的气候条件,成为我国优质葡萄的重要产区,而该地区日趋加剧的盐渍化也严重制约了葡萄产业的进一步发展[2-3]。研究发现,具有耐盐性的砧木可提高葡萄嫁接组合的整体耐盐能力[4-5],不同葡萄砧木品种的耐盐差异因其对盐胁迫的反应机制不同所致[6]。研究葡萄砧木在盐胁迫下生理变化规律是探索葡萄抵抗盐害的重要方式,对避免葡萄生产遭遇潜在的盐渍伤害具有重要意义。
盐碱等生态胁迫极易影响植物光合作用[7]。盐胁迫通常导致植物叶片光合效率下降[8],研究表明,轻度盐胁迫主要因渗透胁迫引起气孔限制所致[9-10],而中高度盐胁迫下光合受阻主要为非气孔限制[11-12]。然而,这种最终导致植物生长停滞乃至死亡的非气孔限制因素及其在不同盐浓度胁迫下的主次作用尚不明确。
二氧化碳作为原料与吸收转化光能的叶绿素是对光合反应产生直接影响的两个重要因素。位于类囊体中的光反应中心是光合原初反应的场所,是光合反应顺利进行的保障。叶绿素荧光参数可实时反映光合结构状态,现已经广泛应用于监测非生物胁迫下果树[13]、蔬菜[14]及作物[15]等多种植物的受害程度。本研究围绕光合反应3个重要指标——叶绿素含量、光合气体交换和叶绿素荧光参数,对盐胁迫下的两种典型葡萄砧木品种进行对比分析,研究不同盐浓度下葡萄抵抗胁迫的方式和主次转化,为耐盐葡萄砧木在盐害多发区的推广应用提供理论依据。
1 材料与方法
1.1 试验材料
基于本课题组前期对21份葡萄砧木耐盐性鉴定结果[16],本研究以具有典型特征的耐盐性品种101-14M(V.riparia×V.rupestris)和盐敏感品种188-08(V.berlandieri×V.riparia)作为试验材料。
1.2 试验设计
试验于2018年4—7月在河北省农林科学院昌黎果树研究所创新基地进行。4月初将1年生砧木苗种植于装有培养基质(蛭石:珍珠岩=3∶1)的大营养钵中,营养钵上口径13.5 cm,下口径9.5 cm,高22.0 cm,底部留孔,于避雨棚内进行常规管理。当砧木苗长到8~10片叶时,选取生长势均匀一致的苗木,用于NaCl胁迫处理。
每天分别用浓度为4、6、8和10 g·kg-1的NaCl溶液对2种砧木品种进行定量(每株0.8 L)灌溉,以同样量的清水灌溉为对照(CK)。每个品种各设5个处理,每处理10株,3次重复。盐胁迫处理28 d后,测定光合气体交换参数和叶绿素荧光参数,并从每株相同位置采6~8片叶用于测定叶绿素质量分数。
1.3 测定指标与方法
1.3.1叶绿素含量测定 参照Arnon[17]的方法,将采集回来的新鲜叶片剪碎混匀,称取0.2 g,于φ=80%丙酮溶液中暗处浸提12 h。用UV-2550紫外可见分光光度计(日本岛津公司)分别于663和646 nm波长下比色测定吸光值。
1.3.2光合气体交换参数测定 选取苗木上部第6~8片功能叶,采用LI-6400XT便携式光合仪(美国LI-COR公司)于晴天9:00—11:00测定光合气体交换参数:叶片净光合速率(net photosynthetic rate,Pn)、气孔导度(stomatal conductance,Gs)、胞间CO2浓度(intercellular CO2concentration,Ci)和蒸腾速率(transpiration rate,Tr)。测定控制光强为1 600 μmol·m-2·s-1,CO2体积分数为360 μL·L-1,相对湿度为70%。
1.3.3叶绿素荧光参数测定 利用PAM-2500便携式调制叶绿素荧光仪(德国Walz公司)测定叶绿素荧光参数。叶片暗适应30 min后执行饱和脉冲,测定初始荧光产量(minimal fluorescence,F0)及最大荧光产量(maximal fluorescence,Fm);自然光照40 min后执行饱和脉冲,测量光下初始荧光(F0′)、光下最大荧光(Fm′)和稳态荧光(fluorescence in stable state,Fa),并计算以下参数。
可变荧光(variable fluorescence,Fv)=Fm-F0
光系统Ⅱ(photosystem Ⅱ complex,PSⅡ)最大光化学效率=Fv/Fm
PSⅡ实际光化学效率Yield=(Fm′-Fa)/Fm′
光化学淬灭系数(photochemical quenching,qP)=(Fm′-Fa)/(Fm′-F0)
非光化学淬灭系数(non-photochemical quenching,NPQ)=Fm/Fm′-1
1.4 数据处理
采用Microsoft Excel 2013进行数据统计与图表制作;使用SPSS 20.0软件以Duncan法对数据进行单因素方差分析。
2 结果与分析
2.1 NaCl胁迫下2种葡萄砧木叶片叶绿素含量变化
图1显示,盐害发生后,2种葡萄砧木的叶绿素a与叶绿素b含量均呈现先升后降的走势。NaCl浓度为6 g·kg-1时,188-08的叶绿素a含量开始下降,低于CK 14.70%;在8 g·kg-1的盐浓度下,101-14M的叶绿素a含量开始显著下降,相比CK下降8.50%。随NaCl浓度升高,188-08叶片叶绿素a含量降低幅度明显大于101-14M。101-14M叶片叶绿素b含量显著高于188-08,但两者间差异随NaCl浓度升高逐渐缩小。可见,在较高浓度NaCl胁迫下,维持较高的叶绿素含量,尤其是叶绿素a的含量,是保证葡萄砧木耐盐能力的关键因素。
2.2 NaCl胁迫下2种葡萄砧木叶片光合气体交换参数变化
净光合速率(Pn)是光合系统功能的直接体现,也是检验植株光合系统工作正常与否的指标。从图2可以看出,随NaCl浓度升高,2种葡萄砧木的Pn显著降低。在4、6、8、10 g·kg-1的盐胁迫下,101-14M的Pn相比于CK分别降低了6.99%、27.12%、49.66%、60.88%;188-08的Pn相比于CK则分别降低了43.01%、52.29%、64.93%、68.66%,下降幅度显著高于101-14M。188-08在NaCl浓度为4 g·kg-1时Pn出现显著下降,在8 g·kg-1时下降幅度相对放缓;101-14M叶片Pn亦随NaCl浓度升高而逐步下降,但均显著高于188-08。
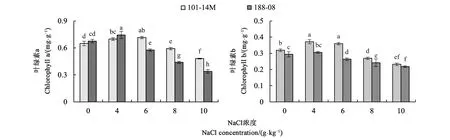
注:不同小写字母表示差异在P<0.05水平具有统计学意义。Note: Different small letters indicate significant difference at P<0.05 level.图1 NaCl胁迫下两葡萄砧木叶片叶绿素含量变化Fig.1 Effect of salt stress on chlorophyll content in leaves of two rootstocks
在4、6、8、10 g·kg-1的NaCl胁迫下,101-14M气孔导度(Gs)明显下降,与CK相比,依次降低了31.66%、53.83%、74.67%、86.81%;188-08的Gs则分别降低了63.29%、70.63%、81.12%、82.17%,在4 g·kg-1的NaCl处理下降幅最大,随后放缓,在8与10 g·kg-1NaCl处理间无显著差异。CK处理下,101-14M的Gs明显高于188-08,直至NaCl浓度达到10 g·kg-1前,101-14的Gs均保持高于188-08。
胞间二氧化碳浓度(Ci)是分析光合速率降低原因的指标之一。图2显示,NaCl胁迫处理中,188-08和101-14M的Ci均显著下降,前者相比CK在各盐浓度处理下分别降低了26.88%、32.30%、40.73%、56.75%,后者则分别降低了37.80%、55.95%、58.04%、56.18%。随NaCl浓度的升高,101-14M的Ci逐渐下降,而188-08的Ci则在6 g·kg-1NaCl处理下趋于稳定。不同盐浓度胁迫下。101-14M叶片Ci始终高于188-08。
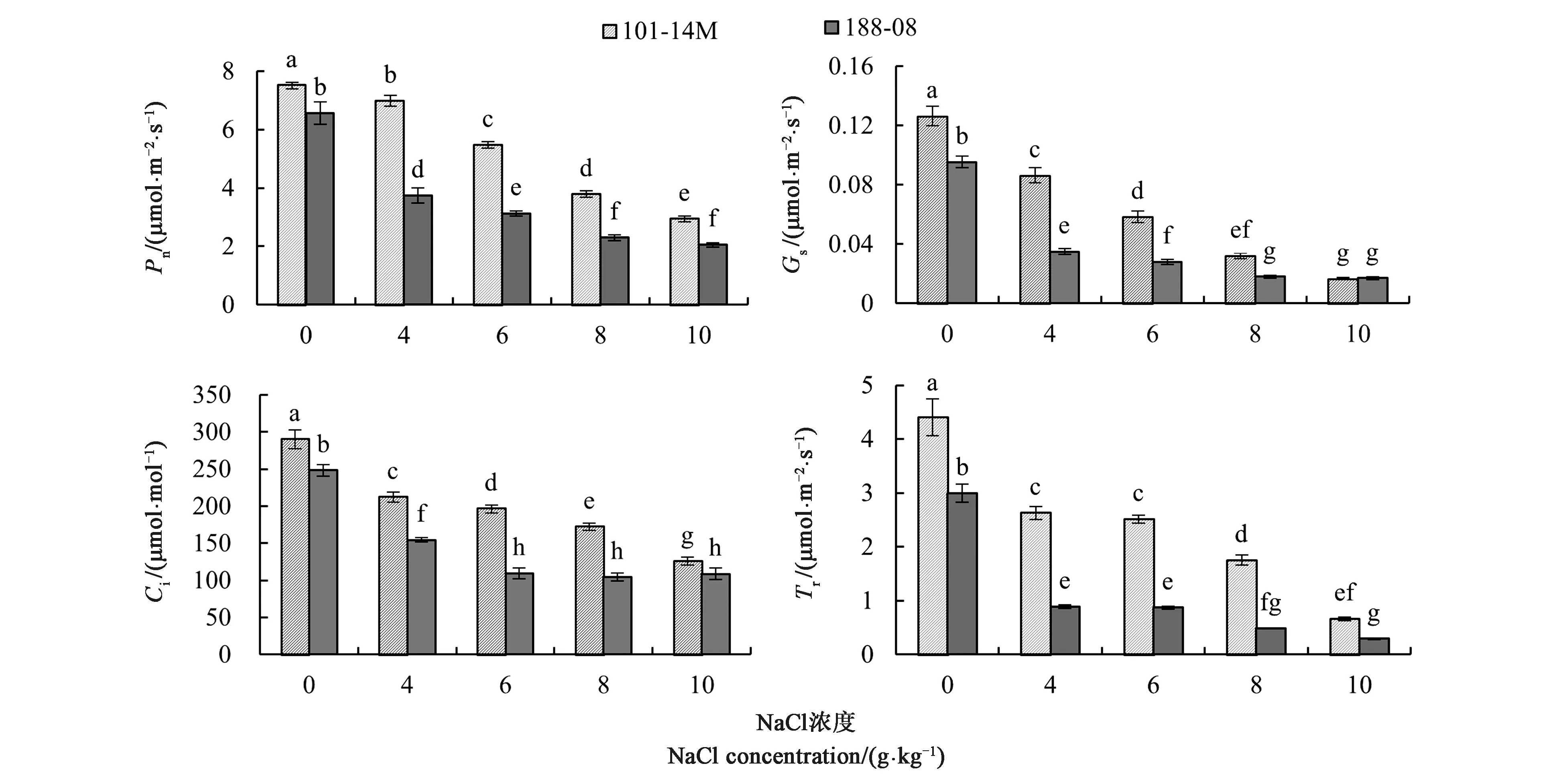
注:不同小写字母表示差异在P<0.05水平具有统计学意义。Note: Different small letters indicate significant difference at P<0.05 level.图2 NaCl胁迫下2种葡萄砧木叶片光合气体交换参数变化Fig.2 Photosynthetic gas exchange parameters changes in leaves of two grape rootstocks under NaCl stress
NaCl胁迫处理后,2种葡萄砧木叶片的蒸腾速率(Tr)显著下降。101-14M和188-08在不同浓度NaCl处理下,Tr均显著低于CK。在4~6 g·kg-1NaCl处理中,2种砧木的Tr较为稳定;NaCl浓度达到8 g·kg-1时,2种砧木品种Tr继续下降。不同浓度NaCl胁迫下,101-14M始终保持着高于188-08的蒸腾速率。当NaCl浓度达到10 g·kg-1时,101-14M和188-08的Tr仅为各自CK的15.08%和9.80%。蒸腾作用最主要的方式是气孔蒸腾,Tr降低主要与Gs有关。
2.3 NaCl胁迫下2种葡萄砧木叶片荧光参数变化
F0为充分暗适应的光合机构的初始荧光强度,是PSⅡ处于完全开放时的荧光产量,代表不参与PSⅡ光化学反应的光能辐射部分。如图3所示,盐胁迫发生后,2种砧木叶片F0先升高后降低。6 g·kg-1NaCl胁迫下,188-08开始降低;NaCl浓度为8 g·kg-1时,101-14M的F0显著降低。F0与叶绿素含量显著相关,也与光合机构关闭程度有关,光合受阻与叶绿素含量增加均会导致F0升高。
暗适应下的最大荧光产量Fm是PSⅡ完全关闭状态下的荧光产量,反映通过PSⅡ的电子传递情况。图3显示,随NaCl浓度升高,Fm呈下降趋势。NaCl浓度达到4 g·kg-1时,2种砧木品种与各自对照无显著差异,随后下降。结果表明,NaCl在4 g·kg-1时,PSⅡ电子传递受外界影响较小;达到6 g·kg-1时,PSⅡ电子传递开始受阻,并随盐浓度升高,电子传递受阻程度逐渐增大。相同浓度NaCl处理下,101-14M显示出高于188-08的电子传递效率。
从图3可以看出,2种葡萄砧木品种CK中的PSⅡ最大光化学效率Fv/Fm、实际光化学效率Yield值无显著差异。NaCl处理中,2种葡萄砧木Fv/Fm、Yield和qP均随着NaCl处理浓度升高而下降,188-08降幅明显,同浓度NaCl处理下低于或显著低于101-14M。2种葡萄砧木CK中Fv/Fm(0.76±0.004 5,0.76±0.004 0)处于活体叶片正常范围内,188-08与101-14M叶片Fv/Fm分别在4和6 g·kg-1的 NaCl处理下,出现显著下降。188-08Yield值下降速度大于101-14M,表明其PSⅡ反应中心更易受到损伤,关闭程度更大。qP是由光合作用引起的荧光淬灭,可反映植物光合活性的高低。2种葡萄砧木叶片qP随着盐浓度升高而逐渐稳定下降,101-14M的qP显著高于188-08,NaCl浓度达到10 g·kg-1时,101-14M与188-08的qP分别为各自CK的68.8%和48.2%。
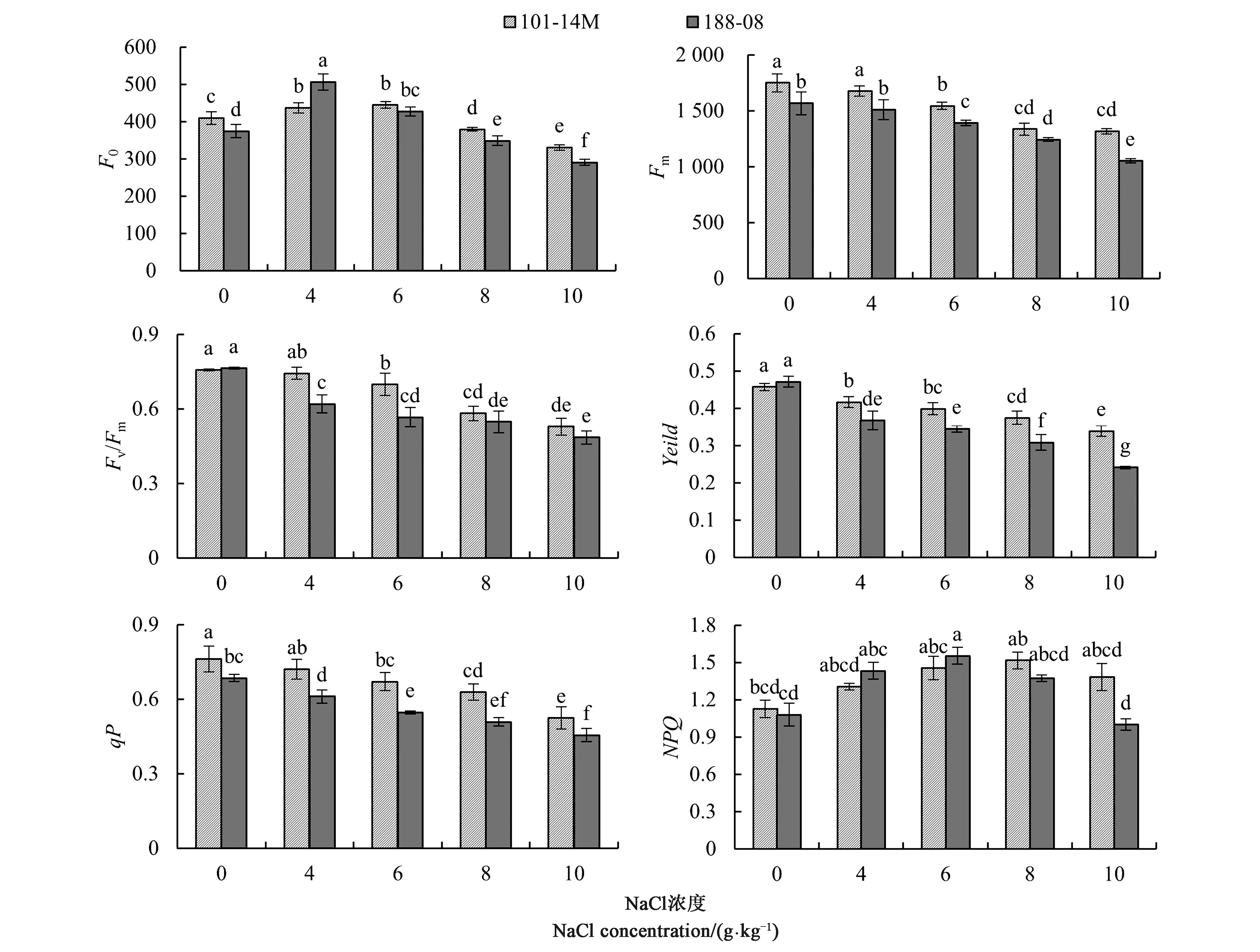
注:不同小写字母表示差异在P<0.05水平具有统计学意义。Note: Different small letters indicate significant difference at P<0.05 level.图3 NaCl胁迫下2种葡萄砧木叶片叶绿素荧光参数变化Fig.3 Chlorophyll fluorescence parameters changes in leaves of two grape rootstocks under NaCl stress
非光化学淬灭系数NPQ是由热耗散引起的荧光淬灭,反映了植物耗散过剩光能为热的能力,即光保护能力。图3显示,2种葡萄砧木的NPQ随NaCl浓度升高而呈先升后降趋势。101-14M 的NPQ先小幅上升,表明光合作用在减弱,使得一部分能量以热量形式耗散,在10 g·kg-1NaCl处理下小幅下降,略高于对照;188-08变化幅度大于101-14M,表明其光合受抑制程度更高,其NPQ在NaCl浓度为8 g·kg-1时开始降低,在10 g·kg-1NaCl处理下显著低于CK。NPQ增加通常伴随F0升高,PSⅡ失活是两者升高的原因之一。
3 讨论
2种葡萄砧木在NaCl胁迫发生时,各生理参数变化趋势相似,但对盐浓度改变时的反应存在差异。植株根部细胞遭受NaCl胁迫使水分向地上部分运输受到阻碍,保卫细胞失水导致气孔逐渐闭合,气孔导度Gs下降,这与盐胁迫下其他植物变化趋势相同[18]。随Gs降低,188-08的Ci在6~10 g·kg-1的NaCl下保持平稳,表明光合过程对CO2的利用率下降,导致光合受阻的已不是气孔限制[19];101-14M的Ci则持续下降,表明CO2同化用量多于吸入CO2量,气孔限制仍是导致其光合下降的原因之一。
伴随细胞失水,盐离子进入细胞,抑制叶绿素前体的合成,中等浓度NaCl促进叶绿素降解[20]。叶绿素通过接收传递和转化光能为光合作用提供能量,叶绿素的减少会直接降低光合产量。本研究中叶绿素含量在较低NaCl浓度下升高,与李学孚等[21]对‘鄞红’葡萄研究结论不同,或与水培法栽植及更长的盐胁迫时长有关。叶绿素含量在低浓度NaCl胁迫下之所以升高或许有以下2个原因:根细胞通过调节膜透性限制Na+[22]或Cl-[23]进入细胞,使叶绿素合成及叶绿素酶降解未受到NaCl明显的影响;气孔限制引起光合受阻使得对叶绿素利用率降低,导致叶绿素累积。在对大豆[24]、棉花[25]和向日葵[26]的相关研究中也得出相似结论。随NaCl浓度升高,2种砧木叶绿素含量先后下降,表明叶绿素合成遭到盐离子较大程度的破坏,叶绿素含量减少进一步限制了188-08的光合作用。这或许亦是8 g·kg-1NaCl胁迫下101-14M叶片光合下降的主要原因之一,在对盐胁迫下黄瑞香[27]、萝卜[28]和荆芥[29]等植物的研究中亦有相同结论。
与此同时,NaCl胁迫导致Fm、Fv/Fm、Yield和qP降低,表明PSⅡ活性在盐害发生后即遭受抑制,这与Athar等[30]对油菜的研究结果一致;同时可以看到,在中低浓度NaCl处理中,部分参数(如4 g·kg-1NaCl下的Fm)相比对照变化不显著,说明植物体对盐胁迫产生了一定的抵抗,这种抵抗能力在101-14M中表现更为明显。Yield下降幅度高于Fv/Fm,也表明盐害发生时就引发了光抑制[31]。F0在盐胁迫发生后升高,升高幅度也反映了光反应活性受抑制程度,随NaCl浓度升高,188-08和101-14M的F0先后降低,或因接收转化光能的叶绿素合成受阻导致,F0与叶绿素含量显著相关。盐胁迫使葡萄砧木叶片荧光淬灭系数qP下降,是光化学反应效率降低的明显特征,这与张玲等[32]对辣椒的研究结果类似。同时过剩的能量以热耗散形式辐射,导致NPQ升高,NPQ随后在高盐浓度下降低,或因叶绿素含量下降致使光能转化率降低所致,同时PSⅡ受损伤亦使光能利用率下降[33]。
总之,盐胁迫下101-14M各生理指标相比188-08变化幅度更小,在高浓度NaCl下仍有相对较高的光合产出。本研究对前人所述的非气孔限制因素开展了进一步研究,认为非气孔限制主要为叶绿素和PSⅡ限制,不同NaCl浓度胁迫引起光合受阻的主要限制因素不同。盐浓度为6 g·kg-1时,188-08光合速率降低主要是由气孔限制转向叶绿素限制,在8 g·kg-1NaCl胁迫下,PSⅡ受到严重损伤;而101-14M叶片在8 g·kg-1盐胁迫下,主要受到叶绿素限制;在10 g·kg-1盐胁迫下,其PSⅡ反应中心遭遇损伤。表明101-14M可忍耐8~10 g·kg-1的NaCl胁迫。同时,通过长时盐胁迫,对葡萄砧木的耐盐能力相对量化,为更好地选择利用耐盐砧木品种提供数据支持。
猜你喜欢
草业学报(2022年5期)2022-07-08 06:24:40
干旱地区农业研究(2022年1期)2022-01-28 04:51:30
草业学报(2021年3期)2021-03-22 02:22:00
落叶果树(2021年6期)2021-02-12 01:29:12
装备制造技术(2019年12期)2019-12-25 03:06:52
科技创新与品牌(2019年12期)2019-02-06 04:03:29
草业学报(2018年10期)2018-10-19 05:25:44
现代园艺(2017年21期)2018-01-03 06:41:45
电镀与环保(2017年5期)2017-12-19 12:06:03
湖南农业(2017年1期)2017-03-20 14:04:46
